特發性震顫相關基因研究進展
2021-09-22 06:28:10萬雅蘭王朝霞
中國現代神經疾病雜志 2021年6期
關鍵詞:研究
萬雅蘭 王朝霞
特發性震顫(ET)亦稱為原發性震顫,是臨床常見的運動障礙性疾病,患病率為0.9%,并且隨年齡的增長而明顯升高,65歲以上人群患病率為4.6%、95歲以上人群可達20%[1]。特發性震顫主要表現為姿勢性震顫和動作性震顫,最常累及手部及上肢,亦可累及下肢、頭面部、軀干及發音。發病機制迄今尚未闡明,20世紀70年代提出的“下橄欖核模型”認為,下橄欖核中起搏神經元過度激活導致橄欖-小腦信號輸出異常,引起震顫[2];“小腦退化模型”成為近年研究熱點,該模型基于特發性震顫患者小腦存在浦肯野細胞軸突和樹突改變及浦肯野細胞移位和丟失、籃狀細胞軸突改變、攀緣纖維與浦肯野細胞連接異常分布,以及齒狀核γ-氨基丁酸受體(GABAR)改變,進而提出小腦病變是導致特發性震顫的關鍵原因[3]。基于腦電圖(EEG)、腦磁圖(MEG)和fMRI的神經網絡研究顯示,特發性震顫患者皮質-橄欖-小腦-丘腦環路活動增強,此環路將現有的特發性震顫相關理論較好地進行概括,且為腦深部電刺激術(DBS)以及周圍電刺激術提供理論基礎[4](圖1)。流行病學調查發現,約50%的特發性震顫患者有家族史,≥80%的早發型(40歲前發病)患者存在1個及以上的一級親屬患病[5],揭示遺傳因素在特發性震顫的發病過程中起重要作用。既往研究多認為特發性震顫高度遺傳且為單基因常染色體顯性遺傳性疾病,至65歲時可完全外顯[6]。但近5年的觀點認為,特發性震顫是一種基因因素與環境因素共同作用的疾病,且存在基因不完全外顯的情況[7],相關表觀遺傳學研究較少。2019年,Paul等[8]對12例特發性震顫患者進行尸檢,發現小腦甲基化改變可能與特發性震顫的發病有關。而其他表觀遺傳學機制,如組蛋白修飾、微小RNA(miRNA)表達等暫未見諸報道。盡管目前僅有特發性震顫常染色體顯性遺傳模式的報道,但仍有學者認為其可能存在雙基因或多基因致病[7]。
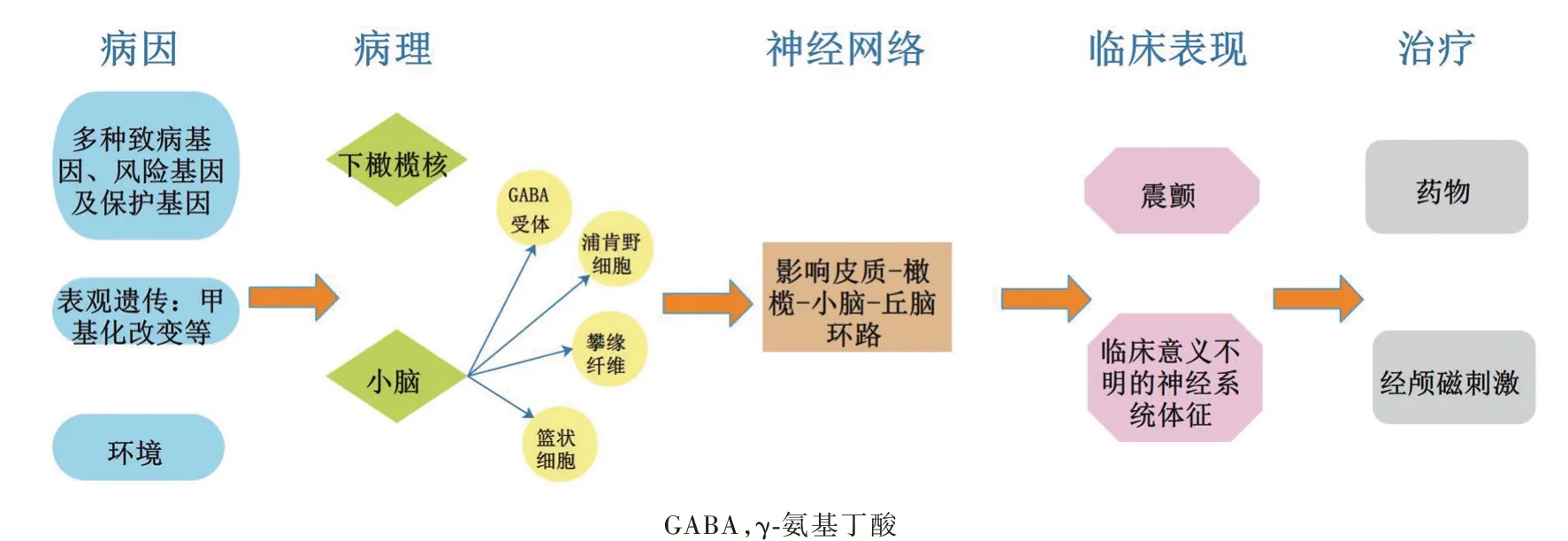
圖1 特發性震顫的疾病模式:基因、表觀遺傳及環境因素共同作用,導致下橄欖核及小腦病變,進而影響皮質-橄欖-小腦-丘腦環路,最終導致震顫,伴或不伴臨床意義不明的神經系統體征Figure 1 Disease model of essential tremor:genetic,epigenetic and environmental factors work together to cause lesions in the inferior olive nucleus and cerebellum,which in turn affect the cortical-olive-cerebellum-thalamus circuits,and ultimately lead to tremor,with or without neurological signs of uncertain clinical significance.
20世紀90年代以來,陸續有學者通過特發性震顫 家 系 連 鎖 分 析(linkage analysis)確 定ETM1、ETM2、ETM3共3個基因座;以及通過全外顯子組測序(WES)在個別患者或家系中發現FUS、TENM4、SCN4A、SCN11A、KCNS2、HTRA2、HAPLN4、SORT1、NOS3基因變異,全基因組測序(WGS)發現CACNA1G基因變異,全基因組關聯分析(GWAS)發現LINGO1、LINGO2、SLC1A2、STK32B、PPARGC1A、CTNNA3等基因單核苷酸多態性(SNP)位點與特發性震顫相關[5]。2020年,唐北沙教授團隊采用長讀長高通量測序(LRS)發現,特發性震顫患者存在NOTH2NLC基因5’非翻譯區(5'UTR)鳥嘌呤-鳥嘌呤-胞嘧啶(GGC)重復擴展突變[9](圖2),未來有望通過第三代測序技術發現其他相關基因。

圖2 特發性震顫致病基因以及風險或保護基因研究的里程碑Figure 2 Milestones in the study of essential tremor pathogenic genes and risk/protective genes.
目前已發現的特發性震顫致病基因有10余個,風險或保護基因也有10余個[5],這些基因涉及的細胞生物學功能包括調控細胞膜離子通道、小膠質細胞功能及髓鞘形成、細胞凋亡、γ-氨基丁酸(GABA)能系統、線粒體功能、細胞核與細胞質之間物質轉運等(圖3)。其中一些基因如DRD3、HS1BP3、TENM4、CACNA1G、HAPLN4,在皮質-橄欖-小腦-丘腦環路中呈高表達,其基因變異可能通過直接影響環路而引起震顫;另一些基因如FUS、USP46、LINGO1,在神經系統呈廣泛表達,但這些基因變異導致的特發性震顫卻僅表現為震顫,推測可能是由于皮質-橄欖-小腦-丘腦環路中神經元較其他腦區有一定特殊性,尚待更多基礎研究加以證實;此外,還有一些基因例如STK32B、PPARGC1A、CTNNA3,在神經系統表達較少,主要表達于其他組織器官如心臟、腎臟,其變異致震顫的作用機制鮮有報道(表1,2)[9-24]。
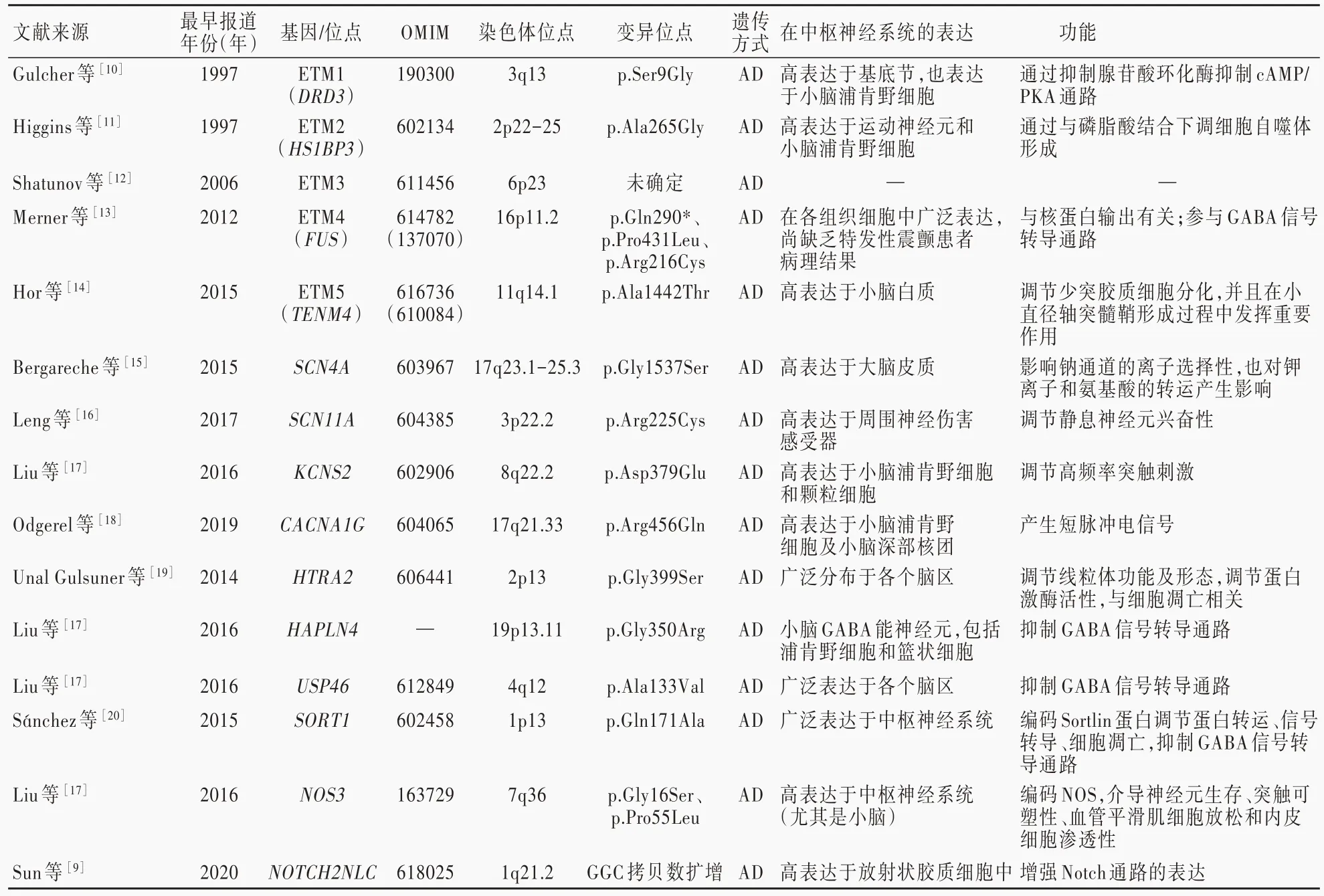
表1 特發性震顫相關致病基因Table 1.Essential tremor related pathogenic genes
一、特發性震顫相關致病基因
1.ETM1、ETM2、ETM3基因座 ETM1、ETM2和ETM3是通過連鎖分析定位的3個與特發性震顫相關的基因座。2006年,Jeanneteau等[25]在23個法國特發性震顫家系中發現位于ETM1基因座的DRD3基因p.Ser9Gly變異。DRD3基因在基底節區呈高表達,在小腦浦肯野細胞也有表達。DRD3蛋白是G蛋白耦聯受體(GPCR)多巴胺D2受體(D2R)家族成員,通過抑制腺苷酸環化酶(AC)而抑制c AMP/蛋白激酶A(PKA)信號轉導通路,而p.Ser9Gly變異可加強此抑制作用,從而導致特發性震顫[26]。2005年,Higgins等[27]在2個美國特發性震顫家系中發現位于ETM2基因座的HS1BP3基因p.Ala265Gly變異。HS1-BP3蛋白在運動神經元和浦肯野細胞呈高表達,通過與磷脂酸結合以減少細胞自噬體形成,促使細胞凋亡,從而導致特發性震顫[28]。ETM3基因座定位于6p23,理論上應該存在致病基因,但對該區域的15個基因進行測序并未確定特發性震顫相關致病基因[17],尚待進一步研究證實。
2.FUS基因FUS基因位于ETM4基因座,對基因修復、基因轉錄、RNA加工、細胞增殖均有調節作用[29]。2012年,Merner等[13]報告一特發性震顫家系存在FUS基因p.Gln290*變異。該變異可阻礙核蛋白輸出,也可通過損害GABA信號轉導通路導致運動障礙。
3.TENM4基因TENM4基因位于ETM5基因座。2015年,Hor等[14]報告3個西班牙特發性震顫家系中存在TENM4基因p.Ala1442Thr變異,但該變異在后續其他研究中未能得到驗證[30]。TENM4蛋白主要表達于小腦白質,可調節少突膠質細胞分化,并在小直徑軸突髓鞘形成過程中發揮重要作用。p.Ala1442Thr變異通過造成小腦白質損害,繼而引起特發性震顫[31]。
4.電壓門控離子通道相關致病基因 鈉、鉀、鈣離子通道相關基因與特發性震顫之間的關系于2015-2019年陸續見諸報道[15-18],這些基因多與調節神經元興奮性有關,但導致特發性震顫的具體機制尚缺乏研究證實。有研究顯示,編碼鈉離子通道α亞基Nav1.4的SCN4A基因p.Gly1537Ser變異和編碼Nav1.9的SCN11A基因p.Arg225Cys變異均與特發性震顫有關[32]。SCN4A基因高表達于肌肉以及
神經系統,通過降低丘腦神經元動作電位閾值以引起震顫。SCN4A基因變異影響鈉離子通道的離子選擇性,降低重復動作電位激發幅度,并影響鉀離子和氨基酸的轉運,從而影響丘腦相關的連接通路,引起特發性震顫[33]。SCN11A基因變異通過延長閾下刺激的去極化,調節靜息神經元的易興奮性,增強神經系統興奮性,繼而引起特發性震顫[5]。2019年,Hosoi等[34]在特發性震顫小鼠模型中發現,編碼Nav1.6的SCN8A基因與特發性震顫具有相關性,但迄今尚未在特發性震顫患者中發現這一變異。2016年,Liu等[17]報告編碼鉀離子通道α亞基Kv9.2的KCNS2基因p.Asp379Glu變異與特發性震顫有關。KCNS2基因高表達于小腦浦肯野細胞和顆粒細胞,對調節高頻率突觸刺激具有重要作用。鉀離子通道Kv3家族也在小腦浦肯野細胞表達[35],但其與特發性震顫的關系尚未見諸文獻報道。2019年,Odgerel等[18]在一特發性震顫家系中發現CACNA1G基因p.Arg456Gln變異。CACNA1G基因高表達于小腦浦肯野細胞和小腦深部核團[36],主要作用是產生短脈沖電信號,作用于非橄欖小腦束起源的運動通路,導致特發性震顫,但其具體機制尚待進一步研究[37]。
5.線粒體功能及細胞凋亡相關致病基因Unal Gulsuner等[19]于2014年在一土耳其特發性震顫大家系中發現HTRA2基因變異。當神經細胞受到凋亡性刺激時,HTRA2蛋白自線粒體膜間釋放入細胞質,參與啟動細胞凋亡[38]。HTRA2基因p.Gly399Ser變異可導致線粒體功能紊亂、形態異常,降低蛋白激酶活性,過表達此變異的細胞在應激時更易死亡。HTRA2基因p.Gly399Ser變異為功能缺失變異,其導致特發性震顫可能與紋狀體神經元缺失有關[19],但這一觀點尚缺乏相關研究證實。
6.GABA信號轉導通路相關致病基因 震顫小鼠模型顯示,注射γ-氨基丁酸A型受體(GABAAR)激動劑可減輕小鼠震顫癥狀[39],提示GABA信號轉導通路表達下調與特發性震顫的發病有關。SORT1[20]、HAPLN4[17]、USP46[17]這3個GABA信號轉導通路基因變異可能與特發性震顫的發病相關。2015年報道的一西班牙特發性震顫家系中存在SORT1基因p.Gly171Ala變異[20]。SORT1基因編碼的Sortlin蛋白在中樞神經系統廣泛表達,Sortlin蛋白不僅通過調節蛋白轉運和信號轉導調節神經元功能,還通過與神經生長因子前體結合調控神經元存亡。SORT1基因p.Gly171Ala變異可導致Sortlin蛋白減少,而與Sortlin結合的P75NTR蛋白增多,后者增多見于GABA信號轉導通路受損[5]。2016年的一項針對37個早發型特發性震顫家系的研究發現HAPLN4基因p.Gly350Arg變異及USP46基因p.Ala133Val變異[17]。推測上述3個基因變異均通過下調GABA信號轉導通路而導致特發性震顫,但二者之間的相關性尚缺乏多中心大樣本研究驗證。
7.一氧化氮信號轉導相關基因 一氧化氮在神經系統中參與介導神經元的生存、突觸可塑性、血管平滑肌細胞松弛和血管內皮細胞滲透性[40]。2016年,Liu等[17]對37個早發型特發性震顫家系進行研究并報告NOS3基因p.Gly16Ser及p.Pro55Leu變異。NOS3基因高表達于中樞神經系統(尤其是小腦)神經元和血管內皮細胞,編碼一氧化氮合酶(NOS),其導致特發性震顫的具體機制尚待進一步研究[40]。
8.NOTCH2NLC基因三核苷酸拷貝數擴增突變 2020年,我國學者Sun等[9]通過LRS測序對197個特發性震顫家系進行研究,發現NOTCH2NLC基因5'UTR GGC拷貝數擴增,約5.58%的患者攜帶該變異。NOTCH2NLC基因高表達于神經系統放射狀膠質細胞,主要作用是增強Notch信號轉導通路的表達,從而對大腦發育、神經元增殖和分化發揮至關重要的作用。但是位于NOTCH2NLC基因5'UTR的動態變異可能并非通過導致編碼蛋白異常表達而致病,其作用機制更加復雜,目前有RNA毒性作用、重復相關非ATG翻譯等假說[41],尚待進一步研究驗證。
二、特發性震顫相關風險和保護基因
1.特發性震顫相關風險基因 (1)LINGO基因家族:2009年,Stefansson等[21]在452例冰島特發性震顫患者中發現LINGO1基因rs9652490和rs11856808單核苷酸多態性位點。全基因組的單核苷酸多態性微陣列分析發現,LINGO1基因在特發性震顫患者中存在拷貝數擴增[42]。LINGO1基因廣泛分布于不同腦區,通過與P75NTR或腫瘤壞死因子受體超家族成員19(TROY19)共同形成軸突生長抑制蛋白-66受體1(NgR1)復合物,參與神經元分化、少突膠質細胞成熟、抑制軸突生長、加速多巴胺能神經元變性死亡。病理學研究表明,特發性震顫患者LINGO1基因主要表達于小腦籃狀細胞軸突內,在浦肯野細胞軸突起始部分形成刷狀結構,從而影響浦肯野細胞功能[5]。此外,LINGO1蛋白作為鈣離子激活的鉀離子通道亞單位,可以引起鉀離子通道的功能性敲除,導致特發性震顫及運動障礙[43]。2010-2011年,陸續有學者發現LINGO2基因rs1412229、rs7033345、rs10812774單核苷酸多態性位點[22]。LINGO2基因僅表達于中樞神經系統,關于其引起特發性震顫的研究甚少,有學者認為其與LINGO1基因功能相似[22]。(2)其他熱點特發性震顫風險基因:2012年,Thier等[23]對990例特發性震顫患者和1537例健康對照者進行病例對照研究,發現SLC1A2基因rs3794087單核苷酸多態性位點。SLC1A2基因編碼的谷氨酸再攝取轉運蛋白興奮性氨基酸轉運體2(EAAT2)主要作用為將谷氨酸從突觸間隙中移除,從而解除谷氨酸的神經毒性作用。SLC1A2基因表達于浦肯野細胞軸突起始部分,在特發性震顫患者小腦皮質表達明顯減少[5]。2016年,Müller等[24]發 現 ,PPARGC1A基 因rs17590046及CTNNA3基因rs12764057、rs10822974、rs7903491單核苷酸多態性位點均可增加特發性震顫的患病風險,但并未在后續研究中得到證實[44]。PPARGC1A基因編碼過氧化物酶增殖激活受體γ共受體1α(PGC-1α),通過與解耦聯蛋白2及核呼吸因子相互作用調節線粒體功能及能量代謝,尸檢研究顯示,帕金森病患者黑質及蒼白球PGC-1α異構體表達升高[45]。CTNNA3基因編碼的α降鈣素3在細胞-細胞黏附過程中發揮重要作用,但其導致特發性震顫的機制尚待進一步研究驗證[44]。
2.特發性震顫相關保護基因 2016年,Müller等[24]對2807例歐洲特發性震顫患者及6441例健康對照者進行病例對照研究,發現STK32B基因rs10937625單核苷酸多態性位點是特發性震顫的保護因素,且這一結論在中國學者針對218例特發性震顫患者的GWAS研究中得以驗證[46]。STK32B基因編碼絲氨酸/蘇氨酸激酶,但其功能尚不十分清楚。推測STK32B基因在特發性震顫患者小腦皮質的表達升高,而rs10937625單核苷酸多態性位點可以導致STK32B基因在小腦皮質的表達減少,從而成為特發性震顫的保護因素。
三、總結與展望
特發性震顫的遺傳特征復雜,致病基因譜十分寬泛[47]。值得注意的是,目前已報道的致病基因在特發性震顫患者中的陽性檢出率均很低,尚未發現高頻率的致病基因,因此特發性震顫并非某一種單一致病基因的疾病,而是一組遺傳異質性譜系疾病。此外,疾病易感基因和表觀遺傳修飾也與特發性震顫的發病有關。因此,特發性震顫的診斷應以病史、臨床表現及體格檢查為基礎,對經濟條件允許的患者可進行基因檢測以輔助診斷與鑒別診斷,但不能過度依賴基因檢測結果。在對基因檢測結果進行分析時,應注意測序結果的判讀,對檢測到的變異應結合既往報道謹慎分析是否為致病性變異。總之,特發性震顫相關基因研究有助于臨床醫師對疾病的認識更加深入。未來研究方向為,通過新技術發現更多新的變異基因及位點、深入研究基因變異致病機制的同時,緊密結合臨床,闡釋不同變異位點與發病年齡、臨床癥狀、藥物反應的關系,從而為精準治療提供線索。
利益沖突無
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
