野生垂穗披堿草成苗期間的耐旱性研究
2021-09-22 08:55:50王傳旗劉文輝張永超周青平
草業(yè)學報 2021年9期
王傳旗,劉文輝,張永超,周青平,2*
(1.青海省青藏高原優(yōu)良牧草種質(zhì)資源利用重點實驗室,青海大學畜牧獸醫(yī)科學院,青海 西寧810016;2.西南民族大學,四川 成都610041)
全球氣溫升高加劇了干旱脅迫,干旱是一個長期存在的全球性生態(tài)環(huán)境問題。相對于其他非生物逆境,干旱具有出現(xiàn)次數(shù)多、持續(xù)時間長、影響范圍大等特點,對農(nóng)業(yè)生產(chǎn)的影響很大,即使輕微的干旱也會降低糧食產(chǎn)量。我國是一個水資源十分匱乏的國家,干旱和半干旱地區(qū)面積占國土面積的47%,占總耕地面積的51%,且在地理上主要分布于西北地區(qū)[1]。然而,即使在我國的濕潤及半濕潤地區(qū),也會時常遭受難以預測的短暫旱災。干旱已對我國經(jīng)濟和社會造成了嚴重的損失。
“三江源”地區(qū)位于青海省南部,地處“世界屋脊”青藏高原的腹地,平均海拔3500~4800 m,素有我國乃至亞洲“水塔”之美譽,不僅是我國長江、黃河流域生態(tài)環(huán)境安全和社會經(jīng)濟可持續(xù)發(fā)展的生態(tài)屏障,其生態(tài)環(huán)境狀態(tài)及變化同時具有全球指標意義[2-3]。“三江源”地區(qū)屬高原大陸性氣候,自然環(huán)境嚴酷,具有低溫、缺氧、輻射強烈、干旱、風沙等氣候特征,是我國面積最大的自然保護區(qū),也是全世界所關(guān)注的生態(tài)脆弱地區(qū)之一。長期以來,“三江源”地區(qū)受自然因素和人類活動影響,該地區(qū)生態(tài)環(huán)境問題依然十分嚴峻,其中大面積的草(場)地退化就是其眾多生態(tài)環(huán)境問題的主要表現(xiàn)之一[4-6]。此外,由于春旱頻繁,牧草的播種成苗也是“三江源”地區(qū)在人工草地建植、草地補播改良和草原生態(tài)修復中存在的一個重要問題。
鑒于此,本研究選取了適宜于“三江源”地區(qū)高寒草地生態(tài)恢復的野生垂穗披堿草(Elymus nutans)為試驗材料,研究了其成苗期間的耐旱性及需水特性,研究結(jié)果將有助于揭示垂穗披堿草干旱適應機理,同時對“三江源”地區(qū)植被恢復和利用垂穗披堿草有效防治土地沙化、水土流失等災害具有重要的科學價值和現(xiàn)實意義。
1 材料與方法
1.1 試驗材料及區(qū)域概況
野生垂穗披堿草采集于青海省海南藏族自治州貴南縣茫拉鄉(xiāng)(35°12′N,100°45′E,海拔3216 m),高寒草原路邊。該地區(qū)屬高原大陸性氣候,年均氣溫為2.3℃,年均日照時數(shù)為2907.8 h,年均降水量為403.8 mm,年均蒸發(fā)量為1378.5 mm,干旱、沙塵暴等自然災害頻繁。采集來的野生種子在青海省海北州西海鎮(zhèn)多年生牧草種質(zhì)資源圃(36°54′35″N,101°49′43″E,海拔3156 m),進行馴化選育,編號I-1-3-5。試驗材料為野生馴化品系。試驗前在室溫(溫度16~23℃,相對濕度53%)條件下進行了預試驗,測得垂穗披堿草種長(1.77±0.03)㎝,種寬(0.13±0.01)㎝,千粒重(5.61±0.05)g,種子含水量8.6%,平均發(fā)芽率97%。
1.2 試驗地點和時間
本項研究試驗在青海大學畜牧獸醫(yī)科學院-青藏高原優(yōu)良牧草種質(zhì)資源利用重點實驗室進行并順利完成。試驗于2020年6月16日開展,6月27日開始干旱脅迫及復水試驗,試驗共歷時28 d。
1.3 試驗設(shè)計
配制水勢為0、-0.15、-0.30、-0.50、-0.75和-1.00 MPa的高分子滲透劑聚乙二醇(PEG-6000)溶液模擬干旱環(huán)境[7]。試驗采用紙上發(fā)芽法,在每個鋪有雙層濾紙的培養(yǎng)皿中排25粒種子并加7 mL不同環(huán)境水勢的PEG-6000溶液。每處理4次重復。將培養(yǎng)皿置于RTOP智能光照培養(yǎng)箱(RTOP-310B,浙江)中,發(fā)芽條件25℃/20℃(晝溫/夜溫),光周期14 h,光照度1250 lx。試驗以根長與種子等長視為發(fā)芽,以芽長2 cm視為出苗。每天統(tǒng)計發(fā)芽數(shù)和出苗率并及時清理發(fā)霉腐爛的種子。此外,試驗期間每隔2 d更換一次濾紙,并加入7 mL對應PEG-6000溶液,以避免種子吸收和水分蒸發(fā)而造成環(huán)境水勢改變。垂穗披堿草種子培養(yǎng)7 d后,將未萌發(fā)的種子用超純水(0 MPa)沖洗干凈并分別轉(zhuǎn)移至加有超純水的培養(yǎng)皿中,繼續(xù)觀察10 d結(jié)束試驗。試驗結(jié)束時,檢測未萌發(fā)種子的生活力,具體操作:用刀片將種子切開,在顯微鏡下觀察,如果胚為固體、白色,視為有生活力種子;如果胚為液體、褐色則為無生活力種子[8]。檢查結(jié)果為97%以上未萌發(fā)的種子是有活力的。
1.4 測定指標與方法
發(fā)芽率=第7天種子發(fā)芽數(shù)/試驗供試種子數(shù)×100%。出苗率=第7天出苗數(shù)/試驗供試種子數(shù)×100%。發(fā)芽指數(shù)為第t天的發(fā)芽數(shù),Dt為相對應天數(shù)[9]。活力指數(shù)均根長+平均芽長[10]。
根長和芽長的測量:從各處理的培養(yǎng)皿中隨機取出15株幼苗,進行根長和芽長的測量,取平均值[11]。
植株根和芽的干鮮重測量:干旱脅迫和復水后,在各溶液濃度處理下隨機取出10株幼苗剪下根和芽,稱重,得其鮮重;然后在105℃烘箱內(nèi)殺青30 min,再在80℃下烘至恒重,得其干重[12]。測量過程中,各處理下的幼苗不夠10株的全部都測。干物質(zhì)積累率=植株干重/植株鮮重。
種子發(fā)芽出苗需水閾值:根據(jù)公式Pi=alogt+b[13],求出各處理下50%的種子群體發(fā)芽出苗的時間t,繼而依據(jù)公式1/t=aψw+b[14],得出50%種子群體在發(fā)芽出苗過程中忍耐的最大水勢(需水閾值)。式中:Pi為種子發(fā)芽出苗百分率的概率單位轉(zhuǎn)換值;t為從發(fā)芽試驗開始到該概率出現(xiàn)的時間;ψw為水勢;-a/b為50%的種子群體發(fā)芽出苗所能忍耐的最大水勢(臨界水勢)。
1.5 數(shù)據(jù)統(tǒng)計與分析
采用Excel 2020和SPSS 19.0統(tǒng)計分析軟件對所測數(shù)據(jù)進行統(tǒng)計分析。采用Tukey’s HSD檢驗,對垂穗披堿草處理間的差異性進行單因素方差分析和Duncan多重比較分析。所有數(shù)據(jù)在分析前要滿足正態(tài)分布和方差齊性。
2 結(jié)果與分析
2.1 干旱脅迫及復水對種子發(fā)芽進程的影響
隨著環(huán)境水勢的降低,垂穗披堿草種子發(fā)芽速率逐漸降低(圖1)。-0.15和-0.30 MPa的水分虧缺對種子發(fā)芽速率影響較小,且種子的最終發(fā)芽率均大于90%;-0.75和-1.00 MPa的水分虧缺對種子發(fā)芽速率影響較大,但復水后種子最終發(fā)芽率仍達到50%以上。
水分虧缺和復水后的各處理下,種子均在第2天開始發(fā)芽(圖1)。對照組、-0.15 MPa和-0.30 MPa處理下,種子發(fā)芽高峰期均在第4天;-0.50 MPa處理下,種子發(fā)芽高峰期在第5天;-0.75和-1.00 MPa處理下,種子發(fā)芽高峰期在第6天。高濃度的PEG-6000處理(-0.50、-0.75和-1.0 MPa)延遲了垂穗披堿草種子群體發(fā)芽的高峰期。復水后,-0.15和-0.30 MPa處理下,種子群體發(fā)芽高峰均在第9天;-0.50 MPa處理下,種子群體發(fā)芽高峰在第10天;-0.75和-1.0 MPa處理下,種子群體發(fā)芽高峰均在第11天。說明復水后,隨著環(huán)境水勢的降低,垂穗披堿草種子群體發(fā)芽高峰期同樣存在滯后現(xiàn)象。
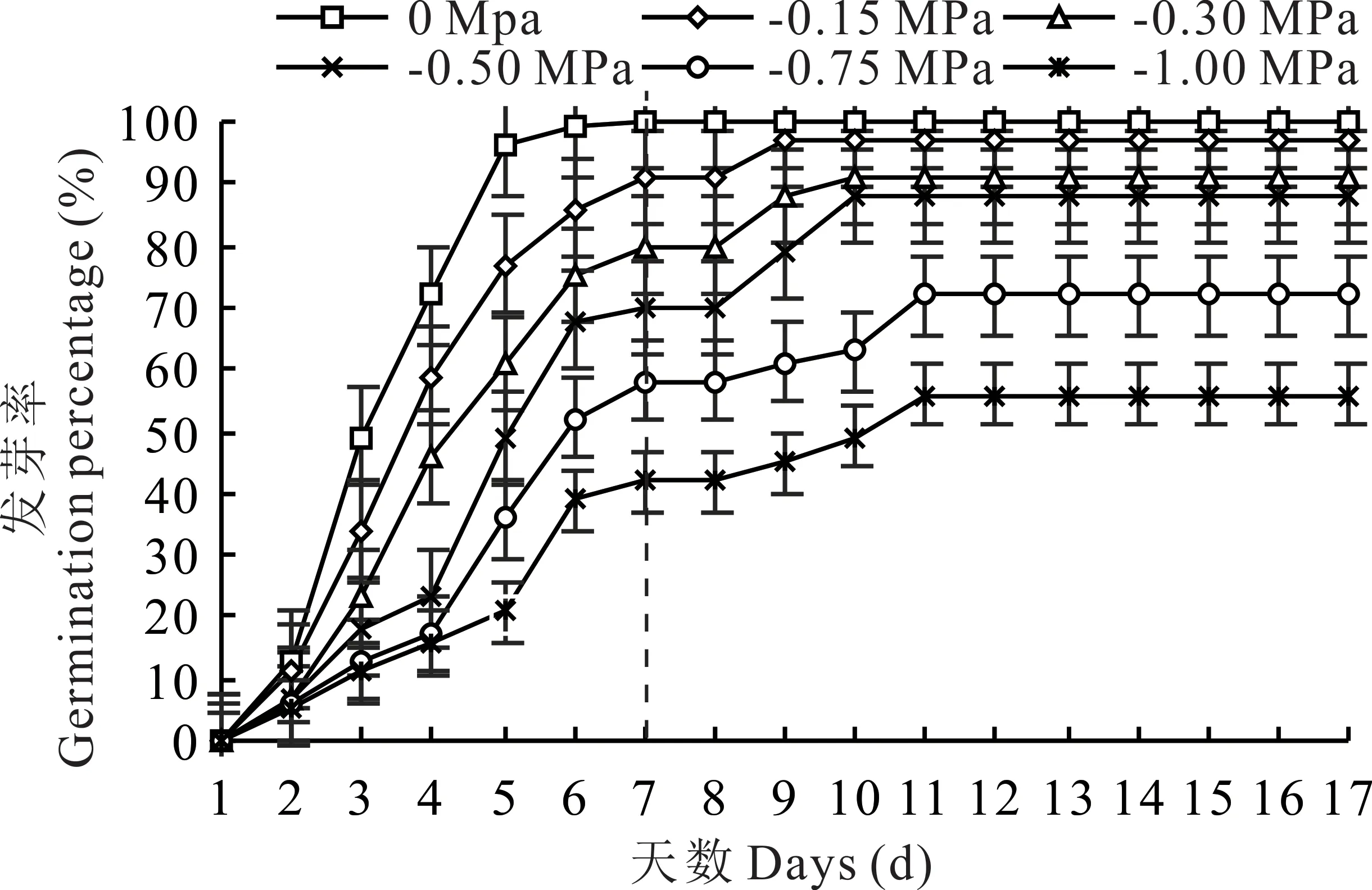
圖1 干旱脅迫及復水對種子發(fā)芽進程的影響Fig.1 Effects of drought stress and rewatering on seed germination process
2.2 干旱脅迫及復水對植物幼苗形成進程的影響
與發(fā)芽進程相似,隨著環(huán)境水勢的降低,幼苗形成速率減緩(圖2)。復水后,除對照組外,各處理下的幼苗形成率均有不同程度的增加。其中,-0.30和-0.50 MPa處理下,垂穗披堿草出苗率增幅較高。-0.75和-1.00 MPa處理下,垂穗披堿草出苗率雖有所增加,但與其他滲透處理相比,其出苗率明顯較低。
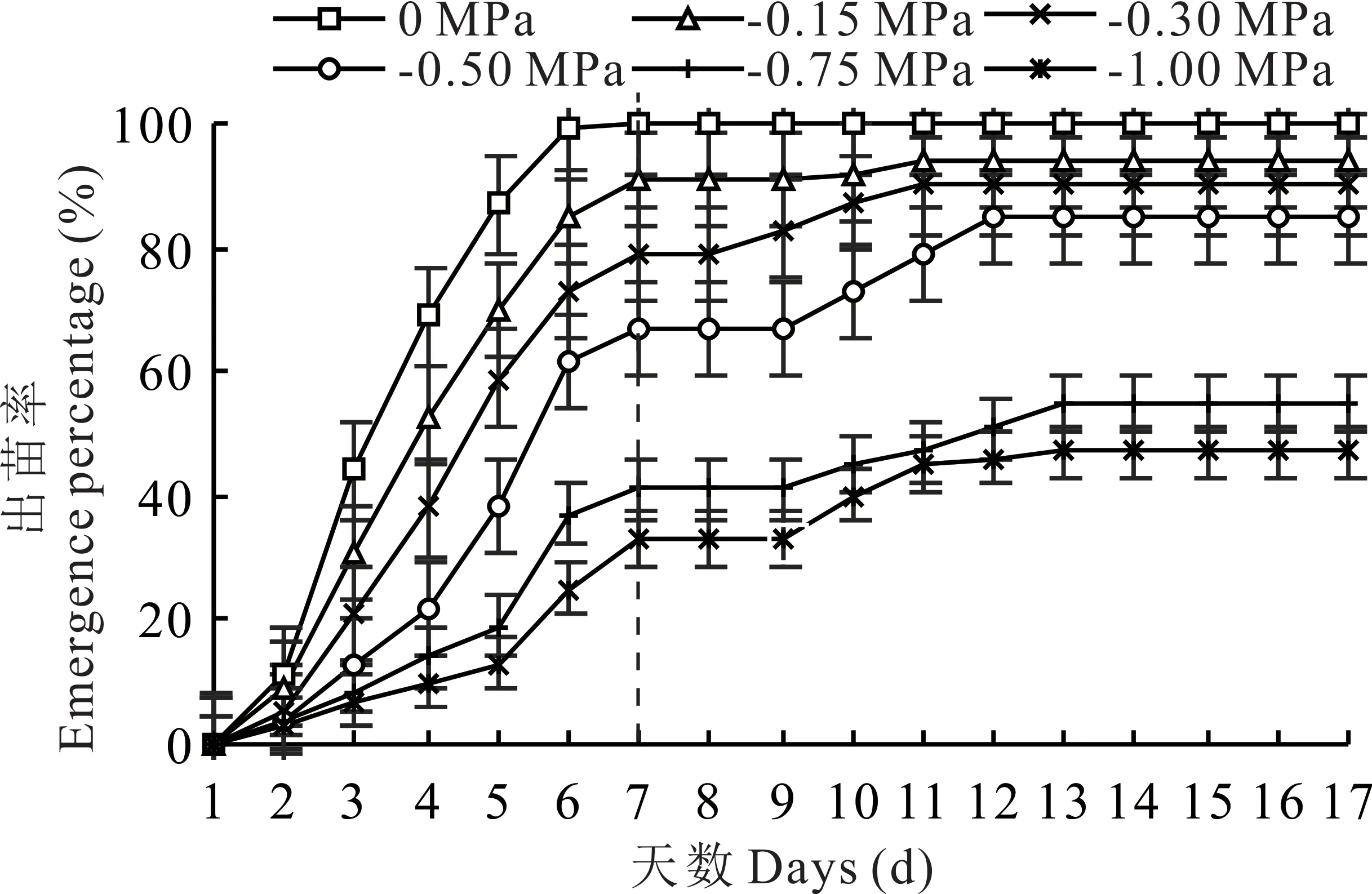
圖2 水分虧缺及復水對種子出苗進程的影響Fig.2 Effects of drought stress and rewatering on seed emergence process
對照組、-0.15和-0.30 MPa處理下,種子群體出苗高峰期在第5天;-0.50、-0.75和-1.00 MPa處理下,種子群體出苗高峰期在第6天。-1.00 MPa處理下,種子群體出苗高峰期在第6和7天。高濃度的PEG-6000處理(-0.50、-0.75和-1.00 MPa)延遲了垂穗披堿草種子群體出苗的高峰期。復水后,-0.15和-0.30 MPa處理下,種子群體出苗高峰期在第10天;-0.50 MPa處理下,種子群體出苗高峰期在第11天;-0.75和-1.00 MPa處理下,種子群體出苗高峰期均在第12天。說明復水后,隨著環(huán)境水勢的降低,垂穗披堿草種子群體出苗高峰期同樣存在滯后現(xiàn)象。
2.3 干旱脅迫及復水對種子發(fā)芽出苗的影響
垂穗披堿草種子發(fā)芽率、出苗率、發(fā)芽指數(shù)和活力指數(shù)均隨環(huán)境水勢的降低而下降(表1)。-0.15 MPa的水分虧缺對種子發(fā)芽出苗基本沒有影響;-0.50 MPa處理下,種子仍可大量發(fā)芽出苗,但其發(fā)芽指數(shù)和活力指數(shù)較對照組下降顯著(P<0.05);-0.75 MPa處理下,與對照組相比,種子發(fā)芽率、出苗率、發(fā)芽指數(shù)均下降顯著(P<0.05)。
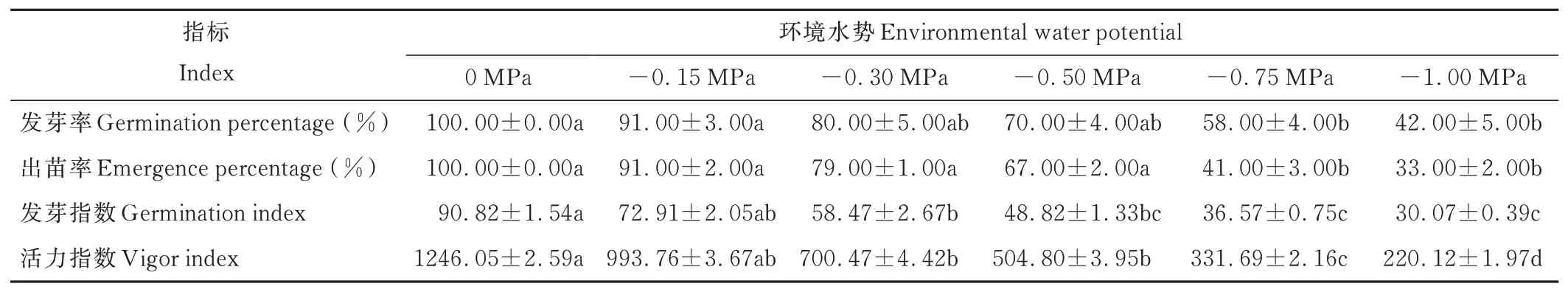
表1 干旱脅迫對種子發(fā)芽和出苗的影響Table 1 Effects of drought stress on seed germination and emergence
復水后,與對照組相比,各滲透處理下的種子發(fā)芽率、出苗率、發(fā)芽指數(shù)和活力指數(shù)均顯著升高(P<0.05)。其中,-0.50 MPa處理下,種子發(fā)芽率、出苗率、發(fā)芽指數(shù)和活力指數(shù)最高,其次為-0.75 MPa處理組,而-0.15 MPa處理組各指標值相對最低。此外,復水后,-1.00 MPa處理下,種子最終發(fā)芽率和出苗率分別達到對照組的56%和47%(表2)。
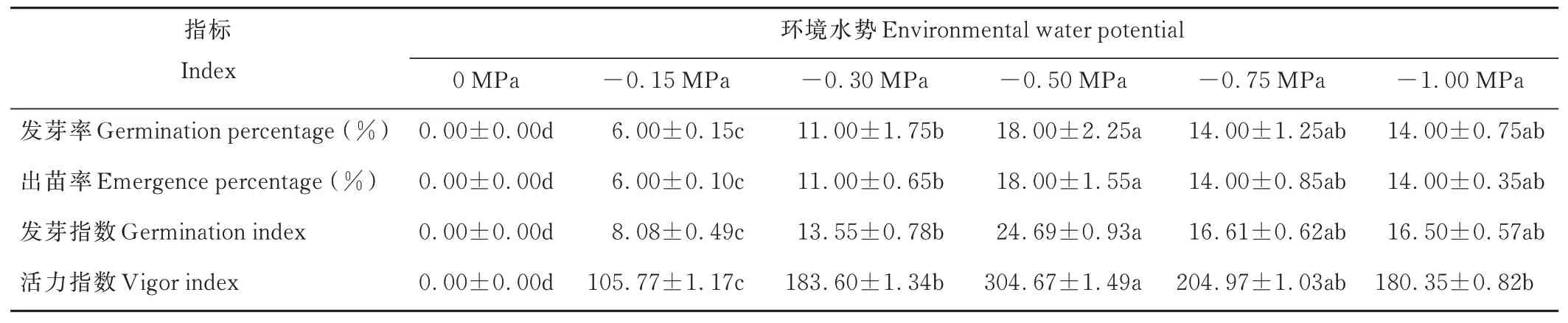
表2 旱后復水對種子發(fā)芽和出苗的影響Table 2 Effects of rewatering after drought stress on seed germination and emergence
2.4 干旱脅迫及復水對垂穗披堿草幼苗生長量的影響
垂穗披堿草芽長隨著環(huán)境水勢的降低而下降,其根長隨著環(huán)境水勢的降低則呈先升后降的趨勢(表3)。-0.15 MPa處理下,垂穗披堿草根長較對照組略長;其他處理組的根長均隨著環(huán)境水勢的降低而下降,但與對照組相比,其下降幅度均不顯著。-0.75 MPa處理下,與對照組相比,垂穗披堿草芽長下降顯著(P<0.05)。隨著環(huán)境水勢的降低,垂穗披堿草根的干鮮重呈先升后降的趨勢,其芽的干鮮重和干物質(zhì)積累率呈下降的趨勢(表4)。-0.50 MPa處理下,與對照組相比,幼苗芽的干重下降顯著(P<0.05);-0.75 MPa處理下,幼苗芽的鮮重和根的干重較對照組下降顯著(P<0.05);-1.00 MPa處理下,幼苗根的鮮重較對照組下降顯著(P<0.05)。
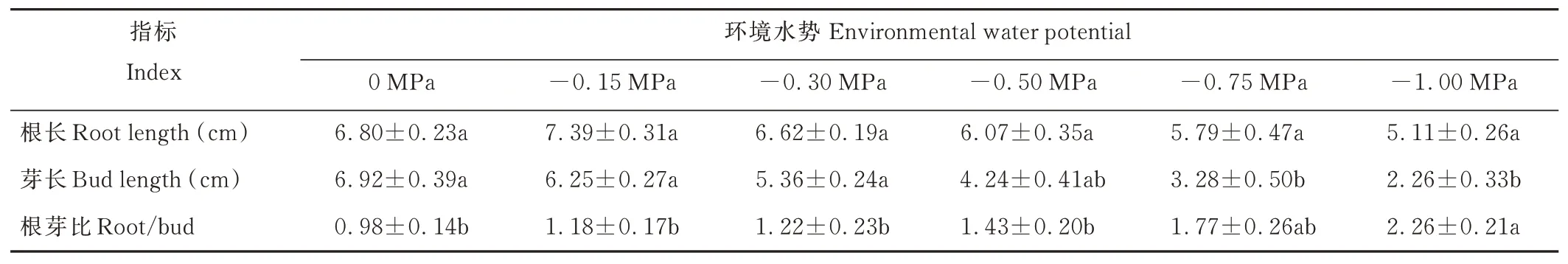
表3 干旱脅迫對垂穗披堿草幼苗根長和芽長的影響Table 3 Effects of drought stress on root length and bud length of E.nutans seedlings
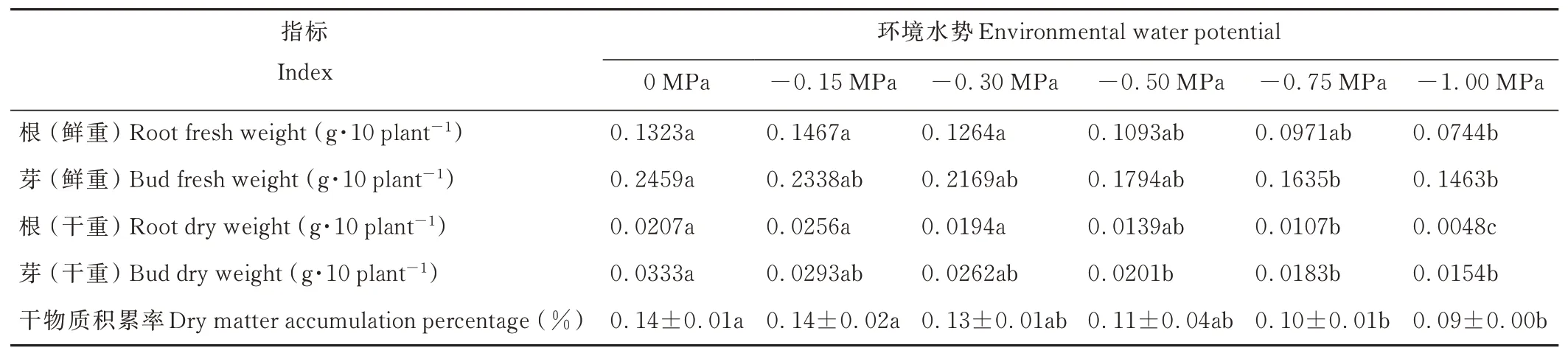
表4 干旱脅迫對垂穗披堿草幼苗干鮮重的影響Table 4 Effects of drought stress on fresh and dry weight of E.nutans seedlings
復水后,垂穗披堿草根和芽的伸長及其干鮮重和干物質(zhì)積累率均得到了較大提升(表5和表6)。-0.15 MPa處理下,與其他各處理相比,垂穗披堿草幼苗根長、芽長最大,但差異不顯著;-0.30 MPa及以上的水分虧缺處理下,幼苗根長、芽長均有略微的下降。復水后,隨著環(huán)境水勢的降低,幼苗根芽比呈先升后降的趨勢,其中-0.50 MPa處理下,幼苗根芽比相對較大。復水后,各處理下的幼苗干物質(zhì)積累率基本一致且與復水前對照組的幼苗干物質(zhì)積累率相當。
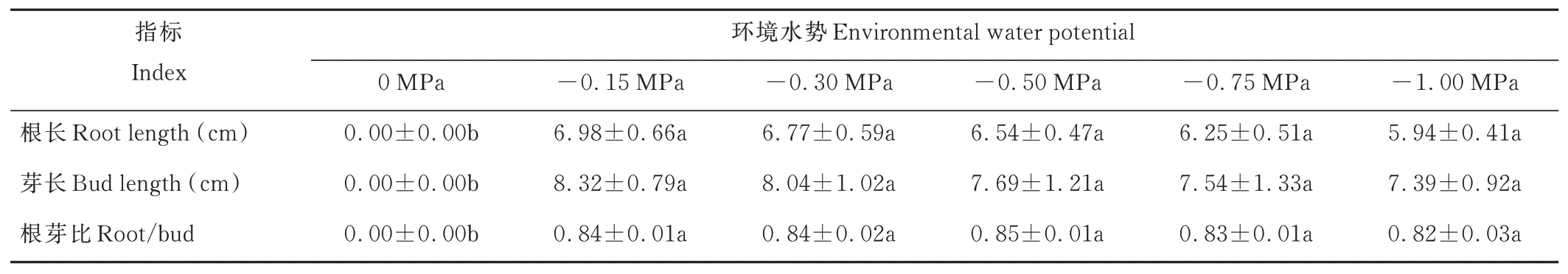
表5 旱后復水對垂穗披堿草幼苗根長和芽長的影響Table 5 Effects of rewatering after drought stress on root length and bud length of E.nutans seedlings
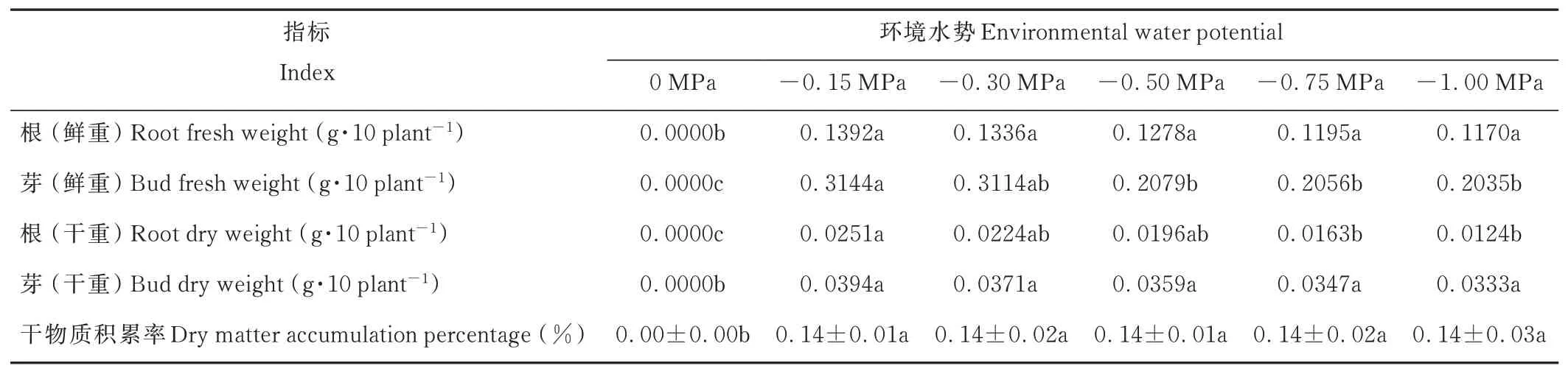
表6 旱后復水對垂穗披堿草幼苗干鮮重的影響Table 6 Effects of rewatering after drought stress on dry and fresh weight of E.nutans seedlings
2.5 干旱脅迫及復水下種子發(fā)芽出苗需水閾值研究
為了闡明不同PEG-6000脅迫處理下的野生垂穗披堿草50%種子群體發(fā)芽和出苗階段的水勢臨界值,本項研究對不同干旱脅迫處理下的垂穗披堿草種子發(fā)芽和出苗階段的概率與相應的時間對數(shù)進行擬合,然后利用最小二乘法進行曲線擬合,最后根據(jù)公式1/t=aψw+b,得出50%種子群體在其發(fā)芽出苗過程中忍耐的最大水勢值(表7~10)。
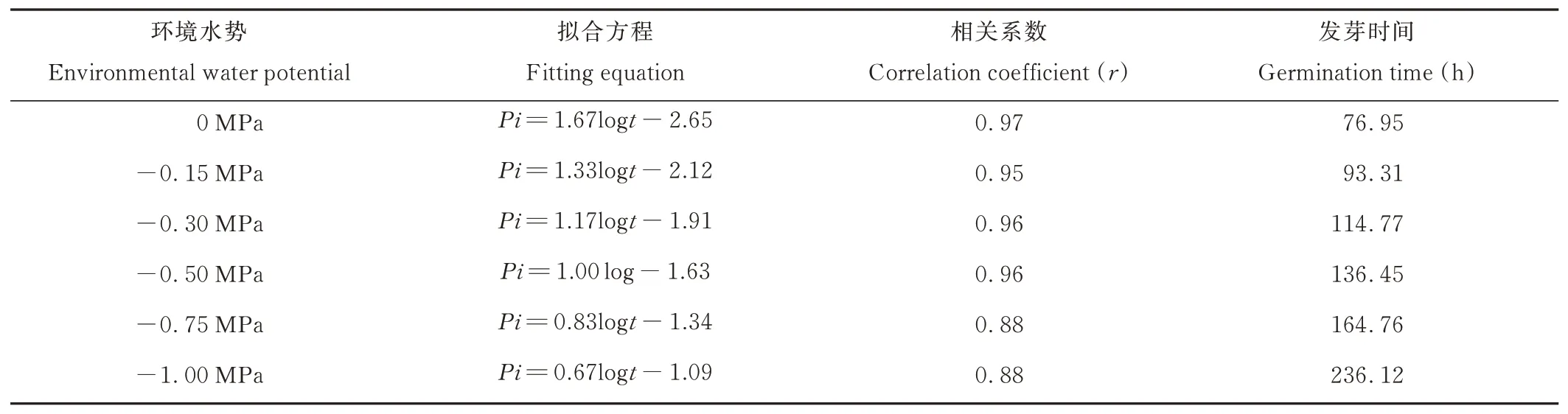
表7 50%種子群體吸水時間與發(fā)芽率的關(guān)系Table 7 Relationship between water absorption time and germination percentage of 50% seeds population

表8 50%種子群體發(fā)芽時間與水勢的關(guān)系Table 8 Relationship between germination time and water potential of 50% seeds population
0 MPa處理下,垂穗披堿草50%種子群體達到發(fā)芽階段所需要的時間為76.95 h。-0.15、-0.30、-0.50、-0.75和-1.00 MPa處理下,50%種子群體達到發(fā)芽階段所需要的時間分別比對照組增加了21.26%、49.15%、77.32%、114.11%和206.85%。進一步分析可知,垂穗披堿草50%種子群體達到發(fā)芽階段所能忍耐的最大水勢為-0.85 MPa。
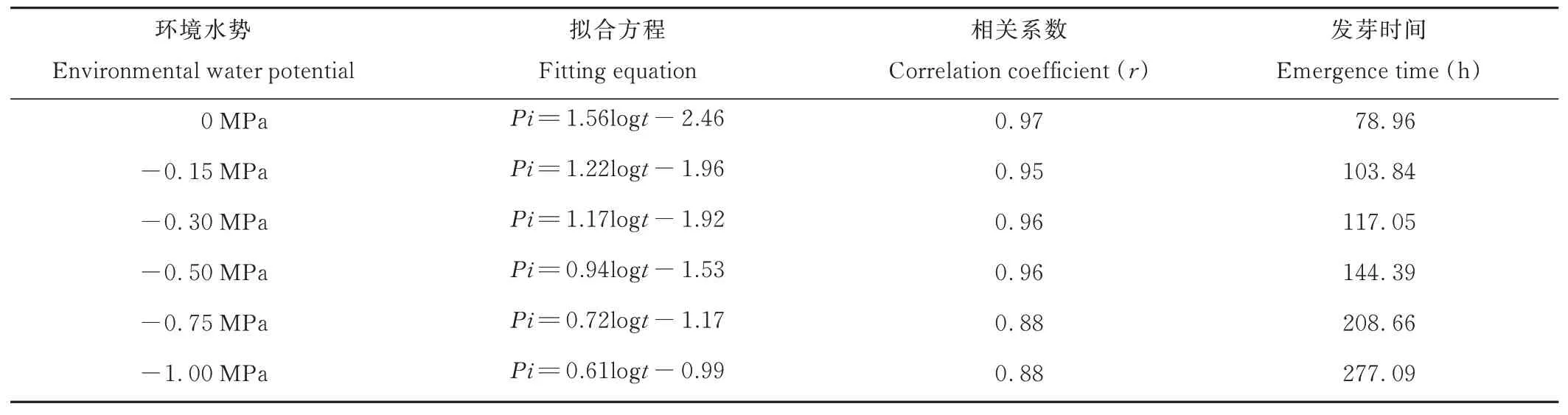
表9 50%種子群體吸水時間與出苗率的關(guān)系Table 9 Relationship between water absorption time and emergence percentage of 50% seeds population

表10 50%種子群體出苗時間與水勢的關(guān)系Table 10 Relationship between emergence time and water potential of 50% seeds population
0 MPa處理下,垂穗披堿草50%種子群體達到出苗階段所需要的時間為78.96 h。-0.15、-0.30、-0.50、-0.75、-1.00 MPa處理下,50%種子群體達到出苗階段所需要的時間分別比對照組增加了31.51%、48.24%、82.86%、164.26%和250.92%。進一步分析可知,垂穗披堿草50%種子群體達到出苗階段所能忍耐的最大水勢為-0.61 MPa。
以上表明,PEG-6000干旱脅迫對垂穗披堿草發(fā)芽和出苗的影響表現(xiàn)在延遲種子群體發(fā)芽和出苗時間。此外,從垂穗披堿草發(fā)芽和出苗階段的臨界水勢相比,其出苗比發(fā)芽階段對環(huán)境水勢的要求更高。
3 討論
3.1 干旱脅迫及復水對垂穗披堿草發(fā)芽出苗的影響
水分是干旱及半干旱地區(qū)最為關(guān)鍵和敏感的生態(tài)因子,也是影響我國西北地區(qū)作物和牧草播種成苗的主要因素。本項研究采用PEG-6000溶液模擬不同程度的干旱環(huán)境,研究了垂穗披堿草成苗期間的耐旱性及需水特性。本研究中,垂穗披堿草種子發(fā)芽進程和幼苗形成進程隨著環(huán)境水勢的降低而表現(xiàn)為種子群體發(fā)芽出苗速率降低和種子群體發(fā)芽出苗高峰期推遲。這與許翩翩等[15]對3種常見邊坡植物對模擬干旱環(huán)境抗旱性能的研究結(jié)果相一致。復水后,種子發(fā)芽率和出苗率均得到不同程度提高,其中-0.15、-0.30和-0.50 MPa處理下,種子的最終發(fā)芽率和出苗率均大于80%;即使-0.75 MPa處理,種子最終發(fā)芽率和出苗率仍可達到50%以上。表明干旱脅迫通過限制種子水分吸收以降低或減緩種子內(nèi)部酶的活性和物質(zhì)代謝過程,最終抑制種子發(fā)芽出苗速率。
眾多研究表明,隨PEG-6000溶液環(huán)境水勢的降低,種子發(fā)芽率、出苗率、發(fā)芽指數(shù)和活力指數(shù)均呈下降的趨勢[16-19]。本試驗也證實了這一點。-0.30 MPa處理下,與對照組相比,種子發(fā)芽指數(shù)和活力指數(shù)下降顯著(P<0.05);-0.75 MPa處理下,種子發(fā)芽率、出苗率較對照組下降顯著(P<0.05)。然而,本試驗中,-0.15 MPa的水分虧缺對種子發(fā)芽出苗影響較小;復水后,-0.50 MPa的水分虧缺對種子發(fā)芽率和出苗率影響較小,均可達到80%以上水平。這說明垂穗披堿草在成苗期間具有較強的耐旱性,而旱后復水對垂穗披堿草播種保苗具有重要作用。
3.2 干旱脅迫及復水對垂穗披堿草生長量的影響
根芽長、根芽比、根芽干鮮重及干物質(zhì)積累率均為反映垂穗披堿草耐旱性的重要生長量指標。本研究中,-0.15 MPa處理下,幼苗的根較對照組略長,根的干鮮重和幼苗干物質(zhì)積累率均比對照組高。說明輕度干旱脅迫更有利于垂穗披堿草幼苗的生長,同時證明水分虧缺并不一定導致植物產(chǎn)量下降。-0.75 MPa處理下,垂穗披堿草幼苗生長量均受到嚴重抑制。有研究表明,植物發(fā)芽成苗特性與其自然分布生境密切相關(guān),這是植物與自然環(huán)境趨同進化的結(jié)果[20-21]。本研究中,水分虧缺對垂穗披堿草幼苗根的伸長影響較小,但對芽的伸長影響較大。-0.15 MPa以下的水分虧缺處理,垂穗披堿草幼苗根芽比越來越大,且在-1.00 MPa處理下,較對照組差異顯著(P<0.05);而其干物質(zhì)積累率越來越小,且在-0.75 MPa處理下,較對照組下降顯著(P<0.05)。表明重度以上的干旱脅迫下,垂穗披堿草以犧牲芽為代價將有限資源優(yōu)先用于根部伸長。這可能是干旱脅迫對幼苗生長造成傷害,垂穗披堿草為了生存而采取的被動適應策略,同時這也是野生垂穗披堿草對其分布生境長期適應的結(jié)果。
山侖[22]的研究指出,在不超過作物缺水的適應范圍時,復水后往往可在生長和產(chǎn)量上產(chǎn)生補償效應,最終在節(jié)約大量用水的同時,其產(chǎn)量不受影響或影響較小。本研究中,除對照組外,復水后與復水前相比,各處理下的幼苗根芽長均有不同程度的增加,且幼苗干物質(zhì)積累率與復水前的對照組相當,說明即使是極度干旱脅迫的-1.00 MPa處理也沒有超出垂穗披堿草幼苗對干旱脅迫的適應范圍。
3.3 干旱脅迫及復水下垂穗披堿草發(fā)芽和出苗需水閾值
有研究報道,干旱脅迫下的種子發(fā)芽和出苗均有一定的閾值要求,低于其閾值便不能達到相應的階段[23-24]。本研究中,隨著水分虧缺的加重,垂穗披堿草種子達到發(fā)芽和出苗階段的時間呈上升的趨勢。這再次證明水分虧缺抑制了垂穗披堿草種子發(fā)芽出苗的速率。
研究顯示,垂穗披堿草種子發(fā)芽的需水閾值為-0.85 MPa,出苗的需水閾值為-0.61 MPa,表明垂穗披堿草種子發(fā)芽和出苗對水分虧缺的耐性不同,其種子出苗過程對水勢的要求比發(fā)芽嚴格。同時,這也表明垂穗披堿草種子發(fā)芽期比出苗期更加耐旱。這與李文嬈等[25]和朱教君等[26]研究報道相一致。
4 結(jié)論
垂穗披堿草在種子發(fā)芽和幼苗形成進程中,水分虧缺降低和推遲了種子群體發(fā)芽出苗速率和群體發(fā)芽出苗的高峰期,且隨著水分虧缺的加重而越發(fā)明顯。復水后,隨著水分虧缺的加重,種子群體發(fā)芽出苗高峰期同樣存在滯后情況。不同濃度PEG-6000溶液造成的水分虧缺抑制了種子的發(fā)芽率、出苗率、發(fā)芽指數(shù)和活力指數(shù),但-0.15 MPa的水分虧缺對種子發(fā)芽率和出苗率影響較小;復水后,-0.50 MPa的水分虧缺對種子發(fā)芽率和出苗率影響較小。-0.15 MPa處理更有利于垂穗披堿草幼苗的生長。-0.75 MPa處理下,垂穗披堿草幼苗生長均受到嚴重抑制。-1.00 MPa處理沒有超出垂穗披堿草幼苗對干旱的適應范圍。垂穗披堿草50%種子群體發(fā)芽的需水閾值為-0.85 MPa,50%種子群體出苗的需水閾值為-0.61 MPa,垂穗披堿草種子發(fā)芽階段比出苗階段更加耐旱。
鑒于野生垂穗披堿草成苗期間具有較強的耐旱性,本研究建議該野生草可作為“三江源”地區(qū)植被恢復的重要參考草種。本研究從生態(tài)生理角度,僅研究了野生垂穗披堿草成苗期間的耐旱性及其需水特性,不足以全面揭示垂穗披堿草在“三江源”地區(qū)半干旱條件下的適應性。因此,今后應繼續(xù)進行該野生草在苗期、拔節(jié)期、孕穗期及花期等階段的耐旱性及需水特性研究,以期實現(xiàn)“三江源”地區(qū)牧草資源的可持續(xù)利用和生態(tài)環(huán)境的可持續(xù)保護。
