組蛋白H3K27me3介導JAK2/STAT3信號通路對足細胞損傷的調控機制*
2021-09-08 04:01:32袁樹珍隋曉露顧鳳娟張艾莎許云鵬謝婷妃曾啟城鄒杰鋒陳繼紅
醫學理論與實踐 2021年17期
袁樹珍 隋曉露 顧鳳娟 張艾莎 許云鵬 謝婷妃 曾啟城 鄒杰鋒 陳繼紅
廣東醫科大學深圳寶安臨床醫學院,廣東省深圳市 518100
足細胞損傷是腎小球疾病發生發展的重要機制。表觀遺傳修飾參與足細胞損傷的過程,組蛋白甲基化水平變化導致炎性反應基因轉錄過程及多個相關信號通路活化異常,破壞足細胞正常結構及其功能,導致足細胞凋亡[1]。組蛋白甲基化轉移酶同源序列增強子2(Enhancer of zeste homolog 2,EZH2)催化組蛋白H3K27發生三甲基化,EPZ-6438是EZH2的抑制劑,降低組蛋白H3K27me3水平,抑制炎性基因和超氧化物歧化酶表達,減輕炎性反應和延緩腎小球疾病的進展[2]。JAK2/STAT3信號通路是細胞內參與凋亡及炎癥信號轉導的重要通路,能被多種代謝產物激活,引起炎癥因子表達增多,促進足細胞凋亡,在多種腎臟疾病中發揮重要作用[3]。JAK2/STAT3信號通路活化可促進EZH2的表達,并且STAT3與EZH2的啟動子區相互結合,參與腫瘤細胞增殖等生理過程。本研究選用小鼠腎足細胞MPC5,給予EPZ-6438干預處理,觀察足細胞組蛋白H3K27me3及JAK2/STAT3信號通路水平變化,探討組蛋白H3K27me3介導JAK2/STAT3信號通路對足細胞損傷的調控機制。
1 材料與方法
1.1 細胞與試劑 小鼠腎足細胞MPC5,購自深圳市拓普生物科技公司。DMEM培養基,EZH2抑制劑(EPZ-6438),RNA反轉錄試劑盒(購自ThermoFisher公司),BCA蛋白濃度測定試劑盒,RIPA裂解液。
1.2 細胞培養及分組 小鼠腎足細胞MPC5用10%胎牛血清(FBS)、1%青鏈霉素混合液DMEM培養基培養,放置37℃、5%CO2培養箱,當足細胞密度達到90%以上時收集細胞。PBS潤洗,加入1ml 0.25%胰酶消化處理,按1/3比例傳代,傳2個10cm2平皿放置37℃、5%CO2培養箱過夜。隨機分對照組及EZH2-組。EZH2-組給予EPZ-6438 10μm 培養48h。對照組給予等量PBS培養48h。
1.3 Western Blot法檢測足細胞組蛋白H3K27me3 收集細胞,加RIPA裂解液裂解30min,離心取上清液。BCA蛋白濃度測定試劑盒檢測總蛋白量,調整蛋白濃度為6μg/μl,上樣電泳,電壓80V跑膠25min,160V跑膠50min。取出蛋白凝膠進行轉膜(200mA,2h)。脫脂牛奶封閉后加入H3K27me3抗體(1∶1 000)4℃孵育過夜,TBST緩沖液洗膜,采用Image J軟件分析靶蛋白帶的灰度水平。
1.4 qPCR 法檢測足細胞JAK2、STAT3mRNA 收集細胞,用Buffer RZ提取足細胞總RNA,參照RNA反轉錄試劑盒(購自ThermoFisher公司)說明書合成cDNA。應用實時熒光定量PCR儀進行PCR擴增, 引物序列見表1,實時定量PCR反應條件為:95℃預變性15min,95℃變性10s、60℃退火延伸30s、72℃延伸30s共45個循環。以β-actin為內參照,采用2-ΔΔCT法進行定量分析,實驗重復3次,取平均值。

表1 JAK2、STAT3的引物序列
1.5 Western Blot法檢測足細胞p-JAK2、JAK2、p-STAT3、STAT3蛋白 收集細胞,加RIPA裂解液裂解30min,離心取上清液。BCA蛋白濃度測定試劑盒檢測總蛋白量,調整蛋白濃度為6μg/μl,上樣電泳,電壓80V跑膠25min,160V跑膠50min。取出蛋白凝膠進行轉膜(200mA,2h)。脫脂牛奶封閉后加入一抗(p-JAK2、JAK2、p-STAT3、STAT3、內參抗體,1∶1 000)4℃孵育過夜,TBST緩沖液洗膜。加入二抗(1∶10 000)室溫2h后進行顯色,采用Image J軟件分析靶蛋白帶的灰度水平。
1.6 ChIP-chip技術檢測組蛋白H3K27me3靶基因及富集分析 (1)文庫構建:收集兩組處于對數生長期的細胞,使用免疫磁珠結合細胞完成分選,使用ChiTag促進H3K27me3抗體與目標蛋白結合。Trizol法提取兩組DNA,使用Nanodrop檢測DNA的純度,使用Qubit對DNA進行定量。對目標DNA進行PCR擴增,修復DNA片段及添加測序接頭后完成純化,完成文庫構建。(2)文庫質檢和測序:使用Agilent 2100 對文庫的插入片段長度進行檢測,排除接頭二聚體污染,使用q-PCR 對測序文庫濃度進行定量,保證樣品純度,文庫質檢合格后于Illumina平臺測序獲取靶基因。(3)組間差異富集峰統計:使用MACS/PePr軟件進行組間富集峰Peak數目統計。(4)數據分析:利用MACS2軟件對兩組樣品進行富集分析(閾值設定為P≤0.05),將富集峰與基因結構元件進行基因組位置比較,確定與富集峰有直接關系的基因集進行基因注釋。(5)GO富集分析:將富集峰相關基因集通過向GO數據庫各Term形成映射,完成Term定位,應用超幾何檢驗得到在生物過程、細胞組成和分子功能方面富集的GO項目上Peak重疊基因的條目和分布情況。(6)KEGG富集分析:將富集峰相關基因集通過KEGG數據庫,應用超幾何檢驗得到在差異表達基因中顯著性富集的信號通路,并確定信號通路中呈現顯著性富集的基因靶點。
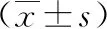
2 結果
2.1 足細胞組蛋白H3K27me3水平 EZH2-組足細胞組蛋白H3K27me3蛋白表達水平為0.85±0.12,對照組為1.25±0.08,EZH2-組顯著低于對照組,差異有統計學意義(P=0.007<0.05)。見圖1。
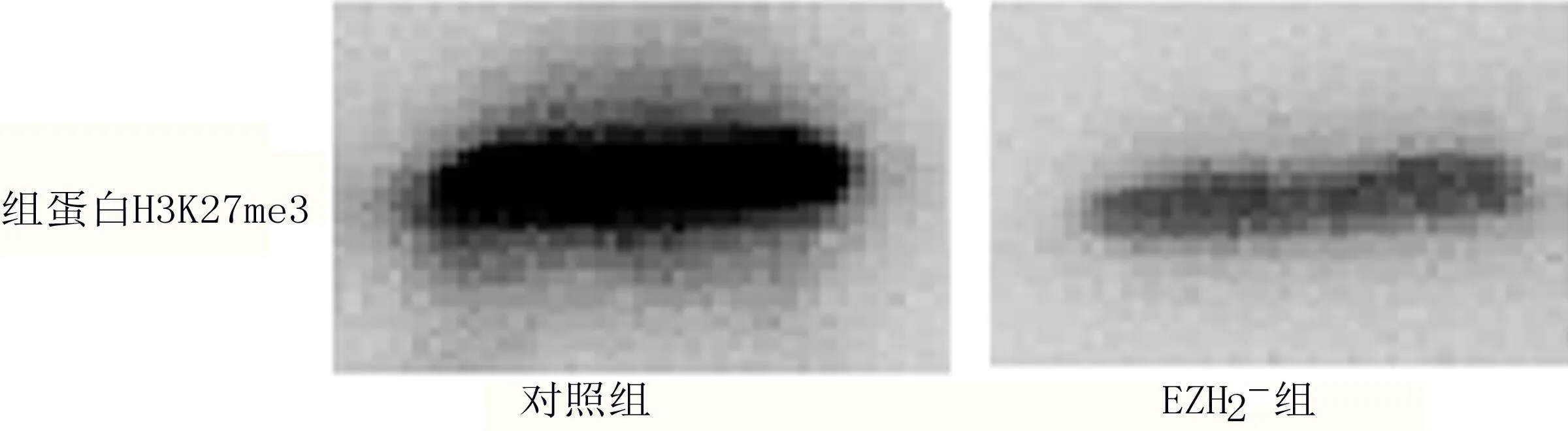
圖1 組蛋白H3K27me3蛋白表達Western Blot電泳圖
2.2 JAK2/STAT3信號通路目標基因mRNA水平 與對照組相比,EZH2-組足細胞JAK2、STAT3mRNA表達顯著增加,差異有統計學意義(P<0.05)。見表2。
表2 兩組足細胞JAK2、STAT3 mRNA水平
2.3 JAK2/STAT3信號通路目標蛋白水平 與對照組相比,EZH2-組足細胞JAK2、p-STAT3、STAT3蛋白表達顯著增加,差異有統計學意義(P<0.05)。見表3,圖2。
表3 兩組足細胞p-JAK2、JAK2、p-STAT3、STAT3蛋白水平

圖2 JAK2/STAT3信號通路目標蛋白表達Western Blot電泳圖
2.4 組間差異富集峰統計 組間差異富集峰Peak數目統計發現EZH2-組有6 919條富集峰,對照組有3 273條富集峰,兩組各列舉前15條。見表4。
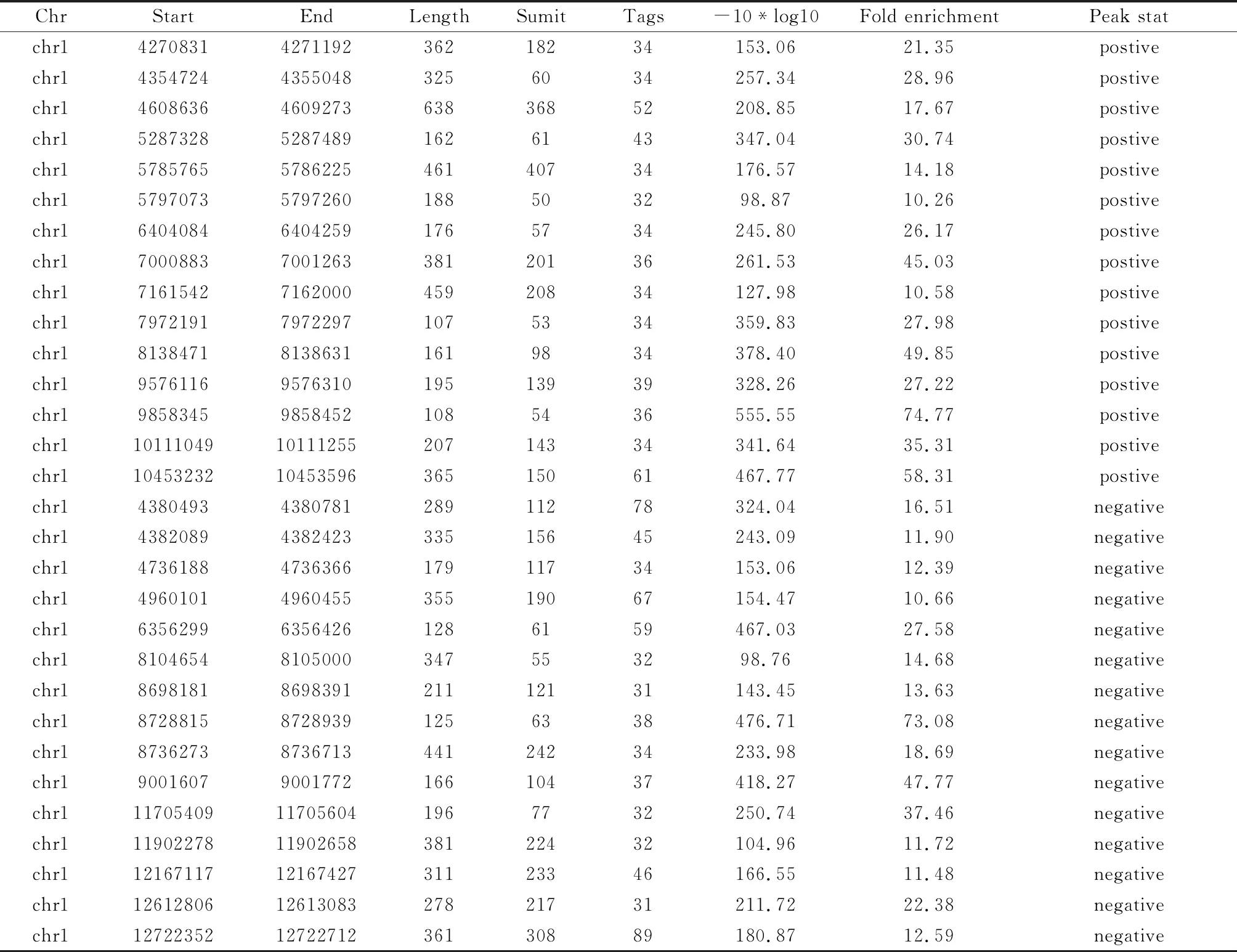
表4 兩組間差異富集峰(Peak)統計
2.5 組蛋白H3K27me3調控靶基因GO富集分析 靶基因功能主要集中在生物過程、細胞組成、分子功能方面(P≤0.05)。見表5,圖3。其中涉及生物過程中Pigu基因呈現顯著性富集,其參與JAK/STAT信號通路調控。見表6。

表6 JAK/STAT信號通路靶基因GO富集分析
2.6 組蛋白H3K27me3調控靶基因KEGG富集分析 靶基因參與調控信號通路主要集中在細胞溶質 DNA 感應途徑、Rap1信號通路、Toll 樣受體信號通路、HIF-1 信號通路和 JAK/STAT 信號通路。見表7,圖4。其中靶基因IL12rb1、IL5ra呈現顯著性富集,其參與JAK/STAT 信號通路調控。見表8。

表8 JAK/STAT信號通路靶基因KEGG功能富集分析
3 討論
足細胞損傷是腎小球疾病發生進行性腎小球硬化和腎纖維化的重要因素,其發病機制包括[4-5]:(1)血流動力學變化使血管緊張素Ⅱ(AngⅡ)水平增加改變足細胞蛋白表達及分布直接損傷足細胞;(2)RAS系統激活可通過促進鈣離子內流和活性氧生成導致足細胞損傷;(3)氧化應激反應通過擾亂內質網穩態,激活足細胞內質網應激系統導致足細胞損傷;(4)表觀遺傳修飾調控基因轉錄過程導致炎性反應基因及多個相關信號通路活化促進足細胞凋亡等。其中,表觀遺傳修飾在足細胞損傷機制具有重要作用。
表觀遺傳修飾是指非基因序列改變所致基因表達水平的變化,包括DNA甲基化、組蛋白甲基化、非編碼RNA等。組蛋白甲基化是發生在N端H3和H4氨基酸殘基上的甲基化,由組蛋白甲基轉移酶催化完成,維持染色質結構和調控基因轉錄,在足細胞損傷中發揮著重要作用。EZH2是EZH基因編碼的組蛋白賴氨酸甲基化轉移酶,是多梳抑制復合體2(PRC2)的一個催化亞基,聚集至啟動子區域,催化組蛋白 H3K27發生三甲基化抑制靶基因轉錄及調控下游炎癥反應信號通路活化過程[6]。EZH2活性減弱使組蛋白H3K27me3水平下降,對microRNA抑制作用減弱,由microRNA調控的抗氧化劑抑制劑硫氧還蛋白互作蛋白(TxnIP)表達增加,機體產生氧化應激損傷足細胞,提示組蛋白H3K27me3修飾水平變化參與足細胞損傷過程[7]。
足細胞損傷與多條信號通路異常活化相關,如p38MAPK信號通路、TGF-β/Smad信號通路、 NLRP3炎癥小體信號通路[8],其中JAK2/STAT3信號通路在足細胞分化、凋亡過程中起重要作用。JAK2是非受體型胞漿酪氨酸蛋白激酶,通過與受體偶聯介導細胞因子相關信號的傳遞。STAT3是DNA結合蛋白,與磷酸化酪氨酸的肽段結合被磷酸化形成二聚體,進入胞核內與靶基因啟動子序列的特定位點結合,激活基因轉錄[9]。體內p-STAT3過表達可上調EZH2活性顯著增強組蛋白H3K27me3表達水平,抑制抑癌基因的表達,促進腫瘤浸潤轉移,提示組蛋白H3K27me3可能通過介導JAK2/STAT3信號通路參與細胞分化、凋亡過程。

圖4 組蛋白H3K27me3調控靶基因KEGG分類
本研究結果顯示:與對照組相比,EZH2-組足細胞H3K27me3蛋白表達顯著下降(P<0.05);JAK2/STAT3信號通路目標基因JAK2、STAT3mRNA 表達顯著增加(P<0.05);JAK2/STAT3信號通路目標蛋白JAK2、p-STAT3、STAT3蛋白表達顯著增加(P<0.05);組蛋白 H3K27me3 調控靶基因 GO 富集分析發現靶基因功能主要集中在生物過程、細胞組成、分子功能方面,其中涉及生物過程中 Pigu 基因呈現顯著性富集,其參與JAK/STAT信號通路調控;組蛋白H3K27me3 調控靶基因 KEGG 富集分析發現靶基因參與調控信號通路主要集中在細胞溶質 DNA 感應途徑、Rap1 信號通路、Toll 樣受體信號通路、HIF-1 信號通路和 JAK/STAT 信號通路,其中靶基因IL12rb1、IL5ra呈現顯著性富集,其參與JAK/STAT 信號通路調控。組蛋白H3K27me3介導 JAK2/STAT3信號通路致足細胞損傷的可能機制如下:(1)p-STAT3屬于核轉錄因子,介導細胞內基因表達過程。EZH2是PRC2中的一個催化亞基,聚集至啟動子區域。在各種刺激因素下,足細胞內啟動子區域EZH2的核定位能力減弱及其活性下降,細胞內組蛋白H3K27me3水平下降,使其染色體結構處于疏松狀態,增強了p-STAT3與基因啟動子的結合,對TGF-β基因等促纖維化關聯基因轉錄抑制作用減弱[10], 與腎臟纖維化相關的炎癥介質及細胞因子如血小板源生長因子(Platelet-derived growth factor)、IL-1β、IL-18等產生增多,導致足細胞損傷。(2)白細胞介素5受體(IL5ra)基因編碼IL5受體的α鏈,該受體依賴腫瘤壞死因子(TNF-α)激活相關信號通路參與細胞凋亡過程[11]。炎癥因子TNF-α與IL5受體結合激活JAK/STAT通路,執行細胞外基質重塑和細胞凋亡。(3) 磷脂酰肌醇多糖錨定生物合成類(Pigu)基因編碼糖基磷脂酰肌醇轉酰胺酶(GPI-T)復合物的第5亞基,GPI-T主要位于內質網并催化GPI錨定蛋白合成,GPI錨定蛋白是一類由蛋白質、脂質和糖鏈共價結合的復雜糖復合物,參與信號轉導過程。Pigu基因過度表達能上調GPI錨定蛋白水平激活JAK/STAT通路,參與細胞增殖過程[12]。(4) Pigu、IL5ra、TGF等多個基因調控JAK2/STAT3信號通路活化過程[13]。代謝產物與酪氨酸激酶相關受體結合激活JAK2/STAT3信號通路,組蛋白H3K27me3水平下降對靶基因抑制作用減弱,Pigu 、IL5ra、TGF等多個基因轉錄活性增強促進JAK2酪氨酸磷酸化作用增強,JAK2激活后催化受體酪氨酸殘基發生磷酸化修飾與周圍的氨基酸序列形成停泊位點(docking site),募集更多含有SH2結構域的STAT3蛋白到這個停泊位點。STAT3蛋白發生磷酸化修飾水平明顯增強進入細胞核內與促纖維化等基因結合,上調TGF-β1、α-SMA等蛋白的表達,造成細胞外基質堆積及足細胞損傷[14]。
本細胞實驗給予組蛋白甲基化酶抑制劑干預,足細胞組蛋白 H3K27me3 表達下調,JAK2/STAT3信號通路表達增強。經組蛋白 H3K27me3靶基因富集分析提示組蛋白 H3K27me3通過調控靶基因Pigu、IL12rb1、IL5ra參與JAK2/STAT3信號通路活化,導致足細胞損傷。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
中國生殖健康(2019年3期)2019-02-01 06:12:26
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
學苑創造·A版(2015年11期)2016-01-14 09:03:27
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
中國火炬(2010年8期)2010-07-25 11:34:30
