石櫟屬物種果實類型及其系統演化研究進展
2021-09-11 02:46:50覃元藝
云南民族大學學報(自然科學版) 2021年4期
覃元藝,陳 曦,2,3
(1.云南大學 生態與環境學院,云南 昆明 650091;2.云南大學 云南省植物繁殖適應與進化生態學實驗室, 云南 昆明 650091;3.云南大學 云南省高原山地生態與退化環境修復重點實驗室,云南 昆明 650091)
殼斗科(Fagaceae)包括12個屬 1 000 余種,是北半球熱帶、亞熱帶及溫帶森林的主要建群樹種,也是最重要的木本植物大科之一[1-2].石櫟屬(LithocarpusBlume)是殼斗科內物種多樣性極為豐富的重要大屬,其320多個種廣布于東亞、東南亞及喜馬拉雅南麓中低海拔河谷[1-4].我國和東南亞是石櫟植物多樣性的2個重要分布中心.石櫟不僅是熱帶亞熱帶常綠闊葉林林冠層的主要樹種和建群種[5-7],其種子也是這些地區眾多動物類群的食物和營養來源[7-10].因此石櫟對維持這些森林系統的碳匯平衡和生態多樣性起著重要作用.
對比同科的櫟屬(QuercusL.),石櫟屬獲得的研究關注相對較少.這首先是因為櫟屬植物在北半球廣泛分布,而很多西方研究者并不熟悉分布于東亞的石櫟.另外,櫟屬植物是重要的經濟樹種,而此前石櫟屬植物由于木質較差被認為經濟價值不高.但最近幾十年來,石櫟屬植物的經濟開發價值受到關注,如食用[11-12]、藥用[13-18]、造林綠化[19-23]等多個方面.可見,除了具有重要生態價值之外,石櫟也兼顧多項潛在的經濟價值.在經濟利益的驅動下,人們開始從自然種群中采集和移栽石櫟物種,但這種缺乏科學管理的采集造成了石櫟種群的過度開發和森林群落生物多樣性的喪失,并在未來可能帶來生物入侵和污染問題,甚至影響全球氣候變化[6, 24-26].
因此,為石櫟資源的科學合理開發提供理論依據,必須首先深化石櫟的基礎研究.在眾多方向的基礎研究之中,以石櫟果實形態和類型為主線拓展出的研究極具特色.由于石櫟物種果實形態變異豐富(見圖1),圖中黑色直線代表 1 cm 長度.因部分石櫟物種在果實成熟時殼斗與果實分離,因此殼斗結構缺失,未能標注.因此其果實形態學成為理解其物種多樣性和進化的關鍵.通過將石櫟的果實形態和果實類型研究與植物生理、形態發育、生物地理、種子散播和物種更新以及分子系統發育等方面貫通融合,得以對石櫟的分布、物種進化和共存、物種和形態多樣性、物化特性與生活史對策等一系列問題有了初步的理解.因此本文歸納了最新的石櫟屬植物的果實形態多樣性與物種分化研究,探討了石櫟果實形態和類型與其分布、物種多樣性、物種更新和分子系統發育之間的重要聯系和科學意義.同時探討石櫟研究過程中存在的問題與重要的研究空白,對未來石櫟基礎研究方向提出建議.
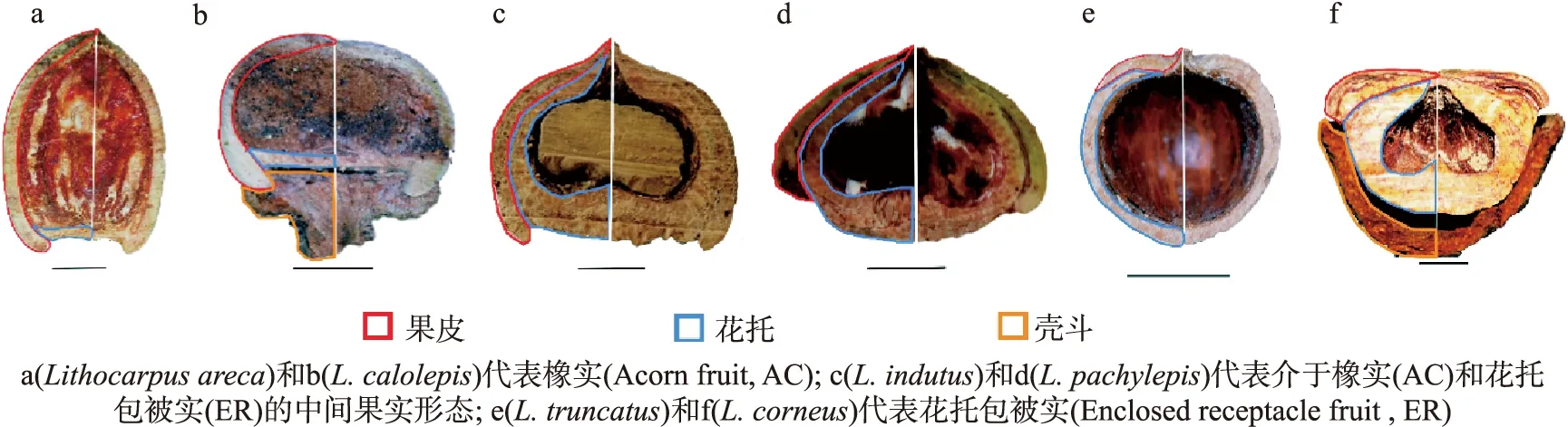
圖1 6個石櫟物種果實的縱截面形態結構
1 石櫟果實形態多樣性與物種分化研究
1.1 石櫟屬2種主要果實類型的形態和物化權衡
由于石櫟物種的無性結構(如枝、葉等)分化不明顯[28],因此形態變異豐富的果實和殼斗被作為物種鑒定的重要證據廣泛應用于傳統分類學中[29-31].石櫟物種單個的堅果果實被杯狀殼斗結構所包被[32](見圖1b、1f).殼斗是殼斗科植物的重要特征,是無性枝融合后形成的扁平狀或碗狀結構[33].殼斗內是單個的石櫟堅果,其果殼由果皮和花托組成[27](見圖1).
早在20世紀中期,法國科學家Camus就注意到石櫟物種的果實形態變異顯著(圖1).2001年,Cannon和Manos首次嘗試分析石櫟果實的形態多樣性,用elliptic fourier analysis (EFA)量化21種熱帶石櫟物種果實的花托和果皮形態差異,發現這21個物種的果實形態分化可以被歸納為2種果實類型:橡實(acorn fruit,后簡稱AC)和花托包被實(enclosed receptacle fruit,后簡稱ER)[28](圖2).圖中黑色直線代表 1 cm 長度.橡實(AC)與櫟屬植物果實(俗稱麻栗果)形態十分類似,單薄的果皮是包被其種子的主要結構,花托存在于種子基部,通常為扁平凹形(圖2a、2c);而花托包被實(ER)的果皮急劇縮短僅覆蓋于種子頂部,花托卻急劇延伸加厚成為包裹種子的主要結構(圖2d、2f).這項研究為石櫟的果實形態和果實類型研究奠定了重要基礎,但其局限性體現在(1)僅包括熱帶的21個石櫟物種,缺乏對亞熱帶石櫟和豐富石櫟物種多樣性(320多個種)的代表;(2)該研究通過肉眼判斷物種果實類型的方法缺乏數據支撐.

圖2 形態學研究對橡實(AC)(Lithocarpus kawakamii,a~c)和花托包被實(ER)(L. corneus,d~f)的果實結構理解的變化發展
為更好地體現石櫟屬層面的果實形態分化,陳曦等針對廣泛分布于中國和東南亞的98個石櫟物種(代表了石櫟屬1/3的物種多樣性),從物種果實縱截面圖像利用Pappus-Guldinus定理重建果實3D形態,量化多項果實形態特性(包括果實和種子體積、外果皮和花托對種子的包被程度等),并進行了種內和種間的果實形態變異分析.研究結果表明,石櫟物種內的形態分化較小,石櫟屬果實多態性主要由種間形態分化造成.且該研究通過對比外果皮和花托對種子的包被程度進行果實類型劃分,相較于Cannon和Manos用肉眼進行果實類型劃分的方法更加客觀準確.該研究結果支持Cannon 和Manos提出的2種主要果實類型:橡實(AC)和花托包被實(ER)代表了石櫟屬層面的果實形態多樣性,但橡實(AC)是石櫟屬更常見和普遍的果實類型(只有不到1/3的物種生產花托包被實(ER)).該研究還證明了2種果實類型存在形態和物理防御分化:花托包被實(ER)和橡實(AC)的主要物理防御結構分別是花托和果皮(圖2),花托包被實(ER)的果實和種子較大,且果實物理防御水平顯著高于橡實(AC)[7].
為理解2種果實類型其物理防御權衡的形成過程和發育差異,陳曦等分析了代表2種果實類型的20個石櫟物種在發育過程中的的多項果實形態動態特征:種子大小、外中果皮聯合體體積(由于2個組織結合緊密,故而作為一個整體進行分析)、內果皮體積和花托體積的增長變化,再現了2種果實類型從果實發育初期到成熟的動態發展過程[27].這項研究的重要性首先在于對石櫟果實結構的修正認定:此前石櫟果實形態和果實類型研究[7, 28, 35]中的被認為包被橡實(AC)的主要結構——“外果皮”(圖2b、2e),實為外、中、內果皮的總和,也就是果皮結構(圖2c、2f) (為避免概念混亂,前研究中出現“外果皮”結構在本文中均糾正為“果皮”)[27].更重要的是,該研究發現2種果實類型在果實發育早期的形態結構和組織分化順序:外中果皮聯合體首先分化出現,內果皮和花托后分化出現,最后才是種子發育;但2種果實類型在果實發育后期的組織分化差異造就了它們在成熟時的果實形態分化和物理防御權衡:果實發育后期,橡實(AC)的內果皮發育顯著,而花托包被實(ER)的花托發育顯著,于是形成了單薄的果皮代表了橡實(AC)的弱物理防御,而木質化加厚的花托是花托包被實(ER)的強物理防御[27].此外,該研究也支持Brett(1964)提出的殼斗功能假說:殼斗為處于發育初期階段的果實,尤其是橡實(AC)提供機械保護.這項修訂明確了石櫟果實結構和2種果實類型的形態差異的發育學原因,不僅為理解2種果實類型功能進化機制奠定了重要基礎[36],也為研究其他果實發育研究提供了模型范本.
除了果實形態和物理防御權衡,這2種果實類型也存在種子化學成分權衡:花托包被實(ER)種子的營養成分(粗蛋白質、脂肪、單糖和總非結構型碳水化合物)較高,而橡實(AC)種子中代表化學防御的拒食素(中性洗滌纖維、酸性洗滌纖維、纖維素、半纖維素、木質素、總單寧,縮合單寧和水解單寧)水平更高[35].出乎意料的是,橡實(AC)中主要的拒食素成分并非為人熟知的單寧[37],而是纖維素[35].部分哺乳動物的唾液中存在有單寧綁定唾液酶(tannin binding silavary proteins, TBSPs),這種酶可以抑制單寧的毒性,從而抑制單寧作為化學防御的角色.因此,研究者們猜想可能石櫟種子的捕食者唾液中存在單寧綁定唾液酶,因此石櫟種子轉而投資生產無法被動物分解的纖維素,來抑制動物對能量和營養的獲取,成為更有效的化學防御[35].
綜合以上石櫟果實類型形態分化和理化特性研究,發現石櫟2種果實類型存在一系列的形態和物化權衡:橡實(AC)的果實和種子較小,單薄的果皮是其種子的主要防御結構,因此物理防御弱,但種子內的化學防御水平高;而花托包被實(ER)的種子和果實更大,種子富含營養,而木質化加厚的花托是包裹種子的主要結構,代表了強物理防御[7, 27, 28, 35](表1).但是,需要注意的是,觀察到有一些石櫟物種代表了不便于劃分、介于2種果實類型間的中間形態(圖1c、1d):它們的花托急劇延長加厚,幾乎覆蓋了整個種子,但同時果皮也沒有縮短,而是覆蓋在花托外部,和花托共同保護種子.陳曦等也指出雖然可以根據花托和果皮對種子的絕對包被程度將這些物種劃分成橡實(AC)或者花托包被實(ER)[7],但是它們在形態上展現出來的特殊性對石櫟果實形態和進化研究有重要意義.通過對具有中間果實形態的物種進行詳細的果實理化特性分析,并與2種果實類型進行對比,是回答中間果實類型與2種果實類型關系和差異的重要環節,也是理解石櫟果實類型進化的關鍵.

表1 石櫟屬兩種果實類型形態和理化特性對比
1.2 2種果實類型的地理分布特性
為了探究石櫟果實形態多樣性與其地理分布的內在聯系,陳曦等人調查了93個石櫟物種的分布情況,發現代表2種果實類型的石櫟物種的經緯度分布有很大的重疊性,共享中國和東南亞2個主要分布中心[7].從群落層面上看,2種果實類型的石櫟物種也常常共生[35, 38-40],甚至具有相似的優勢度[7].廣義線性分析顯示石櫟種子隨海拔升高和緯度降低而變大,且石櫟果實物理防御均隨海拔和緯度升高而增大[7].作者們推測種子變大可能是為了補償由于海拔升高而產生的環境脅迫(如低溫、短光和周期等)的適應性結果,且大種子需要更多的物理防御投資抑制種子捕食;而熱帶地區石櫟種子更大則可能是熱帶環境因子較亞熱帶地區更適于生產大種子、或者熱帶是更多的種子散播媒介等原因綜合影響下的結果[41].
目前,石櫟屬的分布概況和2種主要果實類型的分布格局基本明晰,不難看出2種果實類型的果實形態分化和物化權衡并非由于其地理分布分化造成,而更可能是和與其果實相互作用的動物的長期共同進化的過程中,產生的生態位分化的結果.將在第2部分對石櫟果實和動物的相互關系做詳細的論述.除此之外,氣候因子決定了物種的適宜生境,對物種的分布格局產生直接影響[42-45].并且,有證據顯示地理因子并不能完全解釋物種特征和種間分化[46].因此,在未來分析溫度、降水等重要氣候因子對石櫟物種和2種果實類型分布的影響和規律,是進一步理解石櫟物種多樣性和果實類型分化的重要步驟.
2 石櫟的種子散播和物種更新研究
2.1 石櫟種子的捕食散播研究
石櫟種子是東亞熱帶亞熱帶常綠闊葉林內眾多動物類群的重要食物和營養來源[7-10].其中,昆蟲和脊椎動物是2類最主要的種子捕食者[32],分別代表著與石櫟種子的2種截然不同的相互關系:抑制種子散播的拮抗關系和有助于種子散播的互利共生關系[32, 47-48].昆蟲與石櫟種子的拮抗作用體現在昆蟲消費種子,獲得了能量和繁衍后代的機會,但種子產量被嚴重削弱,種子散播受限,從而對石櫟種群重建產生不利影響[48].而具有分散囤積行為的脊椎動物則和石櫟植物形成了互利共生的關系,它們同時扮演著捕食者和散播者的雙重角色:它們在消費種子獲取食物和能量的同時,也搬運種子使其遠離母樹,將種子埋藏在有利于種子萌發和幼苗建立的土壤淺表,擴大了石櫟的分布范圍和生存空間,以及繁衍后代的機率[8-9,49-50].
目前,針對石櫟果實與動物相互關系對研究十分有限,只了解到一些昆蟲和脊椎動物捕食者的片段信息:如象甲、雪片象、癭蜂可能是某些石櫟物種種子的昆蟲捕食者[51];而目前已知的石櫟的脊椎動物捕食散播者多為嚙齒類動物,如白腹鼠屬(Niviventercoxingi)、鼠屬(Rattus)、巨鼠屬(Berylmys)、姬鼠屬(Apodermus)、小泡鼠屬(Leopoldamys)、巢鼠屬(Micromys)、社鼠(Niviventerconfucianus)和針毛鼠(Niviventerfulvescens)等[9, 49-50, 52],而雖然觀察到更大型的哺乳動物如野豬也捕食石櫟果實[7],但它們是否對種子有散播還有待考證.果實的物理防御和化學成分特性和其動物捕食者密切相關[32],因此,共生且物化特性相異的2種果實類型的石櫟是種子捕食散播研究的理想模型.而且由于代表這2種果實類型的石櫟物種往往共生[7],因此我們推測具有形態和物化權衡的2種果實類型可能是針對昆蟲和脊椎動物不同捕食壓力下,進化形成的2種不同的種子保護、散播和種群更新的適應性對策:花托包被實(ER)的物理防御強,限制了昆蟲對其捕食,同時促進了脊椎動物對果實的散播,其種子大且營養豐富,有利于種子萌發和幼苗建立,但由于單個果實生殖能量投入高,限制了果實總產量和建植的種苗數量;而機械防御弱的橡實(AC)被更多類型的昆蟲捕食,也被脊椎動物更多地原地消費,但其單個果實的生殖能量投入小,果實總產量高,因此足夠數量的果實可以躲避捕食,萌發定植.因此,全面調查2種石櫟果實類型的昆蟲捕食者和脊椎動物捕食散播者,同時結合石櫟果實形態和理化特性,分析2種果實類型與昆蟲和脊椎動物的相互作用差異,是驗證2種果實類型生活史策略的假說和解析石櫟2種果實類型進化動力和其共生機制的關鍵步驟.
2.2 石櫟種子萌發特性及幼苗生長研究
在種子散播后,種子的萌發和定植是影響石櫟種群重建和分布的另一重要步驟.雖然石櫟屬植物可以大量結實,但在成年石櫟的森林中卻很難發現石櫟幼苗,這可能與石櫟種子的低萌發率相關[26, 53-56].種子的萌發率同時受到非生物因子和生物因子兩方面的影響.影響石櫟種子萌發的重要的非生物因子包括溫度、濕度和植物激素等,如研究發現石櫟種子在 25 ℃ 條件下萌發率最高[53-54];且4%的硝酸鉀、濃硫酸,赤霉素等都能促進石櫟種子的萌發[26, 54-55].生物因子如昆蟲,他們捕食石櫟種子,除了降低種子產量并限制種子散播外,昆蟲攜帶的真菌、細菌和病毒也會造成種子感染甚至死亡[32,48,56-57].種子通過各種化學和物理防御來降低被捕食和感染的風險[58-59],但這些防御也可能成為種子萌發的阻礙.例如,不同的研究都發現剝去果殼可以促進石櫟種子的萌發[26,53-54].以上研究讓我們對影響石櫟種子萌發的因素有了一些基礎的了解,但是,我們對石櫟屬2種果實類型的種子在萌發特性上的差異卻不甚了解:較大且富含營養的花托包被實(ER)種子是否比較小的橡實(AC)種子萌發率更高;或者花托包被實(ER)的木質化加厚的果殼是其萌發的阻礙,而橡實(AC)果殼脆弱因此萌發率更高?2種果實類型的種子萌發是否需要不同的適宜環境?解析2種果實類型萌發特性差異與其果實形態和物化權衡之間的聯系是我們理解石櫟物種和果實類型分化的關鍵,也是理解2種類型石櫟物種的共存和生活史策略的重要步驟.
而目前針對石櫟幼苗生長的研究,主要集中于提高苗木成活率的育苗方法:包括(1)適于石櫟幼苗生長的基質配方:如研究發現最適于東南石櫟(L.harlandii)幼苗生長的基質配方為泥炭土∶珍珠巖∶鋸屑為 3∶1∶1[60];(2)肥料對石櫟幼苗的生長和生物量表現的影響:如耳葉石櫟(L.elegans)的幼苗在一公頃 100 kg N尿素肥料劑量下生長和生物量表現最好[26];(3)通過控制蒸騰作用來提高石櫟苗木成活率:如摘除欖葉石櫟(L.oleaefolia)苗木上部分較大葉片,減少葉面蒸騰蒸發的水分可提高其苗木成活率等[61].這些研究為石櫟的造林和引種栽培奠定了重要理論指導,對石櫟的科學合理開采提供了重要的科學理論基礎.但是,我們對自然生態系統中石櫟的幼苗生長和生存卻鮮有了解.石櫟幼苗的形態和成樹形態差異明顯,這造成了野外石櫟幼苗辨認難以及不同石櫟物種幼苗鑒定困難的情況.因此,我們建議在本地尋求專業支持,如林業局或者熟悉植物的本地山民協助進行石櫟幼苗識別和物種鑒定,同時對幼苗生長發育情況進行年間追蹤觀測,深入了解石櫟群落結構和種群重建過程,分析探討石櫟多樣性和分布形成原因.
3 石櫟屬的分子系統發育及兩種果實類型的進化關系
隨著分子生物學的迅猛發展,利用基因數據進行分子系統發育研究成為目前研究的熱點領域之一.Cannon和Manos在2003年利用葉綠體基因(cpDNA)和內轉錄間隔區(nrITS)構建了少量婆羅洲熱帶及亞洲大陸石櫟物種的系統發育樹,并提出石櫟屬起源于東南亞熱帶,隨后向亞洲方向遷徙的理論[62].他們認為向亞洲遷徙的過程中,一次向西擴散到歐洲(只有化石記錄,沒有現存物種),另一次向東擴散到北美西部,有一個現存的物種[62].而后來的研究發現,北美西部的現存物種并非石櫟物種,而應認定為新屬Notholithocarpus[63-64].由于Cannon和Manos的石櫟理論還有待進一步的驗證,Yang等于2018年通過內轉錄間隔區(nrITS)和葉綠體基因的atpB-rbcL序列,構建了84個石櫟物種的3個系統發育樹(包括聯合內轉錄間隔區(nrITS)和葉綠體基因的atpB-rbcL序列構建的一個系統發育樹以及單獨由內轉錄間隔區(nrITS)或葉綠體基因的atpB-rbcL序列構建的2個系統發育樹).相對于Cannon和Manos在2003年構建的熱帶及少量亞洲大陸石櫟物種的系統發育樹,該研究囊括了1/3的石櫟物種多樣性并代表了石櫟的主要分布地區(東亞、東南亞),成為目前已發表的最完善的石櫟屬系統發育樹.另外,他們否定了Cannon和Manos的石櫟熱帶起源的假說[62],指出石櫟屬起源于中國亞熱帶,于中新世早期向東南亞遷移,隨后在大巽他群島進行原位多樣化,造成了目前東南亞石櫟物種豐富的現狀[65].
另外,Cannon和Manos在2001年通過內轉錄間隔區(nrITS)構建了21個熱帶石櫟物種系統發育樹,并和果實形態研究相結合,初步揭示了2種果實類型的系統進化關系:花托包被實(ER)至少在不同進化枝中獨立進化了2次,可能是進化中后出現的果實類型[28].他們還發現果實類型的形態變化和基因變異沒有必然的對應關系:橡實(AC)物種的種內和種間形態變異均小于其基因的變異[66-67],而花托包被實(ER)形態的變異卻大于其基因的變異[28].這說明花托包被實形態學方面臨更大的選擇壓力,同樣暗示2種果實類型可能是對應不同捕食壓力下分化的產物.但這項研究使用的基因數量(一個內轉錄間隔區(nrITS))和物種數量少(21個物種),且其果實類型劃分缺乏數據支持.
因此,對Yang等[65]發表的系統發育樹上的49個石櫟物種的果實類型進行精確劃分,并將其標注到系統發育樹上(圖3).結果顯示,49個物種中,32個物種屬于橡實(AC)類型,17個物種屬于花托包被實(ER)類型.且花托包被實(ER)的在不同的進化枝之中反復多次出現,說明其不是進化選擇的偶然性,在進化上存在一定的優勢.但這些分析還不足以完全理解2種果實類型之間的進化關系.首先,Yang等建立的系統發育樹所使用的基因數量少(只有內轉錄間隔區(nrITS)和葉綠體基因的atpB-rbcL序列),因此系統樹的穩定性較弱,種間關系支持力度不足.因此,增加基因數量來建構完善且穩定的石櫟系統發育關系,并完善石櫟物種果實形態多樣性研究十分必要.更重要的另一個方面是,目前單純的分子系統發育研究存在很多缺陷,如基因組數據集的分析技術存在偏差[68],或者單純依賴分子數據可能會誤判物種的系統發育關系等[69-70].形態學證據是完善分子系統發育和物種間關系的重要環節.因此,將石櫟物種果實形態學特征(如果實類型、果實大小、葉片大小等)與其分子發育系統結合,對其進行祖先形狀重建和系統信號發育研究可以解決分子數據難以處理的物種系統位置、提高分子進化樹的準確性[71].另一方面,通過系統發育樹追蹤果實特征的進化可以初步探究果實類型變化的分子基礎,了解石櫟果實形態多樣性與其物種進化的相互關聯,為探究石櫟屬關鍵種系統位置和理解2種果實類型進化關系提供佐證.這不僅可以幫助我們進一步驗證石櫟的起源(熱帶還是亞熱帶),也是揭示2種果實類型進化關系的關鍵.

圖3 結合分子系統發育樹的石櫟物種果實類型演化關系圖
4 總結與展望
石櫟物種果實形態多態性與其物種進化和多樣性形成密切相關.2種主要的果實類型:橡實(AC)和花托包被實(ER)代表了石櫟屬物種間豐富的果實形態變異.這2種果實類型存在果實形態和物化特性的權衡:橡實(AC)的果實和種子較小,種子內的高拒食素成分是強化學防御,包被種子的單薄果皮代表了弱物理防御;而花托包被實(ER)的種子和果實更大,種子營養成分高,木質化加厚的花托保護種子代表了強物理防御[7, 28, 35].果實發育學證據顯示這2種果實類型的形態權衡主要由花托和果皮在果實發育后期相異的分化速率造成[27].地理分布研究顯示,這2種果實類型的石櫟物種同時共享中國南部和東南亞2個分布中心,且往往在不同地區的不同森林類型中共生[7].
基于以上研究基礎,課題組推測2種果實類型在形態和物化特性上的分化如果不是地理分布差異造成,那它們很可能存在生態位的分化,代表了不同捕食壓力下長期進化出現的2種適應性對策:花托包被實(ER)物理防御強,既限制了昆蟲對其種子的捕食,也促進了脊椎動物對種子的散播,同時高營養的種子利于種子萌發和定植;而機械防御弱的橡實(AC),不僅被更多類型的昆蟲所捕食,也被脊椎動物進行大量原地消費,但其果實小,果實總產量大,最終足夠數量的果實能夠躲避捕食,完成種子萌發定植.因此,未來急需開展石櫟果實形態和理化特性與昆蟲和脊椎動物的相互作用研究,來解析石櫟2種果實類型的進化動力和其共生機制.同樣重要的是,分析對比2種果實類型的石櫟種子的萌發特性和幼苗生長情況,是回答2種果實類型的種群更新差異、生態位分化和生活史策略的另一個關鍵.
通過將果實形態研究與石櫟分子系統發育結合,初步揭示了花托包被實(ER)可能是進化中后出現的果實類型[28].而最新的石櫟系統發育樹雖然物種數量豐富[65],但是使用基因數量較少,所以對種間關系支持力度不足,且缺乏形態數據支撐.因此,完善石櫟的分子系統發育關系,并把果實形態和果實類型研究與石櫟分子系統發育相結合進行祖先狀態重建和系統發育信號分析,將為石櫟屬內關鍵種的系統位置提供果實形態佐證,明晰2種果實類型的進化關系,同時為分析果實多樣性與物種進化的相互關聯提供重要證據.
總之,石櫟果實形態和果實類型研究不僅是石櫟分類鑒定的重要證據,也對理解石櫟的種子散播和物種更新、種間聯系、進化和物種多樣性和形態多樣性關聯方面有重要作用.在此重提植物形態學的重要性,通過將石櫟果實形態研究與生物地理學、系統發育、生態學相結合,為理解石櫟物種分化與多樣性、物種進化和共存研究提供了一個全新的視角,也為其他物種的進化和多樣性研究提供了范本.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
