油脂體穩定性影響因素及機理的研究進展
2021-09-02 06:47:22官夢姝宋超男姜瞻梅江連洲侯俊財
食品工業科技 2021年16期
關鍵詞:油脂
官夢姝,徐 聰,姜 瑞,宋超男,馮 雪,劉 月,李 佳,姜瞻梅,江連洲,侯俊財
(東北農業大學食品學院,黑龍江哈爾濱 150030)
脂肪、碳水化合物和蛋白質是三種主要的能量儲備形式,脂肪是迄今為止人們所知最有效的能量儲存形式,其所含的能量是淀粉或蛋白質的兩倍多[1]。種子中貯藏的脂肪是在種子發育過程中合成的,可以在胚組織中(大豆、油菜籽和葵花籽)合成,也可以在胚乳(蓖麻子等)中合成。脂質生物合成酶在種子胚胎細胞中合成并在干燥的種子細胞中積累[2]。在大多數植物中,貯藏的脂肪為三酰基甘油(triacylglycerol,TAG)的形式,它由3個酯化為甘油骨架的脂肪酰基鏈組成[3]。在種子中,TAG被分割成小的亞細胞液滴,為生命活動過程中脂肪酶的作用提供更大的表面積。種子中的脂肪最終會沉積到油脂體中,油脂體還存在于包括種子外的其它組織或器官中。用電子顯微鏡觀察植物細胞,特別是用戊二醛和四氧化鋨固定后的種子貯藏組織,發現粒子有不透明的電子基質,周圍有電子致密的邊界。這些粒子的特征被解釋為一個脂質基質被一層特殊的膜所包圍,這種特殊的脂肪顆粒被稱為油脂體[4]。很多研究認為,油脂體可能產生于細胞中內質網的特定亞結構區域或原核細胞的質膜中,且其形成及大小受特定蛋白質的調控。具有油脂體的組織能夠抵抗干旱等不良環境,具有長期儲存油脂的能力[5]。
作為天然的預乳化的油,油脂體應用于食品體系中不需要乳化劑和均質過程,并且從植物中獲得的油脂體(如花生、葵花籽和大豆)由于其更健康、經濟等優點逐漸開始以天然乳液形式應用于食品等領域中[6]。本文論述了油脂體結構組成、乳液穩定性影響因素及影響機理、油脂體的應用,為油脂體的深入研究提供參考。
1 油脂體
1.1 油脂體結構與組成
植物種子儲存TAG作為種子萌發和萌發后生長的能量儲備,TAG存在于被稱為油脂體的小的離散的細胞內細胞器中。單個油脂體單位呈球形,其直徑從0.5~2.5μm不等,其大小因油脂體的來源、營養狀況和環境因素的影響有所差異[1]。油脂體主要由TAG,少量的磷脂和蛋白質構成,油脂體內部為液態TAG,外圍是單層磷脂和油體蛋白(Oleosin)組成的“半單位”膜(如圖1),該膜由13個磷脂分子和1個Oleosin蛋白分子組成[7]。由種子制備的油脂體包含約94%~98%(w/w)的中性脂質,0.6%~2%的磷脂和0.6%~3%的蛋白質,中性脂質主要以TAG的形式存在[8]。油體蛋白中間的疏水區域形成柄狀結構伸入到油脂體內部的TAG中,而其余油體蛋白的頭尾部分則覆蓋了大部分表面區域,由于其兩親性結構,油體蛋白被認為覆蓋了整個油脂體膜的表面,使其保持為小的個體單位,這將有利于提高表面積/體積和促進脂肪酶在發芽期間的作用,這個大小可能代表發芽過程中脂質的有效儲存和動員之間的平衡[9]。除了油體蛋白外,油脂體表面還鑲嵌有少量其他蛋白質。油脂體蛋白表現出特定的時間依賴性積累模式,這表明油脂體蛋白分布可能在種子成熟過程中發生變化[10]。
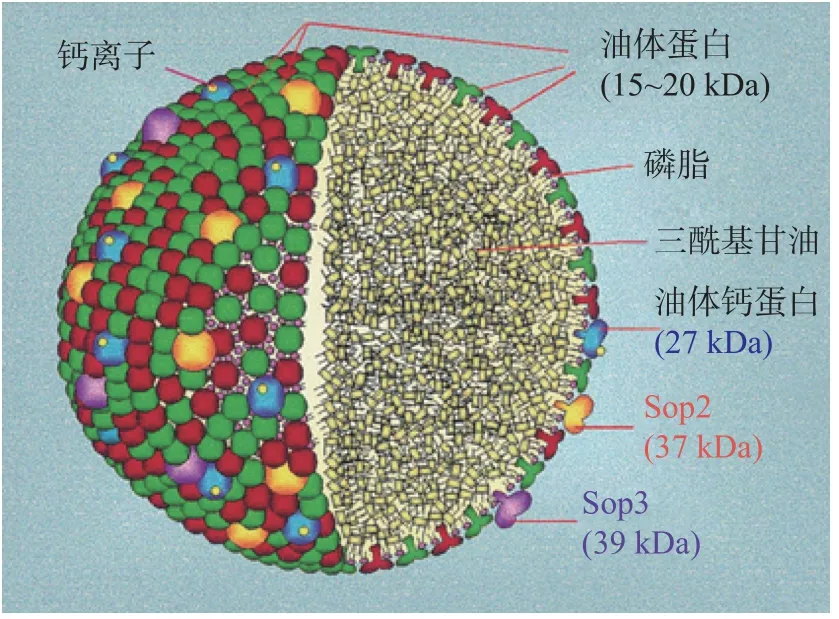
圖1 油脂體結構[3]Fig.1 Structure model of Oil bodies[3]
1.2 油脂體相關蛋白
Tzen等[11]研究發現,通過胰蛋白酶處理油脂體懸浮液,油脂體迅速漂浮到溶液頂部并且油脂體下方懸浮液濁度快速下降,處理僅1 min后觀察到聚結。而與胰蛋白酶處理相反,磷脂酶A 2處理不會導致油脂體懸浮液的濁度以及油脂體粒徑明顯變化,這表明油脂體膜表面的蛋白質對于油脂體的穩定性起著至關重要的作用[7]。對鑲嵌在磷脂膜表面的蛋白進行研究發現,油脂體蛋白質包括油體蛋白(Oleosin)、油體鈣蛋白(Caleosin)和甾醇蛋白(Steroleosin)[12]。
油脂體中蛋白質占比最高的是油體蛋白,油體蛋白是15~30 k Da的堿性蛋白,其被認為是油脂體特有的[1],油體蛋白結構如圖2。裸子植物中僅發現一種油體蛋白即L-油體蛋白,被子植物中存在兩種免疫學上不同的同工型油體蛋白:H-油體蛋白和L-油體蛋白,并且通過免疫熒光顯示它們共存于同一油脂體上,不同種類植物存在相對應的同工型具有較高同源性[13]。油體蛋白包含3個結構區域,一個50~70個殘基的兩親性N末端,一個大約70個殘基的中央疏水性結構域和一個可變長度的兩親性C末端,模型預測,由兩個富含脯氨酸的延伸鏈或“脯氨酸結”連接的反平行鏈形成的中心區域插入油脂體TAG基質中,形成發夾結構,N和C末端結構域駐留在油脂體表面上[11]。油脂體膜上的油體蛋白拓撲結構使帶負電的殘基暴露于細胞質,使油脂體帶電,表面油體蛋白產生的位阻和電斥力可以阻止油脂體的聚集。油體蛋白中含有正確定位于油脂體的信息,可保證大部分油體蛋白分子的穩定[14]。
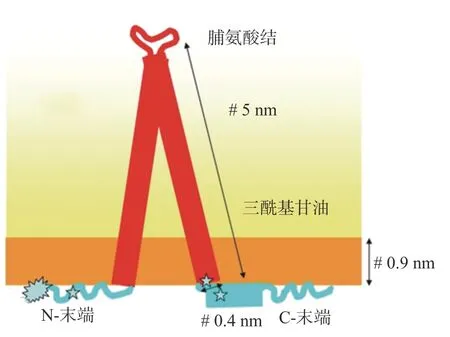
圖2 油體蛋白結構[15]Fig.2 Structure model of oleosin[15]
對芝麻油脂體分離出的蛋白進行研究發現,部分蛋白與具有鈣結合結構域的大米蛋白同源,這部分蛋白被定義為油體鈣蛋白。它們在高等植物中普遍存在,在藻類和真菌中也發現了類似的蛋白質。高等植物的油體鈣蛋白包含3個可辨別的區域,N-末端區域具有相當的親水性,該親水區域朝向細胞質,并包含單個結合Ca2+的EF-hand[16]。EF-hand還結合了Ca2+以外的二價陽離子,并且Ca2+/Mg2+的交換容易影響EF-hand蛋白的構象。由于細胞質中的Mg2+濃度遠高于Ca2+,并且由于體外Ca2+結合測定法人為地使用了較高的Ca2+水平,因此Mg2+可能是單個油體鈣蛋白EF-hand的主要細胞配體[17]。第二個油體鈣蛋白區緊鄰EF-hand,是一個中央疏水域,隨后是脯氨酸豐富的片段。C-末端親水區包含4個潛在的磷酸化位點,其中酪蛋白激酶II 3個,酪氨酸蛋白激酶1個。油體鈣蛋白具有調節油脂體表面積/體積比,加速油脂體中脂類的降解,參與膜和油脂體融合及穩定油脂體結構等作用[18],其功能可能受到鈣結合和磷酸化狀態的調節[3]。
對芝麻油脂體蛋白進行研究發現,部分蛋白能夠與甾醇結合,這類蛋白稱為甾醇蛋白,其分子量大約在40 kD左右。甾醇蛋白的N-末端由α-螺旋構成,具有固醇脫氫酶結合位點;C-末端由α-螺旋組成的7個平行β-鏈形成,其中存在固醇、NADPH結合位點和固醇脫氫酶活性位點[11]。甾醇蛋白是油脂體蛋白質組研究中第三豐富的油脂體相關蛋白,但除了作為油脂體結構蛋白外,其功能尚不清楚。
2 油脂體乳液穩定性研究
油脂體中天然的蛋白-磷脂界面層,使得從油料作物種子中分離出來的油脂體能夠分散在水相中形成水包油(O/W)乳化體系[19]。天然油脂體是軟液滴,但不破裂,并且在高達0.3的壓縮應變下可以完全恢復。由于磷脂-蛋白質膜的存在而產生的和低界面張力有助于油脂體乳液的穩定性[20]。陰離子磷脂的極性頭基與蛋白質之間發生靜電相互作用使得磷脂與油體蛋白相互作用足夠強,可以防止它們在加熱時變性;油體蛋白和兩性離子磷脂之間具有疏水相互作用,可以使油體蛋白具有更穩定的構型[21]。油脂體表面的內源性蛋白提供一定的電荷和空間位阻,使油脂體液滴之間產生較弱的排斥力,也有利于維持油脂體乳液的穩定性[22]。具有較高蛋白質/油比的油脂體,具有較小的尺寸和較強的機械性能,也更穩定。水包油乳液存儲過程中會出現絮凝、聚結和物理失穩,導致乳液液滴粒徑增加,并形成大顆粒的聚集,最終導致相分離[23],油脂的氧化衰敗也會隨之產生。乳液失穩機制如圖3所示。

圖3 乳液失穩機制[24]Fig.3 Mechanism of emulsion instability[24]
在重組油脂體時,研究發現只有當油脂體的TAG、磷脂和蛋白質的比例與天然油脂體中的比例相似時,才能實現油脂體的高穩定性。這表明完整的油脂體結構與組成是維持油脂體穩定性的決定性因素[7]。Niu等[25]在油脂體乳液中加入木瓜蛋白酶后,油脂體表面的蛋白質被酶分解成氨基酸或小分子肽,溶于水相,油滴失去了蛋白質膜,逐漸聚集失穩。油脂體乳液的微觀結構如圖4所示。

圖4 油脂體乳液微觀結構[25]Fig.4 Microscopic structureof oil body emulsion[25]
與人造O/W乳液相比,油脂體乳液在特殊環境下具有更好的理化穩定性。Fisk等[26]比較了合成表面活性劑(DTAB、Tween 20、SDS)穩定的葵花籽油水乳液和天然穩定的葵花籽油脂體懸浮液在不同溫度下的穩定性。結果發現,天然油脂體懸浮液的氫過氧化物的形成顯著低于用合成表面活性劑配制的乳液,并且經鹽洗后純度更高的油脂體乳液顯示出氫過氧化物形成的進一步減少,表明以油脂體形式儲存的油脂具有更高的穩定性,油脂體作為天然食品抗氧化劑具有極大發展潛力。在天然油脂體食品的實際應用和開發中,油脂體乳液的穩定性與油脂體的組成、提取方法、外源蛋白質、溫度、pH和離子強度等條件有關。
2.1 提取方法對油脂體乳液穩定性的影響
采用不同的油脂體提取方法會對油脂體提取回收率、油脂體乳液的理化特性和生產成本產生影響。目前,常用的油脂體的提取方法主要包括水相提取法、緩沖溶液提取法和酶輔助水相提取法三種[8]。提取油脂體時的一個重要決定是確定需要純油脂體還是油脂體和外源蛋白質的混合物[27]。不同提取方法提取的油脂體穩定性存在差異,這主要歸因于不同提取方法導致油脂體表面外源蛋白和酶的不同。擁有植物儲存蛋白的油脂體混合物對于其穩定性可能是有益的[28]。一般來說清洗越完全的油脂體物理穩定性較低,因為油體蛋白的分子量非常低,而且蛋白質親水性部分平鋪在界面上,近距離相互作用的兩個油體蛋白之間的排斥力相對較弱,油脂體相對不穩定[29]。
通過比較三種提取方法得到的油脂體理化性質差異,發現酶法提取油脂體的ζ-電勢和粒徑顯著高于水相法和緩沖溶液法,說明酶法提取油脂體乳液物理穩定性較強;緩沖溶液法提取的油脂體乳液過氧化值顯著高于酶法和水相法,酶法和水相法提取的油脂體的過氧化值無顯著差異,說明酶法與水相法提取油脂體乳液的氧化穩定性較強。田其英等[30]研究發現較水提法提取油脂體,酶和蔗糖輔助提取的油脂體的乳化活性較好,這可能是由于油脂體吸附蛋白量減少和磷脂表面積的增加。蔗糖輔助提取油脂體的乳化穩定性較好,這說明一定比例的脂質和蛋白質存在可以使油脂體保持較好的乳化穩定性。Karkani等[31]研究了不同回收方法對油脂體氧化穩定性的影響,經超濾得到的油脂體乳濁液具有較強的抗氧化穩定性。Nikiforidis等[19]發現將水相提取法與超濾相結合提取的油脂體粒徑較小,不易相互聚集融合,提高了油脂體的穩定性和完整性。
2.2 環境條件對油脂體乳液穩定性的影響
對油脂體乳液穩定性產生影響的環境因素主要是溫度和pH。Yan等[32]發現在熱處理過程中,大豆油脂體的大小主要由其表面蛋白決定。熱處理可以通過影響油脂體相關蛋白的構象而對油脂體乳液的穩定性產生影響[33]。經過熱處理的油脂體乳液在儲存過程中可能更穩定,原因是其中的酶或微生物已被滅活,有利于油體蛋白保留在油脂體上[34]。例如研究發現低溫熱處理對油脂體乳液ζ-電位和表面蛋白質的二級結構無明顯影響,并且降低了乳液的粒徑,避免油脂體顆粒聚集的情況,提高油脂體的物理穩定性,并且通過降低乳液中脂肪氧化酶的活性,提高了油脂體乳液的氧化穩定性;而高溫熱處理使油脂體表面蛋白變性,二級結構發生改變,降低了油脂體的絮凝穩定性并且使油脂體乳液氧化加速[35]。油脂體乳液在等電點時穩定性最差,因為此時表面電荷量最少,油脂體容易發生凝結失穩。偏離等電點時,油脂體表面靜電排斥逐漸增大,較為穩定。并且隨著pH的增加,外源性蛋白會從油脂體上逐漸解離下來,從而影響油脂體的穩定性[36]。Qi等[37]發現在堿處理下,油脂體相關蛋白的β-折疊結構的含量增加,這有助于大豆油脂體的聚集。表明油脂體相關蛋白構象的變化可能對不同p H下油脂體的穩定性起重要作用。表1列舉了部分環境條件對油脂體穩定性影響的研究進展。

表1 環境條件對油脂體穩定性的影響研究進展Table 1 Research progress on the influence of environmental conditions on the stability of grease body
2.3 離子強度對油脂體乳液穩定性的影響
高鹽離子濃度會加速油脂體乳液的聚集和失穩,低鹽離子濃度則有助于維持其穩定性。鹽離子通常是通過電荷的屏蔽效應來調節乳液體系的穩定性[36]。離子對油脂體乳液穩定性影響機制如圖5所示。
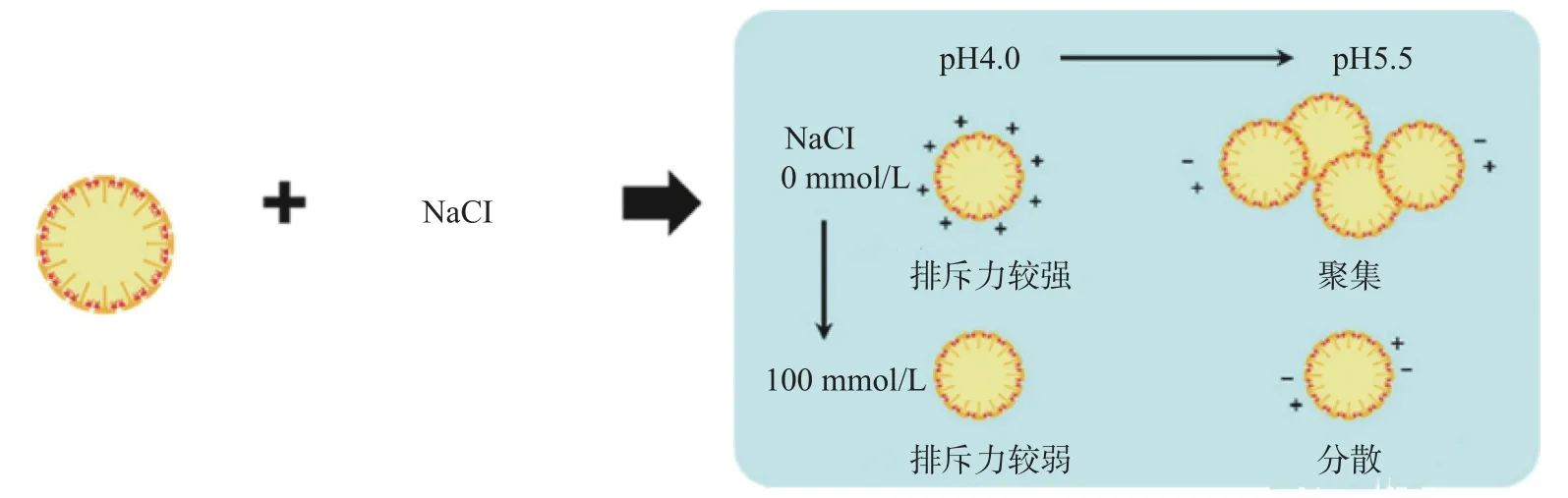
圖5 離子對油脂體的影響機理[41]Fig.5 Mechanism of ions'influence on oil body[41]
Maria等[45]研究發現,含單價陽離子(Na+、K+)的油脂體乳液表現出輕微的聚集,而含相同濃度二價陽離子(Ca2+、Mg2+)的油脂體則廣泛聚集。二價陽離子可能與油脂體膜蛋白相互作用,使其重新配置,抑制蛋白質的疏水相互作用從而對油脂體的穩定性有顯著的負面影響。另外,Kapchie等[39]發現在相同p H時含氯化鐵油脂體乳液的油脂氧化產物較不含氯化鐵油脂體乳液高出數倍,表明鐵離子的存在可以加速油脂體中脂質的降解。
2.4 表面活性劑對油脂體乳液穩定性的影響
在油脂體乳液體系中,表面活性劑的存在不僅會增加靜電斥力,還會破壞油脂體表面蛋白質之間的疏水相互作用(如圖6),從而抑制油脂體液滴的聚集和使油脂體顆粒直徑減小,從而達到穩定油脂體乳液的作用。

圖6 表面活性劑與蛋白質的相互作用方式[46]Fig.6 Interaction modes between surfactants and protein[46]
Nikiforidis等[47]研究了三種外源性生物聚合物或表面活性劑(酪蛋白酸鈉、吐溫80和黃原膠)對不同油脂體穩定性的影響。研究發現,吐溫80、酪蛋白酸鈉或黃原膠的存在對乳液穩定性有明顯的積極影響,其中吐溫80提高油脂體抗凝聚穩定性能力最強。吐溫80或酪蛋白酸鈉加入大豆油脂體乳液中均可提高其乳化穩定性,吐溫80在一定程度上也可有效改善榛子和芝麻油脂體乳液的乳化穩定性。油脂體乳液中吐溫80濃度大于0.75%時,油脂體乳化穩定性和分散穩定性顯著提高。這種改善主要與表面活性劑吐溫80與油脂體膜表面蛋白的競爭置換有關,在油滴表面形成了親水性更高的磷脂-表面活性劑-蛋白膜,表面活性劑主要置換了油脂體膜表面的甾醇蛋白和油體鈣蛋白,置換的蛋白質部分約占最初吸附蛋白質的40%[48]。Sukhotu等[49]研究發現陰離子表面活性劑SDS在低pH下可以通過打破蛋白質之間的疏水相互作用并誘導蛋白質-SDS復合物的形成,提高靜電排斥來提高油脂體的穩定性 。
2.5 食物成分對油脂體乳液穩定性的影響
近年來,有學者們發現食品中的一些成分例如NaCl、蛋白質、酚類和大分子多糖類物質會對油脂體乳液的穩定性產生影響。
食品中的NaCl可能會對油脂體的穩定性產生負面影響,這是因為油脂體膜表面的蛋白質帶負電,當遇到帶正電的Na+離子時易發生結合,從而中和了油脂體表面蛋白所帶的電荷,降低油脂體液滴間的靜電斥力,使油脂體易發生聚集。隨著NaCl濃度的增加,油脂體表面蛋白被中和的電荷增加,油脂體越不穩定。Hou等[50]研究發現NaCl濃度為50 mmol/L時乳液的乳化活性和乳化穩定性顯著降低,隨著NaCl濃度的增加,乳液絮凝現象加重。
在酸性和中性條件下,酚類可能與蛋白質形成非共價鍵,例如氫鍵和疏水作用力[51],從而導致油脂體的聚集。多酚還可能會改變蛋白質的等電點,影響油脂體乳液的穩定性。此外,多酚能夠有效清除乳液中的自由基和螯合過渡金屬,從而提高油脂體的氧化穩定性。Ding等[52]研究了表沒食子兒茶素沒食子酸酯(EGCG)對大豆油脂體氧化穩定性的影響,研究發現在p H5和7時,EGCG對油脂體乳液的長期穩定有促進作用,但在p H 9時,EGCG并沒有提高油脂體乳液的穩定性,相反導致了油脂體的聚集。EGCG在p H 7時表現出明顯提高油脂體氧化穩定性的能力,而在p H 5時,脂質氧化反應速率加快。
近年來,人們逐漸開始將大分子多糖如黃原膠、阿拉伯膠、卡拉膠和果膠等用于提高油脂體乳液穩定性的研究中。帶電的多糖可以與油脂體表面蛋白相互作用,通過改善空間排斥力,減少絮凝損耗和形成第二“膜”來提高油脂體乳液穩定性[53],多糖包裹油脂體原理如圖7所示。Nikiforidis等[54]發現加入0.1%黃原膠后,油脂體乳液的穩定性顯著提高,乳液pH在6時表現出最高的穩定性。Su等[55]研究了陰離子多糖海藻酸鈉(ALG)對大豆油脂體乳液穩定性的影響。研究發現,ALG吸附在油脂體表面上并形成較厚的帶電涂層,因此較低濃度的ALG(0.25%~0.35%)可以提高油脂體的抗聚集穩定性,但是高濃度的ALG 下,油脂體乳液穩定性下降,這可能是由于連續相中高濃度的非吸附ALG引起的絮凝損耗所致,ALG對油脂體的作用原理如圖8所示。在中性條件下,用ALG層覆蓋大豆油脂體可以極大地擴展穩定p H范圍,大大提高了油脂體乳液的穩定性,并且ALG涂層在凍融循環過程中也起到保護油脂體的作用[56]。Wu等[57]研究發現卡拉膠能顯著提高大豆油脂體乳液的鹽穩定性,對其在食品中的應用具有重要意義。
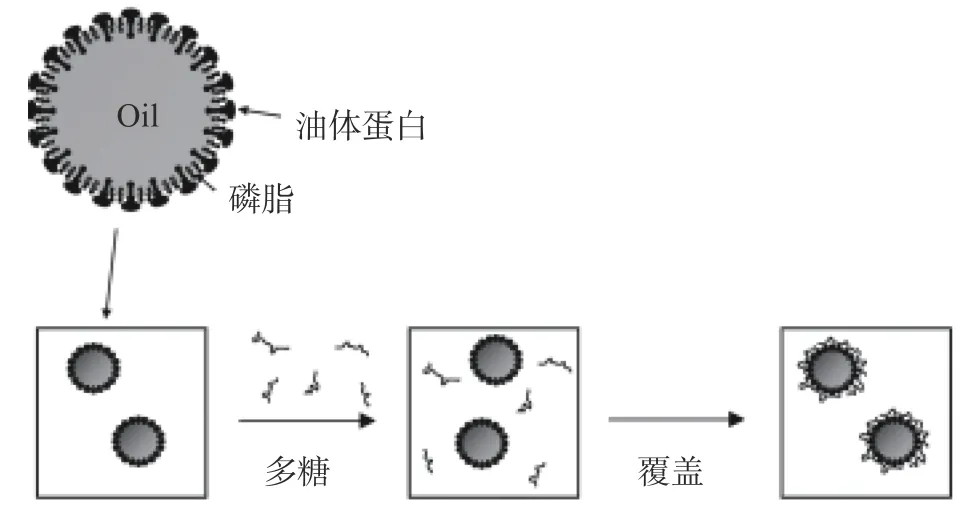
圖7 多糖包裹油脂體原理圖[22]Fig.7 Schematic diagram of coating of oil bodies by polysaccharide[22]
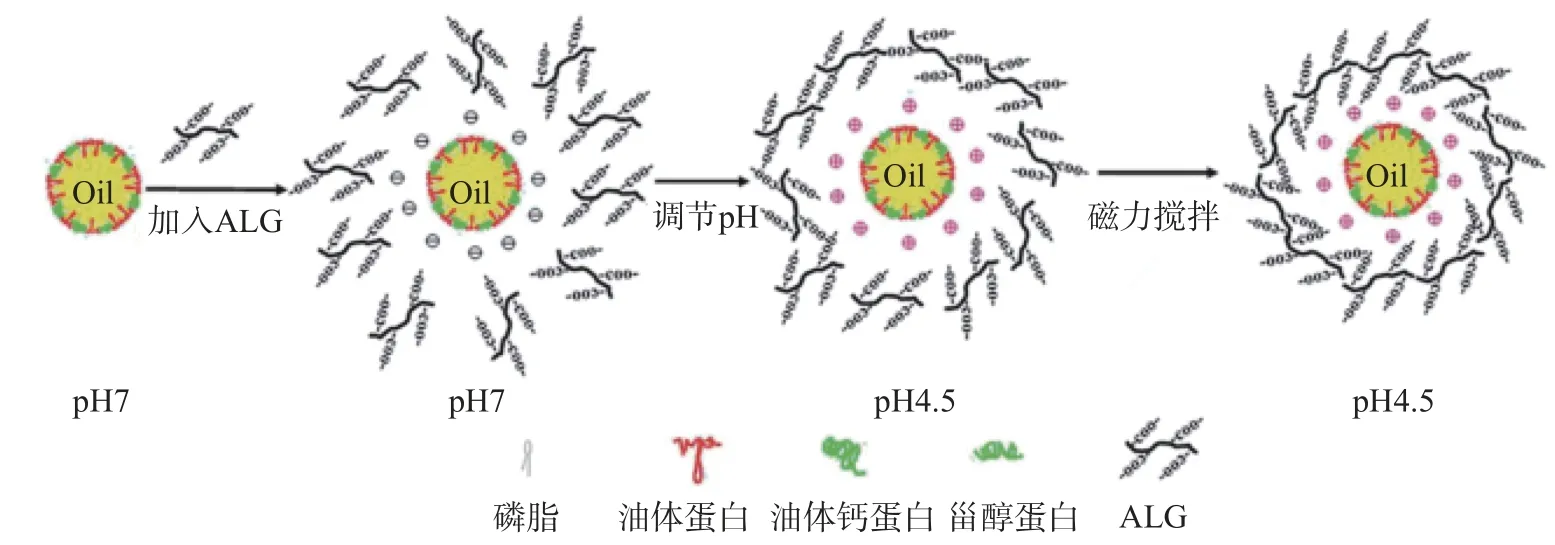
圖8 ALG作用油脂體原理[55]Fig.8 Schematic description of coating ALG on oil body[55]
2.6 不同油料作物對油脂體乳液穩定性的影響
除上述影響因素外,從不同油料作物中提取的油脂體的乳液穩定性也存在一定的差異,這可能與不同油料作物油脂體結構及組成不同有關。不同來源天然油脂體表面蛋白質和磷脂的組成存在差異,由于蛋白質疏水性不同導致油脂體乳液穩定性的不同。卵磷脂與蛋白質表面疏水殘基的疏水相互作用更強,暴露的疏水殘基越多,乳液越穩定[58]。
姜夢婷等[59]研究發現,在60℃加速氧化的條件下大豆、花生和葵花籽油脂體乳液的氧化穩定性順序為大豆油脂體>葵花籽油脂體>花生油脂體。梁新婷等[60]比較了高油大豆和低油大豆油脂體的穩定性差異,發現高油大豆油脂體乳液的乳化穩定性顯著高于低油大豆油脂體,而氧化穩定性顯著低于低油大豆油脂體。
3 油脂體的應用
3.1 作為食品乳化劑
由于油脂體膜表面的蛋白質是具有親水性和疏水性的兩親性結構,油脂體可以作為天然乳化劑應用于食品中。根據目前研究,油脂體作為乳化劑可以應用于蛋黃醬、冰淇淋、千島醬、布丁和果汁等中。大豆油脂體作為乳化劑適用于高鹽低酸性食品,如千島醬、蛋黃醬和沙拉醬等中[61]。例如,將大豆油脂體部分替代蛋黃制備蛋黃醬,油脂體蛋黃醬蛋白質含量顯著高于普通蛋黃醬而膽固醇含量卻顯著低于普通蛋黃醬,且油脂體蛋黃醬與普通蛋黃醬相比具有更好的貯藏穩定性,普通蛋黃醬和油脂體蛋黃醬在外觀形態和氣味上無顯著差異[62]。利用天然的大豆油脂體替代傳統沙拉中的油脂,制成的沙拉具有含油量低、脂肪消化慢、營養素天然健康,且儲存穩定等優點[63]。將油脂體乳化劑用于果汁中,不僅可以使果汁更加濃稠并且可以防止固體成分的沉淀。在制作冰淇淋過程中,可以添加油脂體乳化劑以替代價格昂貴的非脂乳固體,不僅保持了冰淇淋原有口感還降低了冰淇淋的制作成本[64]。由于大豆油脂體的良好性狀,也可將其應用到乳制品、仿制牛奶、酸奶和奶油等產品中。與精煉大豆油相比,大豆油脂體由于富含油脂伴隨物,如磷脂、維生素E和植物甾醇等,并且卡路里要大大低于從牛奶中分離出的奶油,可作為一種新型的油脂產品進行深入研究和應用[65]。將卵磷脂和花生油脂體的組合物應用于可可脂及代可可脂中,該組合可以降低產品的黏度,保持產品質地和口感,提高了產品的抗氧化性,降低生產成本[66]。
3.2 作為載體
由于油脂體膜的特殊性質,可將疏水性分子截留并保護在油脂體TAG核中。將生物活性物質包封在油脂體中進行噴霧干燥可以有效提高生物活性物質的抗氧化穩定性和生物可及性,并且經噴霧干燥制成的固體產品相比于液態形式在運輸和儲存過程中更方便、安全。例如,蝦青素是一種非常容易氧化的分子,可被包封在油脂體核心中,包封的蝦青素很好的保持了其抗氧化活性[67]。姜黃素由于其疏水性,生物利用度較差。研究發現利用納米級油脂體包封姜黃素顯示出高封裝效率和負載能力[68]。利用該性質還可以將油脂體應用于污水中有毒有機分子的去除,當油脂體被放置在有機農藥污染的水環境中,有機分子可以被吸收到油脂體的內部疏水性核中并被去除[69]。
4 結語
油脂體主要以乳狀液的形式應用于食品、藥品、飼料及個人護理產品等領域中。隨著對天然油脂體研究和開發的不斷深入,人們發現油脂體乳液的穩定性與油脂體的組成、提取方法、外源蛋白質、溫度、p H值和離子強度等條件有關。這些影響因素通過改變油脂體表面電荷量和表面張力,形成非共價鍵,鈍化脂肪氧化酶活性,或清除自由基和螯合過渡金屬等機制來影響油脂體乳液的理化穩定性。油脂體膜表面的蛋白質是維持油脂體穩定性的關鍵。要深入了解油脂體膜中的分子相互作用對于油脂體的實際生產應用,探索新的潛在應用非常重要。然而,目前有關油脂體相關蛋白結構與功能關系及穩定功能的研究仍然有限,有待于今后的深入研究。總之,隨著對油脂體認識的逐步深入及其在相關行業應用的不斷擴大,油脂體產品將逐漸出現在人們的生活中。
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
