呂梁山瀕危植物珊瑚蘭資源調查及形態解剖學研究
2021-09-11 01:29:53楊陽曾令天景夢岳
江蘇農業科學 2021年16期
楊陽 曾令天 景夢岳


摘要:調查研究呂梁山野生珊瑚蘭生存現狀、資源分布及花芽分化的形態解剖學特征,為建立呂梁地區瀕危植物珊瑚蘭種植資源庫和保育性開發提供依據。采用野外調查、路線踏查并輔以典型樣地取樣的方法對呂梁山野生珊瑚蘭資源進行實地保護性調查;并用常規實體解剖石蠟切片法,顯微觀察珊瑚蘭花芽分化及發育的過程。結果表明,野生珊瑚蘭分布于111°13′10″~111°13′15″E,37°5′37″~37°5′31″N之間,海拔高度影響野生珊瑚蘭分布。中性土壤和 9~20 cm土壤腐殖質能夠為珊瑚蘭植株生長提供一個穩定的生長環境。珊瑚蘭的花芽形態特征與普通蘭花保持一致,即由3片外輪花被片、2片內輪花被片、1個唇瓣、1個合蕊柱構成,在合蕊柱頂端有花藥、蕊喙,底端為子房,特立中央胎座著生胚珠。研究發現,呂梁山有野生珊瑚蘭資源分布,海拔高度、土壤腐殖質顯著影響野生珊瑚蘭居群分布。
關鍵詞:呂梁山;瀕危植物;珊瑚蘭;資源調查;形態解剖;花芽分化
中圖分類號: S682.310.1 文獻標志碼: A 文章編號:1002-1302(2021)16-0122-05
野生珊瑚蘭(Corallorhiza trifida Chat.)是蘭科珊瑚蘭屬植物,多年生腐生草本,非光合蘭花,瀕危(EN)級,保護等級Ⅱ級。在我國,珊瑚蘭主要分布在西北、四川等地區[1-2]。近年來,在北京門頭溝百花山發現有珊瑚蘭生長,成為北京新記錄的植物之一[3]。珊瑚蘭屬約有13個種,多數種原產于美洲,1個種原產于亞歐大陸[4]。珊瑚蘭是腐生植物,無葉,沒有葉綠素,無法像常見的綠色植物一樣進行光合作用。它們依靠一些真菌進行共生,以此來獲取土壤中的養分[5]。珊瑚蘭對土壤要求較高,要求腐殖質豐富、沒有污染的林下環境或灌木環境[6-8]。珊瑚蘭作為一種腐生植物,生長方式較常見綠色植物大為不同,在增加植物多樣性方面有至關重要的作用[9]。珊瑚蘭在生長發育過程中,會與共生菌形成2個十分關鍵的時期,一是開花結果期,二是種子萌發及幼苗建成期,這2個時期共生菌極大地影響珊瑚蘭的生長情況。雖然其種子數量巨大,然而種子胚發育不完全,沒有胚乳,在自然條件下種子必須有特定真菌為其提供營養才能成功萌發,隨后建立穩定的共生關系[10],經查閱文獻可知,珊瑚蘭對共生菌具有極高的特異性[2]。McKendrick等研究發現,珊瑚蘭有外生菌根真菌從其他植物中獲取營養并傳輸給珊瑚蘭供其生長[11]。蔣玉玲等對珊瑚蘭菌根真菌多樣性做過研究[2],這為珊瑚蘭根的研究提供了一些研究基礎,但對于珊瑚蘭花芽的研究,鮮有報道。為研究珊瑚蘭的發育過程,須要對珊瑚蘭的花芽進行解剖研究。采用石蠟切片法[12-13],研究花芽的解剖結構和花芽發育進程,為研究珊瑚蘭提供形態解剖學依據。同時,珊瑚蘭作為瀕危植物,生存狀況極不樂觀,保護性研究刻不容緩。
此外,蘭科植物一般分布于溫暖、濕度大、通風和具散射陽光的環境,蘭科植物的生境特點決定了我國蘭科植物地理分布不均勻,主要集中在長江流域及以南地區。比較而言,由于我國北方地區蘭科植物多樣性較低,科研人員開展調查和保護工作主要集中在蘭科植物的多樣性中心即西南地區,很少關注資源相對匱乏的北方地區。但對不同地區的蘭科植物開展調查和進行多樣性保護研究,可以保護不同的基因型和變異,為蘭科植物種質基因庫的建設提供更多選擇。呂梁地區位于華北地區山西省的中部西側,境內有森林覆蓋率較高的山區,生物資源雄厚,符合蘭科植物的生境特點。但記錄該地區的野生蘭花的文獻資料較少,開展的相關工作匱乏,野生蘭花種質資源總體分布狀況不明,特別是野生珊瑚蘭在呂梁地區的分布狀況幾無報道,所以盡快對呂梁地區瀕危植物珊瑚蘭種植資源進行調查分析和保育性開發顯得尤為重要。
1 材料與方法
通過提前查閱資料確定呂梁地區交城縣龐泉溝、中陽縣上頂山和離石區吳城九鳳山等5個有代表性的,具備蘭科植物生境特點的樣地,采用野外調查、樣本采集與鑒定、整理、分類、編目相結合,選線與跑面相結合,專業踏察與當地群眾了解訪問相結合的方式,根據不同地區、山體的生態條件、植被特征及其自然環境等狀況,采取路線踏查、抽樣調查、典型樣地調查等相組合的方法,進行呂梁地區野生珊瑚蘭種質資源調查。調查工作從2016年4月上旬開始,2019年7月結束。調查內容包括居群分布情況、生長環境、生物學特征、生殖過程和種群發育情況等,并對調查結果進行分析。
采樣分析生境、物候期特征和生殖生物學特征,包括伴生植物、坡度、土壤結構、物候期(萌動、抽莖、花期、果期、枯萎)。野外用甲醛-乙酸-乙醇(FAA)固定液采樣,用4 ℃ 冰盒取回,通過固定—沖洗—脫水—透明—浸蠟—包埋—切片—粘片—烘片—染色封片,顯微鏡觀察珊瑚蘭的花芽分化及形態特征,其中切片厚度為7.5 μm。
2 結果與分析
2.1 野生珊瑚蘭居群分布情況分析
2016年4月至2019年7月經過多次踏查,擬踏察的龐泉溝自然保護區的八水溝、八道溝、福祿溝,離石九鳳山、白馬仙洞景區等5個調查地均未發現野生珊瑚蘭種群。在地理位置為111°13′9″E、37°5′37″N的中陽縣上頂山發現有野生珊瑚蘭種群分布。
2017年4月開始對中陽縣上頂山野生珊瑚蘭居群分布情況進行詳細調查。按照不同的海拔高度和經緯度,選取代表性區域作為典型樣地,每個樣地10 m2,重復3次記錄數據。隨海拔高度變化踏查,開始發現野生珊瑚蘭群落的海拔高度為 1 820 m,經度為111°13′10″E,緯度為37°5′37″N。
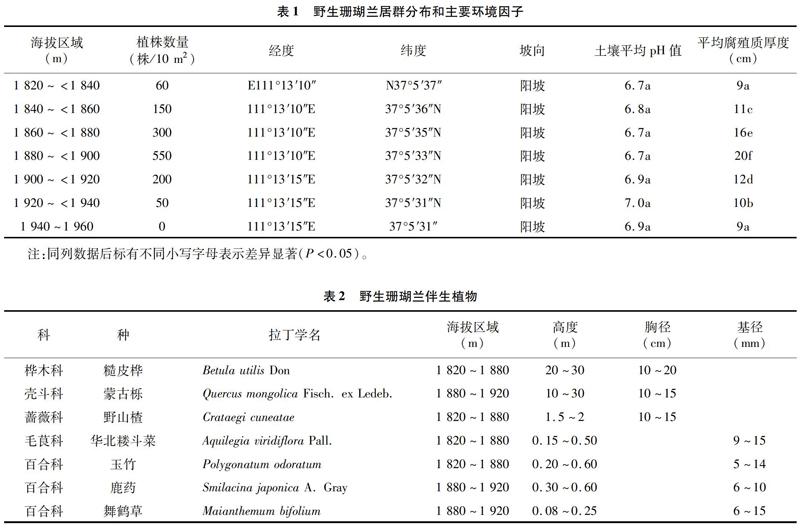
由表1可知,上頂山野生珊瑚蘭群落分布于111°13′10″~111°13′15″E,37°5′31″~37°5′37″N,海拔高度為1 820~1 940 m,坡向為陽坡。樣地野生珊瑚蘭植株根際附近土壤平均pH值為6.7~7.0,平均腐殖質厚度為9~20 cm。土壤平均腐殖質厚度由9 cm升高至20 cm時,植株數量由平均 60株/10 m2 升高至550株/10 m2;當土壤平均腐殖質厚度降低至9 cm時,植株數量下降至0株/10 m2。可以看出,海拔高度、土壤平均腐殖質明顯影響野生珊瑚蘭居群分布。
2.2 野生珊瑚蘭伴生植物分析
伴生植物與野生珊瑚蘭分布有著緊密聯系,樣地調查中,隨著海拔高度的提升,野生珊瑚蘭伴生植物也發生了變化(表2)。海拔為1 400~1 800 m的區域,多為華北落葉松、白皮松、油松、白樺、野山楂、山荊子、杜梨、暴馬丁香、薔薇、車前、舞鶴草、玉竹、華北烏頭、藜蘆、瓣蕊唐松草、芍藥等。海拔為 1 820~<1 940 m的區域,喬灌木層可分為10~30 m 的大中型喬木,主要是糙皮樺、蒙古櫟;還有1.0~3.0 m的灌木,主要是野山楂、薔薇;草本層植物分布比較密集,主要包含了玉竹、鹿藥、華北耬斗菜、舞鶴草等。海拔為1 940~1 960 m的區域,喬木層植物主要為山楊樹、糙皮樺,蒙古櫟幾乎沒有,草本層植物有高山野罌粟、東方草莓、水楊梅等。
2.3野生珊瑚蘭生物學特征
野生珊瑚蘭為總狀花序,植株平均高度為10.43~13.41 cm,花葶高度為10~13 cm,每株小花數量平均4~7朵。海拔高度影響株高和小花數量。海拔為1 880~< 1900、1 900~<1 920 m的區域,野生珊瑚蘭平均株高分別為12.07、13.41 cm,明顯高于其他海拔的植株;平均小花數量為7朵,也明顯高于其他海拔的植株(表3)。
野生珊瑚蘭植株根狀莖,直徑為1.5~3.0 mm,呈珊瑚狀,白色,多分枝[14]。海拔為1 860~<1 900 m的區域,野生珊瑚蘭平均根狀莖直徑為3.0 mm,明顯高于其他海拔的植株(圖1)。
2.4 野生珊瑚蘭花芽分化及解剖結構分析
野生珊瑚蘭成年以后,在呂梁山特殊環境因素的影響下完成成花誘導,形成花器官原基,由營養生長期進入生殖生長期[15]。本試驗研究其花芽分化及解剖結構,為蘭科植物種質資源保護工作奠定基礎。
試驗發現,珊瑚蘭的花器官形態符合典型蘭花形態標準,由內到外依次為合蕊柱、內輪花被片、外輪花被片,內外輪花被片均為3片,其中一片內輪花被片特化為唇瓣,這也是蝴蝶蘭[16]、寒蘭、文心蘭[17]等蘭花的特有現象。珊瑚蘭的花蕾呈梭形,外由花萼包被,長3~6 mm,寬1.0~1.5 mm。花被下方有綠色梭形鼓囊,外有長條形棱,是珊瑚蘭花的子房,長2~6 mm,直徑為1.5~2.5 mm。花被片長度與子房長度基本符合1 ∶ 1的長度比例。
珊瑚蘭花芽分化開始于珊瑚狀根狀莖生長點,花序原基最先分化,生長點突起,花原基伸長,細胞質濃,細胞小(圖2-1)。花序原基繼續分化,進入頂花芽花被原基分化期。頂花芽在雄蕊原基分化期,分化出雄蕊原基,整體明顯向外突出,由內到外依次為雄蕊、花被、苞片(圖2-2、圖2-3)。還分化出雌蕊原基,由內到外依次為雌蕊、花柱、雄蕊、2層花被、花苞(圖2-4、圖2-5)。
珊瑚蘭花芽由外到內,依次是花被片、唇瓣、合蕊柱;其中花被片包括萼片和花瓣,蕊柱頂端著生1
個花藥,1個花藥里有2個花粉塊,觀察顯微圖(圖2-5)可知,花粉塊密度明顯大于周圍組織,花粉塊下部有蕊喙、花柱、子房。花被片,包覆在花芽的最外側,外輪花被片日后發育成為3片萼片,內輪花被片則發育成為2片花瓣。
唇瓣位于合蕊柱之上,花被片之下,呈芭蕉葉狀扣在花藥頂部。附著點在合蕊柱側面,由2~3層細胞構成,細胞排列緊密均勻。
合蕊柱位于花芽正中,周圍由花被片包覆,頂端是有2個花粉塊的花藥,往下依次是蕊喙、花柱、子房。其中,花粉塊的細胞密度明顯大于周圍組織,十分容易分辨。其次花粉塊下方有1個略向上伸的突起,此處為蕊喙,別稱小喙,是蘭科植物的蕊柱前面舌狀突起的部分。在柱頭的3個裂片中,2個形成柱頭,不結實性的1個即變成這種蕊喙(圖2-5)。
子房在整個花結構處于靠下部分,中空,子房內壁上附著很多圓卵形小體,稱為胚珠,受精后發育成種子,胚珠附著在子房中的胎座上。
胚珠是一種受精后發育成種子的結構,被稱為種子的前體。子房內,胚珠包被,通過珠柄著生于子房內壁的胎座上(圖2-6、圖2-7)。
3 結論與討論
呂梁山野生珊瑚蘭資源分布于111°13′10″~111°13′15″E,37°5′31″~37°5′37″N;4月上旬開始萌動,4月下旬至5月下旬為開花期,6月中下旬種子成熟;中性土壤和9~20 cm土壤腐殖質能夠為珊瑚蘭植株生長提供一個穩定的生長環境;海拔高度、土壤平均腐殖質明顯影響野生珊瑚蘭居群分布。海拔為1 820~1 940 m的區域,野生珊瑚蘭主要的伴生植物喬木層為10~30 m的大中型喬木,主要是糙皮樺、蒙古櫟;1.0~3.0 m的灌木,主要是野山楂、薔薇;草本層植物主要半生植物為舞鶴草、鮮有玉竹、鹿藥、華北耬斗菜。
通過觀察珊瑚蘭的花芽發現,珊瑚蘭的花芽形態特征與普通蘭花保持一致,即由3片外輪花被片、2片內輪花被片、1個唇瓣、1個合蕊柱構成,在合蕊柱頂端有花藥、蕊喙,底端為子房,特立中央胎座著生胚珠。常規實體解剖石蠟切片法觀察珊瑚蘭花芽分化依次為花序原基分化期、花被原基分化期、雄蕊原基分化期、雌蕊原基分化期、胚珠形成期。
關于呂梁山瀕危植物珊瑚蘭資源調查的工作由2016年4月開始,歷經3年,在前期充分查閱資料制定了交城縣龐泉溝、中陽縣上頂山和離石區吳城九鳳山等5個有代表性的,具備蘭科植物的生境特點的樣地,但是在踏查的過程中文獻記錄龐泉溝自然保護區八水溝、八道溝、福祿溝有野生珊瑚蘭居群分布,但在調查過程中,通過3次調查均未發現有珊瑚蘭居群分布,分析原因可能因為龐泉溝自然保護區旅游和礦業開發破壞了珊瑚蘭生境,具體原因還需要進一步調查才能確定。中陽縣上頂山屬呂梁山關帝山脈,自然生態保護較好,但是在發現野生珊瑚蘭居群分布后,筆者所在課題組又擴大范圍對該地區野生珊瑚蘭資源進行了調查,但是遺憾的是除了本研究范圍,其他地方均未發現珊瑚蘭。珊瑚蘭自身無法進行光合作用,只能由其共生的菌類供給營養。此前有學者研究,珊瑚蘭的共生真菌有較強的專一性,要滿足珊瑚蘭及其共生菌能夠同時良好地生長,對環境的要求必然是較為嚴格的。目前關于珊瑚蘭的研究很少,野生珊瑚蘭外形奇特,花期較長,有很大的開發價值,現今野生種群數量呈明顯下降趨勢。由于種群數量較少,繁殖生境要求嚴格,直接導致種群擴散緩慢,保護和恢復該植物種群的結構和數量顯得格外重要[18-20]。
參考文獻:
[1]侯天文. 四川黃龍溝優勢蘭科植物菌根真菌多樣性研究[D]. 北京:北京林業大學,2010.
[2]蔣玉玲. 遼寧省內九種蘭科植物菌根真菌多樣性研究[D]. 沈陽:沈陽農業大學,2018.
[3]沐先運,劉 曉,林秦文,等. 北京百花山自然保護區蘭科植物多樣性及保護評價[J]. 北京林業大學學報,2010,32(增刊1):14-17.
[4]石昌魁. 甘肅省蘭科植物系統分類與區系地理[D]. 蘭州:甘肅農業大學,2008.
[5]侯天文. 四川黃龍溝優勢蘭科植物菌根真菌多樣性研究[D]. 北京:北京林業大學,2010.
[6]冉景丞,魯成巍. 茂蘭自然保護區蘭科植物資源現狀及保護利用途徑[J]. 安徽農業科學,2009,37(11):5209-5211.
[7]孫 悅,李 標,郭順星. 腐生型蘭科植物研究進展[J]. 廣西植物,2017,37(2):191-203.
[8]董建新,朱毓永,李鐵軍,等. 小五臺山野生蘭科植物研究[J]. 承德民族師專學報,2009,29(2):54-55.
[9]孫曉穎. 五種野生兜蘭植物菌根真菌多樣性研究[D]. 北京:北京林業大學,2014.
[10]孫曉穎,張武凡,劉紅霞. 帶葉兜蘭種子原地共生萌發及有效菌根真菌的分離與鑒定[J]. 熱帶亞熱帶植物學報,2015,23(1):59-64.
[11]McKendrick S L,Leake J R,Taylor D L,et al. Symbiotic germination and development of myco-heterotrophic plants in nature:ontogeny of Corallorhiza trifida and characterization of its mycorrhizal fungi[J]. The New Phytologist,2000,145(3):523-537.
[12]劉桂芝. 石蠟切片標本的制作過程[J]. 通化師范學院學報,2004(11):90-91.
[13]李淑嫻. 墨蘭成花機理及花期調控技術研究[D]. 福州:福建農林大學,2016.
[14]王長寶,徐增奇,王 鶴,等. 瀕危植物珊瑚蘭(Cypripedium macranthos SW.)地下器官的形態解剖學研究[J]. 東北師大學報(自然科學版),2015,47(3):111-115.
[15]龔 湉. 寒蘭成花機理及花期調控研究[D]. 福州:福建農林大學,2015.
[16]韋 莉. 蝴蝶蘭花芽分化的解剖學觀察及成花機理研究[D]. 南京:南京林業大學,2010.
[17]彭 芳. 文心蘭花芽形態分化及其生理生化的研究[D]. 南寧:廣西大學,2011.
[18]董林水. 晉西基巖山地與黃土丘陵區生態過渡帶植被特征及生態恢復機理研究[D]. 北京:中國林業科學研究院,2005.
[19]李忠超,王武源. 瀕危藥用植物八角蓮生態生物學特征[J]. 熱帶亞熱帶植物學報,2006,14(3):190-195.
[20]張 毓,張啟翔,趙世偉,等. 瀕危植物大花杓蘭胚與珠被發育的研究[J]. 園藝學報,2010,37(1):72-76.
