氮添加對不同植被類型中毛竹與常綠闊葉林木生長的影響
2021-09-12 05:51:49賴雁南鐘全林李寶銀程棟梁余華徐朝斌薛翔海鄭文婷
防護林科技 2021年2期
賴雁南 鐘全林 李寶銀 程棟梁 余華 徐朝斌 薛翔海 鄭文婷



摘 要 氮沉降背景下毛竹向常綠闊葉林擴張問題是當前我國亞熱帶區域面臨的主要生態問題與研究熱點之一。以毛竹、竹闊混交林及常綠闊葉林為對象,通過1 a的氮添加試驗,分析毛竹與常綠闊葉林木在不同植被類型中生長的差異,以探究氮沉降對不同植被類型中毛竹與常綠闊葉林木生長的影響。結果顯示:(1)竹闊混交林中毛竹的平均胸徑增長量、平均單株生物量增長量及生物量相對生長速率高于毛竹林中的毛竹,但竹闊混交林中闊葉林木的平均胸徑增長量、平均單株生物量增長量及生物量相對生長速率則顯著低于常綠闊葉林中的闊葉林木(P<0.05);(2)低氮添加會減小毛竹在竹闊混交林與毛竹林中的個體差異,但會增大常綠闊葉林木在竹闊混交林與常綠闊葉林之間的生長差距,低氮添加有利于毛竹向常綠闊葉林擴張;(3)高氮添加會顯著抑制毛竹的生長,一定程度上可減緩毛竹向常綠闊葉林擴張的速度。
關鍵詞 氮添加;生物量;相對生長速率;毛竹;常綠闊葉林木
中圖分類號:S714.8;S718.54 ? ? 文獻標識碼:A ? doi:10.13601/j.issn.1005-5215.2021.02.001
Abstract The expansion of Phyllostachys pubescens to evergreen broadleaved forests under the background of nitrogen deposition is one of the main ecological problems and research hotspots facing China's subtropical regions. Taking Phyllostachys edulis,Phyllostachys edulisevergreen broadleaved mixed forest and evergreen broadleaved forest as the object,the growth difference between Phyllostachys edulis & evergreen broadleaved forest in different vegetation types were analyzed through 1year nitrogen addition test,and explores the influence of Phyllostachys edulis expansion into evergreen broadleaved forest under the background of nitrogen deposition on the growth of Phyllostachys edulis and evergreen broadleaved forest. Results showed that: (1) In Phyllostachys edulisevergreen broadleaved mixed forest average growth in DBH,average individual biomass growth and biomass per plant relative growth rate is greater than Phyllostachys edulis in Phyllostachys edulis forest,Phyllostachys edulisevergreen broadleaved mixed forest,evergreen broadleaved forest significant differences mainly growth parameters,Phyllostachys edulis expansion leads to the average individual DBH growth rates of evergreen broadleaved forest biomass growth and biomass per plant and average relative growth rate dropped significantly;(2) Low nitrogen addition can reduce the growth difference of Phyllostachys edulis in Phyllostachys edulisevergreen broadleaved mixed forest and Phyllostachys edulis forest,but increase the growth difference of evergreen broadleaved forest in Phyllostachys edulisevergreen broadleaved mixed forest and evergreen broadleaved forest. Low nitrogen addition is beneficial to the expansion of Phyllostachys edulis into evergreen broadleaved forest;(3) The addition of high nitrogen can significantly inhibit the growth of Phyllostachys edulis and alleviate the expansion of Phyllostachys edulis to evergreen broadleaved forest.
Key words nitrogen addition;biomass;relative growth rate;Phyllostachys edulis;evergreen broad leaved trees
氮(N)是陸地生態系統凈初級生產力最主要的限制元素之一[1,2]。工業革命以來,人類生產生活方式的改變使得全球大氣活性氮化合物含量上升 [3],導致大氣氮沉降格局的改變。當前世界氮沉降分布中心正由歐美等老工業基地轉移至各發展中國家,出現由溫帶逐漸擴散至熱帶亞熱帶的趨勢[4]。隨著經濟快速發展,我國部分地區已出現高氮沉降問題,特別是在南方亞熱帶森林區域,氮沉降量已接近歐洲最高氮沉降水平[5-7]。預計未來幾十年我國氮沉降量仍將持續增加,日益嚴峻的氮沉降問題已成為我國生態建設中被關注的熱點[8]。
氮沉降對植物及森林生態系統的影響受其沉降量的不同而表現不同 [9],適度的氮沉降能增加土壤可利用氮含量,促進植物生長與生物量的積累,提高生態系統的生產力,但過量的氮沉降則會抑制植物的生長,降低生態系統的生物多樣性[10]。氮沉降對植物生長的影響還取決于樹種差異,不同樹種對氮沉降的響應機制不同,導致其對氮的利用和配置存在差異[11]。有研究發現,氮添加能夠提高土壤中活性N含量,從而促進植物生長,提高草原生態系統總生物量 [12];氮添加量與植物地上生物量呈顯著正相關 [13],但也有人研究發現,氮添加量與生物量呈負相關關系,較低的氮添加可使得中國東北落葉闊葉林的生物量達到最大值[14]。林木對氮添加的響應程度,不僅受到氮添加量的影響,還與研究區域、樹種及施氮量、氮肥種類、施氮年限等因素有關[15-18]。目前,國內外在氮添加對森林生態系統方面的研究主要集中于同一生態系統中森林的生長狀況、土壤理化性質、土壤微生物及植物凋落物分解等方面[19],而有關氮添加對植物在不同森林類型中的生物量及其相對生長速率差異的影響研究則相對較少。因此有必要開展這方面研究,以探究毛竹及常綠闊葉樹種在不同林分類型內對氮添加的響應特征,為進一步開展氮沉降背景下常綠闊葉林可持續經營技術及揭示氮沉降背景下毛竹向常綠闊葉林內的擴張機制等提供理論依據。
常綠闊葉林為我國亞熱帶地區的典型地帶性植被,其物種多樣性豐富,在保護區域生態環境、平衡全球性碳水循環及維持人類社會可持續發展等方面發揮著重要作用[20-22]。毛竹(Phyllostachys edulis)是禾本科剛竹屬的一種散生喬木狀竹類植物。常綠闊葉林周邊的毛竹依靠地下根莖(即竹鞭)向常綠闊葉林區蔓延而實現種群擴張,致使闊葉純林演替為竹闊混交林,甚至在人為干預下逐漸形成毛竹林[23]。毛竹擴張導致森林生態系統的功能與景觀質量退化問題正逐漸為林學及生態學等學科相關學者所關注。已有研究發現,毛竹擴張引發了諸多生態環境問題,但人們往往因其具有較高的經濟效益而忽視其造成的負面影響[24,25]。因此,研究氮添加對毛竹與常綠闊葉樹種生長的影響,不僅有助于揭示毛竹與常綠闊葉樹種對大氣氮沉降的響應機制,對開展常綠闊葉林保護技術研究等也具有重要現實指導意義。
1 研究區概況與試驗方法
1.1 研究區概況
試驗地位于江西陽際峰自然保護區,地處江西省東北部,武夷山北麓(27°51′10″—28°02′20″ N, 117°11′30″—117°28′40″ E)。保護區森林總面積10 946 hm2,其中活立木蓄積量622 802 m3,立竹量9.8萬株,森林覆蓋率高達99.7%,天然常綠闊葉植被類型分布面積占總面積的68%,為我國亞熱帶常綠闊葉林的典型代表,同時也是毛竹適生區[26]。該地屬中亞熱帶濕潤季風氣候區,年均溫11.4~18.5 ℃,年均降水量1 870~2 191.3 mm,年均無霜期200~256 d,四季分明,溫暖濕潤,水熱條件良好,有利于動植物生長發育,為試驗研究提供了理想的平臺。
1.2 模擬氮沉降試驗設計
于2017年3—4月分別在天然毛竹林、天然常綠闊葉林及其中間過渡地帶的竹闊混交林內各設置9個樣地,共27個樣地,其中毛竹林樣地規格為10 m×10 m,常綠闊葉林與竹闊混交林樣地規格均為15 m×15 m;3種植被類型均為經自然更新達20 a以上的天然林,所選設的樣地海拔約為730 m;同一森林類型樣地海拔相近,毛竹林海拔相對低于常綠闊葉林。分別植被類型在所選設的9個樣地中隨機設置對照(CK,0 kg?a-1)3種處理,每處理設置3個重復。所添加的氮肥為尿素,各處理的氮添加量中均未包含自然氮沉降量。將所確定的年氮添加量均分成各月添加量,并換算成各樣地面積的月氮添加量。于2017年5月—2018年8月,于每月月底晴朗無風的9:00—11:00進行,持續施氮16個月。每次噴施之前,將稱取的氮肥溶解于蒸餾水中(毛竹林樣方是將氮肥溶解于10 L水中,竹闊混交林與天然常綠闊葉林樣方是將氮肥溶解于20 L水中),使用雙噴頭噴霧器將其水溶液人工來回均勻噴灑在林地上,其中,對不施肥的對照組則噴灑等量的蒸餾水,以消除誤差。3種植被類型樣地基本信息見表1。
1.3 指標選取及測定
分別于2017年4月第1次施肥之前及2018年8月對樣地內毛竹和胸徑≥5.0 cm的林木進行調查,測定胸徑、株高(常綠闊葉樹種株高為樹冠最高處離地面的垂直距離,毛竹的為枝下高)及樹齡,用于計算各樣地植物的生物量。
1.3.1 常綠闊葉林木與毛竹平均胸徑與胸徑增長量的計算
常綠闊葉林木與毛竹平均胸徑(DBH)計算公式如下:
DBH=1N∑ni=1(DBHi)2 (1)
式中:DBH為常綠闊葉林木(或毛竹)的平均胸徑(cm);n為樣地內胸徑≥5.0 cm常綠闊葉林木(或所有毛竹)的數量;DBHi為樣地內第i株常綠闊葉林木(或毛竹)的胸徑。
常綠闊葉林木與毛竹胸徑增長量計算公式如下:
ΔDBH= DBH2- DBH1(2)
式中:ΔDBH為常綠闊葉林木(或毛竹)的平均胸徑增長量(cm);DBH1與DBH2分別為期初與期末常綠闊葉林木(或毛竹)的平均胸徑(cm)。
1.3.2 常綠闊葉林木與毛竹生物量的計算
(1)常綠闊葉林木生物量的計算采用材積源生物量法[27]和已有文獻所建立的回歸方程[28]計算不同植被類型常綠闊葉林木的生物量。回歸方程如下:
1.3.3 生物量增長量與相對生長速率的計算
1.4 數據處理
利用Microsoft Excel 2010軟件對數據預處理。運用SPSS 20.0軟件對氮添加和植被類型影響下單株毛竹及闊葉林木生長指標進行雙因素方差分析;對毛竹和林木在同一植被類型不同氮添加水平下及同一氮添加水平下同一樹種在不同植被類型中的生長指標進行單因素方差分析,用最小顯著差異法(LSD) 進行處理間差異顯著性比較(α=0.05),使用Origin2018軟件進行繪圖。
2 結果與分析
2.1 植被類型與氮添加對毛竹與常綠闊葉林木生長的影響
雙因素方差分析結果(表2)顯示,生長在毛竹純林與竹闊混交林中的毛竹僅其平均單株生物量存在顯著差異(P<0.05),但生長在常綠闊葉林與竹闊混交林中的常綠闊葉林木的平均高、平均單株胸徑、平均單株生物量、平均單株生物量相對生長速率、單位面積生物量相對生長速率5個指標均有顯著影響(P<0.05);氮添加對毛竹各生長指標均無顯著影響,但對林木的生物量及生物量相對生長速率則有極顯著影響(P<0.01)。植被類型和氮的交互作用對毛竹沒有顯著影響,但對闊葉林木的平均單株生物量和平均單株生物量相對生長速率有極顯著影響(P<0.01)。
2.2 毛竹與常綠闊葉林木在不同植被類型中的生長差異
通過對未添加氮的毛竹林、常綠闊葉林及竹闊混交林數據進行分析,結果發現,生長在竹闊混交林內的毛竹,其平均單株生物量增長量及生物量相對生長速率雖高于其在毛竹林內的生長量,但兩者差異不明顯(P>0.05,圖1A),表明生長在竹闊混交林中的毛竹個體生物增長量雖大于毛竹純林中的個體生物增長量,但兩者差異不明顯;生長在竹闊混交林內的常綠闊葉林木,其平均單株生物增長量及平均單株生物量相對生長速率明顯低于其生長在常綠闊葉林內(P<0.05,圖1B),說明與竹闊混交林相比,常綠闊葉林木更適合在常綠闊葉林環境中生長。
2.3 氮添加對毛竹林及竹闊混交林中毛竹生長的影響
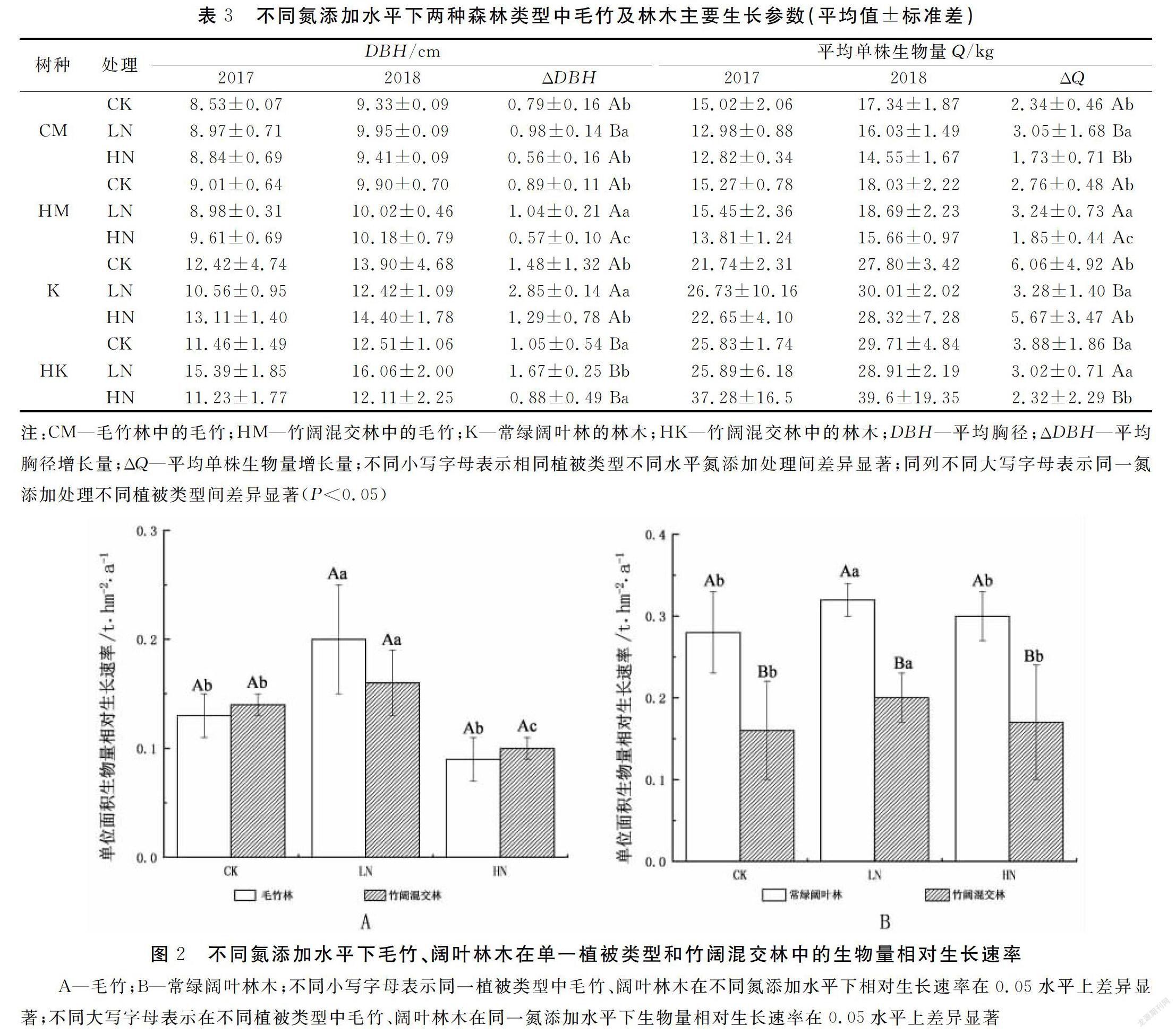
由表3和圖2A可以看出,毛竹林中的毛竹在添加低氮后,平均單株胸徑增長量、平均單株生物量增長量及生物量相對生長速率均有顯著增加(P<0.05),但高氮添加則會造成毛竹林中的毛竹平均單株胸徑增長量、平均單株生物量增長量及生物量相對生長速率均有減少,且與對照組差異不明顯(P>0.05)。
在竹闊混交林中,氮添加會顯著影響竹闊混交林中毛竹的平均單株胸徑增長量、平均單株生物量增長量及生物量相對生長速率。低水平的氮添加,會顯著增大竹闊混交林中毛竹的平均單株胸徑增長量、平均單株生物量增長量、生物量相對生長速率(P<0.05);高水平的氮添加,則會顯著減少竹闊混交林中毛竹的平均單株胸徑增長量、平均單株生物量增長量、生物量相對生長速率(P<0.05)(表3)。
另從表3與圖2A中可看出,氮添加會在一定程度促進竹闊混交林中的毛竹生長,使混交林中的毛竹胸徑生長量、平均單株生物生長量及單位面積生物量生長速率高于毛竹純林,但差異不明顯,說明氮添加背景下有利于毛竹向竹闊混交林擴張。
2.4 氮添加對常綠闊葉林及竹闊混交林中林木生長的影響
由表3和圖2B可以看出,在常綠闊葉林中的闊葉林木在添加低氮后,平均單株胸徑增長量、平均單株生物量增長量、生物量相對生長速率均有顯著增加(P<0.05)。高氮添加后,闊葉林木的平均單株胸徑增長量、平均單株生物量增長量及生物量相對生長速率無顯著變化。
在竹闊混交林中,不同水平氮添加對混交林中的闊葉林木平均單株胸徑增長量、平均單株生物量增長量、生物量相對生長速率影響程度不同。低水平氮添加,會顯著增大竹闊混交林中闊葉林木的平均胸徑增長量、平均單株生物量增長量、生物量相對生長速率(P<0.05);但高氮添加則對竹闊混交林中闊葉林木的平均胸徑增長量、平均單株生物量增長量、生物量相對生長速率影響不明顯。
上述結果表明,不施氮時毛竹擴張會顯著抑制林木的生長;低氮添加會顯著促進常綠闊葉林中的闊葉林木生長,會增大闊葉林木在常綠闊葉林和竹闊混交林中生長的差異,有利于毛竹向闊葉林擴張;高氮添加對竹闊混交林中的毛竹和闊葉林都有抑制作用,毛竹受抑制程度強于闊葉林,因此高氮一定程度上緩解了毛竹向闊葉林擴張的速率。
3 討論
3.1 毛竹與常綠闊葉林木在不同植被類型中的生長差異
常綠闊葉植被類型分布區同時也是毛竹適生區,毛竹擴張引發森林景觀變化、闊葉樹種生長減緩及碳儲量的改變[30]等生態變化,這些變化可表現為植物部分形態特征和生長指標的改變。本研究中混交林中的毛竹平均胸徑增長量、枝下高、平均單株生物量增長量相比毛竹林均有增加但不顯著,毛竹單位面積生物量在混交林和毛竹林存在顯著差異。這一結論符合毛竹的生長特性,春筍破土前竹鞭的發育、土壤狀況、水肥等條件決定了春筍的大小,春筍后期不會增大決定了新竹胸徑的大小[31,32]。
種間競爭是塑造植物形態和生活史的主要動力之一,Grime理論認為,相對生長速率(RGR)是衡量植物競爭的關鍵指標之一[33,34]。本文通過生物量RGR的分析發現,受毛竹擴張影響形成的竹闊混交林中常綠闊葉林木RGR有顯著下降,并且竹闊混交林中闊葉林木的平均胸徑增長量、單株生物量增長量、單位面積生物量相比常綠闊葉林也均有顯著下降,這與趙雨虹的研究結果[35]相符,表明隨毛竹擴張程度加劇,常綠闊葉樹胸徑、生長速率逐漸降低。這很大程度可能緣于毛竹具有生長快速、繁殖力強、形態多樣、集團協同等內稟優勢[36]。毛竹這些生長特點有利于毛竹在常綠闊葉林中的擴張,毛竹擴張后在竹闊混交林中相對生長速率提高,而毛竹和闊葉林木的種間競爭導致常綠闊葉林木的相對生長速率下降,各項生長指標增長量減少。
毛竹對常綠闊葉林的擴張影響了常綠闊葉林的系統結構和功能。森林植被生物量是研究森林生態系統結構與功能的重要指標,生物量估算已經成為研究森林結構和功能的重要內容。本研究通過計算毛竹及常綠闊葉林木在竹闊混交林與毛竹林、常綠闊葉林中平均單株生物量增長量的差異,發現毛竹在竹闊混交林(2.76 kg -1)中平均單株生物量增長量(ΔQ)與在毛竹林(2.34 kg -1)相比提高17.9%,而常綠闊葉林木在竹闊混交林中單位面積生物量增長量(3.88 kg -1)與在常綠闊葉林(6.06 kg -1)相比下降超過36.0%。這是因為當植物入侵原有森林生態系統時,會對其碳循環產生影響,進而導致植被生物量變化[37],如楊清培等[38]在研究毛竹入侵常綠闊葉林生態系統時發現植被碳儲量出現顯著降低,森林的總碳儲量減少了15.37%, 其中植被碳儲量減少了12.01%, 土壤碳儲量減少了17.30%。毛竹擴張使得在竹闊混交林中毛竹生長加快,常綠闊葉林木生長減慢,從而減少常綠闊葉林物種多樣性及森林植被碳儲量,影響了常綠闊葉林生態系統的穩定性。
3.2 不同氮添加對毛竹及常綠闊葉林木生長的影響
氮沉降會改變森林的物種生物量的增長速率,其變化的程度取決于森林種類、林木徑級與氮沉降濃度等因素的綜合影響[38]。本研究中低濃度氮添加促進毛竹生長,高濃度氮添加抑制毛竹生長,表現為毛竹的平均單株生物量增長量、生物量相對生長速率隨氮添加量增加(50、100 kg ?hm-2 -1)出現先升高后下降的變化,這和杜瑩等[39]研究發現隨添加的氮增加(30、60、90 kg -2 -1)表現為先升高后下降結論相符。這可能是因為不同水平的氮添加對毛竹光合作用的影響不同,進而影響了毛竹生物量的變化。劉寧的研究結果也支持了這一猜想,氮沉降促進了毛竹的光合速率,但是長期高水平的氮沉降會對毛竹的光系統Ⅱ產生一定傷害[40]。氮添加縮小了毛竹在毛竹林和竹闊混交林中生長的差異。這可能和單位面積內物種間競爭及環境最大承載量有關,毛竹林的毛竹在單一植被類型中生長不存在種間競爭,限制樣地中毛竹最大生長數量的主要是環境最大承載量而不是氮素含量的高低。竹闊混交林中,毛竹與闊葉林競爭生長,毛竹對氮素的更高效利用有利于毛竹侵占闊葉樹種更大的生長空間,最終表現出氮添加縮小了毛竹在毛竹林和竹闊混交林中的生長差異,都趨向于樣地毛竹的最大環境承載量。
中國東北的落葉闊葉林的大量研究得出較低的氮添加(25 kg hm-2 a-1)導致最大的凈生產力,隨著氮添加的增加(50 kg ?hm-2 a-1)正面效應降低,氮添加量最大時(75 kg -2 -1)正面效應消失[41]。本研究中低氮添加會顯著增加常綠闊葉林木的平均胸徑增長量、平均單株生物量增長量及生物量相對生長速率,高氮添加對闊葉林木的生長無顯著抑制作用。這可能和試驗林下噴施氮肥的施肥方式有關。尿素適宜深施覆土,尿素在轉化前是分子態的,不能被土壤吸附,容易隨水流失,轉化后的氨也易揮發。而高氮仍對竹闊混交林中的毛竹表現出顯著抑制作用,這可能和毛竹的氮飽和值低于闊葉林木有關,陶晨悅等的研究支持這一猜想,干旱脅迫條件下,隨著氮沉降水平的增加,毛竹生物量逐漸增大,但當氮沉降量大于60 kg ?hm-2 ?a-1時,生物量不再增大甚至有減小的趨勢[42]。這表明氮素添加對毛竹的正面效應在氮濃度超過60 kg ?hm-2 a-1時消失,闊葉林木的氮飽和值要高于毛竹。低氮添加會加大闊葉林木在常綠闊葉林和竹闊混交林中生長的差異。這也是在竹闊混交林中物種間競爭能力低于毛竹的表現,短期的低氮添加有利于毛竹向常綠闊葉林的擴張。高水平的氮添加會顯著抑制竹闊混交林中毛竹生長,對闊葉林木抑制作用不顯著,表明短期的高氮添加一定程度上能緩解毛竹在常綠闊葉林中的擴張趨勢。
上述分析結果僅是16個月的氮添加處理試驗結果,隨著持續的氮沉降,可能有些結果會發生變化。另外,毛竹擴張的地下根系入侵是擴張過程中非常關鍵和重要的部分,本研究受限于試驗條件,沒有探討毛竹和林木地下部分對毛竹擴張和氮添加的響應。今后還需對其作進一步的深入研究,有利于進一步揭示毛竹在氮沉降背景下對闊葉林入侵的機制。
4 結論
毛竹擴張后,竹闊混交林中毛竹的平均單株生物量增長量及生物量相對生長速率均大于毛竹林,竹闊混交林中的林木平均單株胸徑增長量、平均單株生物量增長量及相對生長速率相比常綠闊葉林有顯著下降,表明毛竹擴張能夠在一定程度上促進毛竹生長,顯著抑制闊葉林木的生長;低氮添加會加大闊葉林木在竹闊混交林和常綠闊葉林的生長差異,減小毛竹在竹闊混交林和毛竹林的生長差異,有利于毛竹向常綠闊葉林的擴張;高氮添加會顯著抑制毛竹生長,對林木生長無顯著抑制作用,一定程度上可緩解毛竹向常綠闊葉林擴張。
參考文獻:
[1] 周曉兵,張元明.干旱半干旱區氮沉降生態效應研究進展[J].生態學報,2009,29(7):3835-3845
[2] ?David S,Kathleen K.Nitrogen Limitation of Net Primary Productivity in Terrestrial Ecosystems Is Globally Distributed[J].Ecological Society of America,2008,89(2):371-379
[3] ?Galloway J,Dentener F,Capone D, et al.Nitrogen Cycles: Past,Present,and Future[J].Kluwer Academic Publishers,2004,70(2):153-226
[4] 魯顯楷,莫江明,張煒,等.模擬大氣氮沉降對中國森林生態系統影響的研究進展[J].熱帶亞熱帶植物學報,2019,27(5):500-522
[5] 王小治,朱建國,高人,等.太湖地區氮素濕沉降動態及生態學意義:以常熟生態站為例[J].應用生態學報,2004(9):1616-1620
[6] 孫本華,胡正義,呂家瓏,等.大氣氮沉降對闊葉林紅壤淋溶水化學模擬研究[J].生態學報,2006(6):1872-1881
[7] ?MacDonald J A,Dise N B,Matzner E,et al.Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests[J].Global Change Biology,2002,8(10):1028-1033
[8] 常運華,劉學軍,李凱輝,等.大氣氮沉降研究進展[J].干旱區研究,2012,29(6):972-979
[9] 張歆陽,王喬姝怡,邢娟,等.氮添加對武夷山米櫧徑向生長的影響[J].北京大學學報:自然科學版,2017,53(6):1143-1149
[10] 鄒宇星,鐘全林,游雅玲,等.短期氮-水處理對刨花楠幼苗細根根序形態的影響[J].應用生態學報,2018,29(7):2323-2329
[11] Hyvnen R,Persson T,Andersson S,et al.Impact of longterm nitrogen addition on carbon stocks in trees and soils in northern Europe[J].Biogeochemistry,2008,89(1):121-137
[12] 潘慶民,白永飛,韓興國,等.氮素對內蒙古典型草原羊草種群的影響[J].植物生態學報,2005(2):311-317
[13] 李祿軍,曾德慧,于占源,等.氮素添加對科爾沁沙質草地物種多樣性和生產力的影響[J].應用生態學報,2009,20(8):1838-1844
[14] ?Yan G,Xing Y,Wang J,et al.Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China:Effects of nitrogen deposition[J].Agricultural & Forest Meteorology,2018,248:70-81
[15] 汪金松.模擬氮沉降對暖溫帶油松林土壤碳循環過程的影響[D].北京:北京林業大學,2013
[16] Gundersen P,Emmett B A,Kjonaas O J,et al. Impact of nitrogen deposition on nitrogen cycling in forests:A synthesis of NITREX data[J].Forest Ecology and management,1998,101(1):37-55
[17] 周利勛,劉廣平,王金波.落葉松人工林的施肥效應[J].東北林業大學學報,2004(2):16-18
[18] RappeGeorge M O,Gardenas A l,Kleja D B.The impact of four decades of annual nitrogen addition on dissolved organic matter in a boreal forest soil[J].Biogeosciences Discussions,2012,10(3):1365-1377
[19] 于美佳,葉彥輝,韓艷英,等.氮沉降對森林生態系統影響的研究進展[J].安徽農業科學,2021,49(3):19-24,27
[20] 李超.中亞熱帶九種常綠闊葉樹種幼苗葉結構型性狀相關性與葉經濟譜分析[D].北京:中國林業科學研究院,2016
[21] 周博,范澤鑫,杞金華.哀牢山中山濕性常綠闊葉林木徑向生長季節動態及其對氣候因子的響應[J].生態學報,2020,40(5):1699-1708
[22] Xu Y,Zhang Y,Yang J,et al.Influence of tree functional diversity and stand environment on fine root biomass and necromass in four types of evergreen broad-leaved forests[J].Global Ecology and Conservation,2020,21(C):e00832-e00832
[23] 《江西森林》編委會.江西森林[M].南昌:江西科學技術出版社,1986:220-230
[24] 歐陽明,楊清培,陳昕,等.毛竹擴張對次生常綠闊葉林物種組成、結構與多樣性的影響[J].生物多樣性,2016,24(6):649-657
[25] 童冉,周本智,姜麗娜,等.毛竹入侵對森林植物和土壤的影響研究進展[J].生態學報,2019,39(11):3808-3815
[26] 樂新貴.陽際峰自然保護區生態經濟評價[D].南京:南京林業大學,2005
[27] 曹軍,張鐿鋰,劉燕華.近20年海南島森林生態系統碳儲量變化[J].地理研究,2002(5):551-560
[28] ?Fang J Y, Chen A P, Peng C H, et al.Changes in forest biomass carbon storage in China between 1949 and 1998[J].Science,2001,292(5525):2320-2322
[29] 曾掌權, 田育新, 戴成棟, 等.湖南毛竹林生物量模型研究[J].湖南林業科技,2016,43(6):56-59
[30] 蔡愛明. 生物和非生物壓力對入侵和本地木本生長和競爭的影響[D].武漢:華中農業大學,2018
[31] 胡可易,柳麗娜,李雪濤,等.經營干擾對毛竹林土壤肥力及胸徑分布的影響[J].浙江林業科技,2013,33(5):52-56
[32] 洪宜聰. 毛竹純林和混交林竹筍-幼竹高生長差異分析[J]. 西南林業大學學報, 2013, 33(5): 64-68
[33] 樊江文, 鐘華平, 杜占池,等. 草地植物競爭的研究[J]. 草業學報, 2004(3):1-8
[34] ?Grime J P . Vegetation classification by reference to strategies[J]. Nature, 1974, 250:26-31
[35] 趙雨虹.毛竹擴張對常綠闊葉林主要生態功能影響[D].北京:中國林業科學研究院,2015
[36] 楊清培,楊光耀,宋慶妮,等.竹子擴張生態學研究:過程、后效與機制[J].植物生態學報,2015,39(1):110-124
[37] 徐曉,楊丹.湖南省馬尾松林生物總量的空間分布與動態變化[J].中南林業科技大學學報,2012,32(11):73-78
[38] 楊清培,王兵,郭起榮,等.大崗山毛竹擴張對常綠闊葉林生態系統碳儲特征的影響[J].江西農業大學學報,2011,33(3):529-536
[39] 杜瑩,鮑永新,裘子炎,等.模擬氮沉降對毛竹非結構性碳的影響[J].林業科學,2017,53(7):10-17
[40] 劉寧. 氮沉降和生物炭添加對毛竹生理特性的影響[D]. 臨安:浙江農林大學,2019
[41] Guo Y Y,Xing Y J,Wang J Y,et al.Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China: Effects of nitrogen deposition[J]. Agricultural and Forest Meteorology,2018,248:70-81
[42] 陶晨悅,邵珊璐,史文輝,等.氮沉降對干旱脅迫下毛竹實生苗生物量和保護酶活性的影響[J].林業科學,2019,55(9):31-40
