內(nèi)生真菌對(duì)高寒草地紫花針茅凋落物分解的影響
2021-09-22 08:56:18宋梅玲王玉琴王宏生鮑根生
草業(yè)學(xué)報(bào) 2021年9期
關(guān)鍵詞:影響
宋梅玲,王玉琴,王宏生,鮑根生
(省部共建三江源生態(tài)與高原農(nóng)牧業(yè)國(guó)家重點(diǎn)實(shí)驗(yàn)室,青海大學(xué)畜牧獸醫(yī)科學(xué)院,青海省高寒草地適應(yīng)性管理重點(diǎn)實(shí)驗(yàn)室,青海西寧810016)
凋落物分解是陸地生態(tài)系統(tǒng)中碳素周轉(zhuǎn)和礦質(zhì)養(yǎng)分循環(huán)的重要過(guò)程[1],并且具有調(diào)節(jié)土壤有機(jī)物組成的作用[2],是釋放礦質(zhì)養(yǎng)分供植物生長(zhǎng)的重要過(guò)程,并由此影響陸地生態(tài)系統(tǒng)凈生產(chǎn)力[3]。凋落物分解率可以被生物和非生物因素所影響,決定分解率的因素主要有3個(gè),分別為氣候因素(如溫度和水分)、凋落物質(zhì)量(如化學(xué)成分)以及分解生物的特性和數(shù)量(如細(xì)菌、真菌和土壤無(wú)脊椎動(dòng)物)[4]。最初的凋落物質(zhì)量(如:C/N、纖維素、木質(zhì)素)可以影響參與凋落物分解的微生物的多度、組成和活性,而這些也反過(guò)來(lái)決定凋落物的分解速率和養(yǎng)分循環(huán)[5]。
近年來(lái),各國(guó)學(xué)者對(duì)于內(nèi)生真菌與其宿主植物(禾草)和食草動(dòng)物之間的相互關(guān)系及其在不同生態(tài)環(huán)境中的多樣性已開(kāi)展了大量的研究[6-9]。內(nèi)生真菌與禾草在生態(tài)系統(tǒng)中維持著一種互利共生的關(guān)系,對(duì)于內(nèi)生真菌來(lái)說(shuō),宿主禾草保證了其在植物體內(nèi)生長(zhǎng)所必需的營(yíng)養(yǎng)物質(zhì)和可靠有效的傳播,而對(duì)于禾草來(lái)說(shuō),內(nèi)生真菌通過(guò)其自身產(chǎn)生的次生代謝物保護(hù)了植物,使其免受外界環(huán)境脅迫的危害[10-11]。這種共生關(guān)系已經(jīng)被證明可以影響生態(tài)系統(tǒng)過(guò)程,如土壤呼吸、凋落物分解等[12-13]。內(nèi)生真菌通過(guò)改變微生物量及微生物群落結(jié)構(gòu),或宿主植物凋落物的數(shù)量和質(zhì)量[14-15]影響凋落物分解速率。
紫花針茅(Stipa purpurea)草原是青藏高原地區(qū)高寒草地中分布面積較廣、最具有代表性的草地類型。其建群種紫花針茅系寒旱生植物,粗蛋白含量高,粗纖維少,營(yíng)養(yǎng)價(jià)值比較高[16],且具有耐寒、耐旱、耐踐踏和抗風(fēng)沙等特性[17-18]。主要分布在海拔1900~5150 m的高原地帶,是青藏高原、帕米爾高原和中亞高山地帶的特有植物[19]。紫花針茅內(nèi)生真菌主要為無(wú)性繁殖的Epichlo?屬內(nèi)生真菌,近年來(lái),有關(guān)紫花針茅與內(nèi)生真菌之間相互關(guān)系的研究較多,如紫花針茅內(nèi)生真菌分離和鑒定,紫花針茅內(nèi)生真菌共生體對(duì)根寄生危害的響應(yīng)等[19-20]。然而,關(guān)于內(nèi)生真菌對(duì)紫花針茅凋落物分解的影響及其機(jī)制的研究尚未見(jiàn)報(bào)道。
本研究通過(guò)對(duì)比帶內(nèi)生真菌與不帶內(nèi)生真菌紫花針茅之間的凋落物分解特征差異,明確內(nèi)生真菌對(duì)宿主植物凋落物分解過(guò)程的影響(包括凋落物重量和質(zhì)量等變化),以期揭示禾草內(nèi)生真菌對(duì)草地生態(tài)系統(tǒng)元素循環(huán)中凋落物分解的作用,為禾草內(nèi)生真菌共生體在高寒草地的利用提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 凋落物采集
2017年9月,于青海省海北州海晏縣(N 37°04′01″,E 100°52′48″)天然草場(chǎng)采集紫花針茅花序,每單株裝于一個(gè)信封袋。將采自野外的紫花針茅花序分別按照李春杰等[21-22]的方法進(jìn)行內(nèi)生真菌檢測(cè),采集帶菌植株的種子并通過(guò)高溫高濕法進(jìn)行去除內(nèi)生真菌的處理,建立帶內(nèi)生真菌(E+)與不帶內(nèi)生真菌(E-)無(wú)性繁殖系種群。將處理好的E+、E-種子分別種植于花盆中,置于溫室,使其自然生長(zhǎng)。待植株長(zhǎng)到枯黃時(shí),采集地上植株,自然風(fēng)干,作為凋落物備用。
1.2 試驗(yàn)設(shè)計(jì)
2018年12月,將E+、E-紫花針茅植株凋落物分別剪成5 cm的小段,稱取5.00 g裝入1 mm孔徑的尼龍網(wǎng)袋(大小為:10 cm×10 cm),共計(jì)36個(gè)網(wǎng)袋,放置到海北州海晏縣試驗(yàn)點(diǎn),每個(gè)網(wǎng)袋間距大于10 cm,放置前清除地上植物組織,確保網(wǎng)袋均勻平鋪在地表與土壤直接接觸,并用大頭針固定。在放置網(wǎng)袋后的第0、3、6、8、10、12個(gè)月隨機(jī)采集凋落物網(wǎng)袋,每次每個(gè)處理采集3個(gè)網(wǎng)袋,以備后續(xù)試驗(yàn)用。
1.3 分析方法
網(wǎng)袋取出后,將凋落物全部取出,放置到實(shí)驗(yàn)室自然風(fēng)干,之后過(guò)0.25 mm篩后稱重。根據(jù)Olson[23]的指數(shù)分解模型公式,計(jì)算凋落物分解速率k值:

式中:X表示經(jīng)過(guò)t年分解時(shí)間后凋落物的殘留量;X0為凋落物初始量(5 g)。根據(jù)此公式,可模擬出凋落物殘留率隨時(shí)間變化的指數(shù)回歸方程,進(jìn)而推算出不同凋落物分解50%即半分解時(shí)間和分解95%所需的時(shí)間。
每次采集的網(wǎng)袋中的樣品都需要測(cè)定其全氮、木質(zhì)素和纖維素含量。其方法為:全N含量運(yùn)用全自動(dòng)凱氏定氮儀(Kjeltec 2300 Auto SystemⅡ,F(xiàn)oss Tecator AB,瑞典)進(jìn)行測(cè)定。木質(zhì)素和纖維素的含量則依據(jù)酸性洗滌纖維法進(jìn)行測(cè)定[24]。同時(shí),計(jì)算每個(gè)分解階段的凋落物木質(zhì)素/N以及N殘留率、木質(zhì)素殘留率和纖維素殘留率等。
元素殘留率(R):
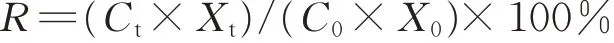
式中:Ct為t時(shí)刻凋落物元素含量(mg·g-1);Xt為t時(shí)刻凋落物干重(g);C0為初始元素含量(mg·g-1);X0為初始干重(g)。
1.4 數(shù)據(jù)分析
數(shù)據(jù)運(yùn)用SPSS 17.0(SPSS,Inc.,Chicago,IL)軟件進(jìn)行分析,E+與E-之間凋落物重量、全N、木質(zhì)素和纖維素含量、木質(zhì)素∶N、纖維素∶N以及全N、木質(zhì)素和纖維素的殘留率等的差異顯著性采用獨(dú)立樣本T檢驗(yàn)的方法進(jìn)行檢驗(yàn),內(nèi)生真菌對(duì)紫花針茅凋落物重量、全N、木質(zhì)素和纖維素含量、木質(zhì)素∶N、纖維素∶N以及全N、木質(zhì)素和纖維素的殘留率等不同時(shí)間的影響采用重復(fù)度量方差分析方法進(jìn)行檢驗(yàn),顯著性區(qū)間定義為95%水平。
2 結(jié)果與分析
隨著時(shí)間的延長(zhǎng),E+與E-紫花針茅凋落物重量都大幅度下降,其中0~8個(gè)月,凋落物重量下降速度較快,而8~12個(gè)月,下降速度趨于平緩(圖1)。相對(duì)而言,E+紫花針茅的凋落物重量下降速度比E-快,且自第6個(gè)月以后,E+植株的凋落物重量顯著低于E-(P<0.05,圖1)。時(shí)間和內(nèi)生真菌均對(duì)紫花針茅凋落物重量產(chǎn)生了顯著影響(P<0.001),二者之間無(wú)顯著交互作用(P>0.05,表1)。

圖1 E+和E-紫花針茅凋落物重量隨時(shí)間的變化Fig.1 The change of E+and E-S.purpurea litters weight over time
E+與E-紫花針茅凋落物分解系數(shù)分別為0.793和0.616(表2)。據(jù)此推算出,E+紫花針茅凋落物的半分解時(shí)間為0.84年,分解95%的時(shí)間為3.75年。而E-紫花針茅凋落物的半分解時(shí)間為1.14年,分解95%的時(shí)間為4.87年(表2)。因此,內(nèi)生真菌縮短了紫花針茅植株凋落物的分解時(shí)間。

表2 E+和E-紫花針茅凋落物分解特征參數(shù)Table 2 Decomposition characteristics parameters of E+and E-S.purpurea litters
E+和E-紫花針茅凋落物中全氮含量隨時(shí)間的延長(zhǎng)有增加的趨勢(shì),其中,第6、10和12個(gè)月,E+紫花針茅凋落物中全氮含量均顯著高于E-(P<0.05,圖2a),內(nèi)生真菌和時(shí)間對(duì)凋落物全氮含量產(chǎn)生了顯著影響,且二者之間有顯著的交互作用(P<0.05,表1)。另一方面,E+和E-紫花針茅凋落物中木質(zhì)素含量均隨著時(shí)間的延長(zhǎng)迅速降低(圖2b),至第8個(gè)月時(shí),E+和E-凋落物中木質(zhì)素含量均降至初始含量的1.5%以下,且隨著時(shí)間的延長(zhǎng),E+凋落物的木質(zhì)素含量由初始的顯著高于E-(P<0.05),逐漸變?yōu)榕cE-無(wú)顯著差異(P>0.05,圖2b);時(shí)間對(duì)紫花針茅凋落物的木質(zhì)素含量有顯著影響(P<0.001),內(nèi)生真菌與時(shí)間對(duì)木質(zhì)素含量無(wú)顯著的交互作用(P>0.05,表1)。E+和E-紫花針茅凋落物中纖維素含量與木質(zhì)素含量有相同的變化趨勢(shì),均隨著分解時(shí)間的延長(zhǎng)而逐漸降低(圖2c),不同的是,E+凋落物中纖維素含量始終低于E-,并在第3、8、10、12個(gè)月時(shí)達(dá)到顯著水平(P<0.05,圖2c);內(nèi)生真菌和時(shí)間對(duì)紫花針茅凋落物纖維素含量的影響顯著,但二者之間沒(méi)有顯著的交互作用(表1)。

表1 內(nèi)生真菌在不同的時(shí)間對(duì)紫花針茅凋落物重量以及全氮、木質(zhì)素和纖維素含量的重復(fù)度量方差分析Table 1 Repeated measures ANOVA for the effects of endophyte at different times on litter weight,total N,lignin and cellulose contents of S.purpurea litters

圖2 E+和E-紫花針茅凋落物中全氮、木質(zhì)素和纖維素含量隨時(shí)間的變化Fig.2 The change of total N,lignin and cellulose contents in E+and E-S.purpurea litters over time
E+和E-紫花針茅凋落物中木質(zhì)素∶氮值均隨著時(shí)間的延長(zhǎng)而逐漸降低(圖3a)。E+凋落物中木質(zhì)素∶氮值始終高于E-,并在分解第3和6個(gè)月時(shí)達(dá)到顯著水平(P<0.05),在此過(guò)程中,內(nèi)生真菌對(duì)紫花針茅凋落物中木質(zhì)素∶氮值未造成顯著影響,但是時(shí)間對(duì)其比值影響顯著,二者無(wú)明顯的交互作用(表3)。另一方面,E+和E-紫花針茅凋落物中纖維素∶氮值也隨著時(shí)間的延長(zhǎng)而降低,但是E+凋落物中纖維素∶氮值始終低于E-,并在第3、6、10、12個(gè)月時(shí)差異顯著(P<0.05,圖3b),內(nèi)生真菌和時(shí)間對(duì)紫花針茅凋落物中纖維素∶氮值產(chǎn)生了顯著的影響,但二者之間無(wú)明顯的交互作用(表3)。

圖3 E+和E-紫花針茅凋落物中木質(zhì)素∶氮和纖維素∶氮隨時(shí)間的變化Fig.3 The change of lignin∶N and cellulose∶N ratios in E+and E-S.purpurea litters over time
E+和E-紫花針茅凋落物中氮?dú)埩袈示S著時(shí)間的延長(zhǎng)而逐漸下降(圖4a),相對(duì)E-來(lái)說(shuō),在分解后期E+凋落物氮?dú)埩袈首兓呌谄骄徤踔劣猩叩内厔?shì)。到第8和10個(gè)月時(shí),E+凋落物氮?dú)埩袈曙@著低于E-(P<0.05,圖4a)。E+和E-紫花針茅凋落物中木質(zhì)素殘留率均隨著時(shí)間的延長(zhǎng)而逐漸下降,在此過(guò)程中,E+與E-之間差異不顯著(P>0.05,圖4b)。纖維素與木質(zhì)素殘留率有相同的變化趨勢(shì),即E+和E-凋落物中纖維素殘留率均隨著時(shí)間的延長(zhǎng)而降低,至第8和10個(gè)月時(shí),E+凋落物中纖維素殘留率顯著低于E-(P<0.05,圖4c)。總體而言,內(nèi)生真菌顯著影響紫花針茅凋落物氮和纖維素殘留率,分解時(shí)間對(duì)于氮、木質(zhì)素和纖維素殘留率均產(chǎn)生了顯著影響(P<0.05),但是內(nèi)生真菌與分解時(shí)間對(duì)于三者均未產(chǎn)生明顯的交互作用(P>0.05,表3)。

表3 內(nèi)生真菌在不同的時(shí)間對(duì)紫花針茅凋落物木質(zhì)素∶氮和纖維素∶氮比值以及全氮、木質(zhì)素和纖維素殘留率的重復(fù)度量方差分析Table 3 Repeated measures ANOVA for the effects of endophyte at different times on lignin∶N ratio,cellulose∶N ratio,and total N,lignin and cellulose residual rates of S.purpurea litters

圖4 E+和E-紫花針茅凋落物中全氮、木質(zhì)素和纖維素殘留率隨時(shí)間的變化Fig.4 The change of total N,lignin and cellulose residual rates in E+and E-S.purpurea litters over time
3 討論
內(nèi)生真菌在植物凋落物被分解之初,通過(guò)植物組織的自然衰亡而獲得優(yōu)先于其他腐生真菌的定殖優(yōu)勢(shì)[25],它們可能直接降解凋落物,也可能通過(guò)拮抗、資源競(jìng)爭(zhēng)等方式與后定殖的腐生菌相互作用,從而間接影響凋落物分解速率[26-28]。本研究中,帶內(nèi)生真菌的紫花針茅凋落物具有更快的分解速率,進(jìn)而,E+紫花針茅相比E-具有更短的凋落物分解周期(圖1和表1),這可能與以上因素有關(guān)。凋落物分解速率對(duì)養(yǎng)分轉(zhuǎn)化的速度產(chǎn)生直接影響,因此,內(nèi)生真菌的存在加快了紫花針茅凋落物的分解,這有利于短期內(nèi)對(duì)草原生態(tài)系統(tǒng)養(yǎng)分的補(bǔ)充[29-30]。內(nèi)生真菌對(duì)一些禾草凋落物分解的影響已經(jīng)被證實(shí)。Bell-Dereske等[31]發(fā)現(xiàn)內(nèi)生真菌的侵染提高了美國(guó)沙茅草(Ammaophila breviligulata)的凋落物分解速率,本研究結(jié)果與之一致。然而,早期一些研究發(fā)現(xiàn)內(nèi)生真菌具有降低禾草分解速率的可能,Omacini等[14]通過(guò)研究多花黑麥草(Lolium multiflorum)中的內(nèi)生真菌對(duì)凋落物分解的影響,發(fā)現(xiàn)凋落物分解速率減少了17%;Lemons等[32]的研究也表明,與對(duì)照相比,E+處理的凋落物分解速率降低了8%。這可能主要與宿主植物的種類和共生體的生存環(huán)境等有關(guān)。Siegrist等[33]發(fā)現(xiàn),盡管帶菌高羊茅(Schedonorus arundinaceus)的凋落物中具有較高的生物堿含量,也未能造成帶菌與不帶菌高羊茅凋落物的干物質(zhì)損失之間的較大差異,其可能原因是高羊茅的枯黃組織中生物堿的濃度非常低,以至于不能影響到微生物分解者。Gundel等[34]的研究也表明,在初始階段凋落物體內(nèi)生物堿含量越高,則其分解速率越大,但是長(zhǎng)期觀察的結(jié)果顯示凋落物分解速率與生物堿含量之間沒(méi)有相關(guān)性。因此,生物堿可能沒(méi)有直接影響凋落物分解,但是可能參與到了初始微生物侵染凋落物的過(guò)程當(dāng)中,進(jìn)而影響凋落物分解速率。在與其宿主植物共生的過(guò)程當(dāng)中,內(nèi)生真菌也可能通過(guò)產(chǎn)生一些罕見(jiàn)的化合物而比生物堿更加直接地調(diào)控這種反應(yīng)[33-34]。
凋落物質(zhì)量與其分解速率密切相關(guān),是影響其分解的內(nèi)在因素,凋落物質(zhì)量通常以凋落物養(yǎng)分含量的高低來(lái)衡量,并以各種含碳化合物(木質(zhì)素、纖維素、單寧等)與養(yǎng)分元素(氮、磷、鉀等)的含量比值來(lái)表示,或者直接以養(yǎng)分含量表示[35-36]。在凋落物分解過(guò)程中,水溶性物質(zhì)和容易分解的碳水化合物可快速降解,初期呈現(xiàn)較快的分解速率,而木質(zhì)素等難分解物質(zhì)不斷累積,結(jié)合成更加不易分解的高分子化合物,使得凋落物分解速率明顯降低[37],許多研究發(fā)現(xiàn),木質(zhì)素在凋落物分解后期呈現(xiàn)凈積累的現(xiàn)象[38-39]。本研究中,紫花針茅凋落物木質(zhì)素含量隨著分解時(shí)間的延長(zhǎng)而逐漸降低,并且木質(zhì)素∶N以及木質(zhì)素殘留率也持續(xù)降低(圖2b,圖3a和圖4b),一方面可能是因?yàn)橄鄬?duì)木本植物而言,草本凋落物木質(zhì)素含量較低[40],另一方面,本研究中凋落物分解時(shí)間僅持續(xù)一年,尚未進(jìn)行到后期階段。凋落物質(zhì)量損失率小于80%時(shí)認(rèn)為分解處于前期階段,而大于80%處于后期階段[41-42],本研究中E+和E-紫花針茅凋落物的質(zhì)量損失率分別為51.8%和43.8%,并沒(méi)有到達(dá)木質(zhì)素調(diào)控分解速率的后期階段,這可能是木質(zhì)素含量持續(xù)降低的原因之一。在分解初始階段,E+紫花針茅凋落物的木質(zhì)素含量顯著高于E-,而隨著分解時(shí)間的延長(zhǎng),二者之間差距逐漸減小,直至沒(méi)有顯著差異(圖2b),并且E+與E-紫花針茅凋落物的木質(zhì)素殘留率沒(méi)有顯著差異(圖4b),表明E+紫花針茅凋落物中的木質(zhì)素相比E-而言其降解速率更快,內(nèi)生真菌的調(diào)節(jié)作用在凋落物分解中可能發(fā)揮了顯著作用。隨著分解過(guò)程的延長(zhǎng),E+和E-紫花針茅凋落物的纖維素含量均逐漸降低,同時(shí)纖維素∶N以及纖維素殘留率也持續(xù)降低(圖2c,圖3b和圖4c),可見(jiàn),在紫花針茅凋落物分解過(guò)程中,纖維素也隨之逐漸被降解,這與之前的研究結(jié)果一致[39,43]。凋落物C質(zhì)量被認(rèn)為是決定分解的首要因子,纖維素、半纖維素作為凋落物C主要的承載者,一般認(rèn)為在分解過(guò)程中,其含量也逐漸降低[39,44]。另一方面,凋落物纖維素主要是由真菌與細(xì)菌分解的,微生物分解者的數(shù)量及活性直接影響著它們的分解速率[45],因此,內(nèi)生真菌可能通過(guò)影響微生物群落進(jìn)而改變紫花針茅凋落物中纖維素的降解速率[14-15]。本研究中,在分解初期,E+與E-紫花針茅纖維素含量沒(méi)有顯著差異,隨著分解時(shí)間的延長(zhǎng),E+紫花針茅的纖維素含量、纖維素∶N以及纖維素殘留率顯著低于E-。這也可能與纖維素酶的活性有關(guān)。許多研究表明,較高的N含量能增加纖維素酶的活性從而促進(jìn)纖維素降解[46-47],本研究發(fā)現(xiàn)E+紫花針茅至分解后期有更高的全N含量(圖2a),這可能部分解釋了后期E+紫花針茅凋落物纖維素的降解速率顯著高于E-的原因。
凋落物分解過(guò)程中,由于養(yǎng)分釋放速度慢于凋落物質(zhì)量損失速率,N、P等養(yǎng)分元素相對(duì)含量一般會(huì)逐漸升高[48]。本研究中,紫花針茅凋落物全氮含量隨著時(shí)間的延長(zhǎng)有上升的趨勢(shì)(圖2a),與前人的研究一致[36]。凋落物在分解過(guò)程中養(yǎng)分元素的遷移模式主要有:淋溶-富集-釋放、富集-釋放和直接釋放3種模式[49-50]。元素的殘留率與釋放率密切相關(guān),所以利用化學(xué)元素殘留率的趨勢(shì)圖可分析元素在分解過(guò)程中的釋放情況,即殘留率下降表示元素在釋放,殘留率上升表示元素在富集[36,49-50]。E+和E-紫花針茅凋落物分解過(guò)程中N元素的殘留率基本呈下降-升高-下降的趨勢(shì),表明紫花針茅的凋落物分解N元素的遷移可能呈淋溶-富集-釋放模式。其中E+紫花針茅凋落物中的N殘留率相比E-而言變化更快,且在后期,E+凋落物的N殘留率顯著低于E-(圖4a),這可能是由于E+紫花針茅分解過(guò)程中微生物活動(dòng)更加強(qiáng)烈,對(duì)N的需求更高,這也部分解釋了E+紫花針茅凋落物相比E-具有更快的分解速度的原因。凋落物分解主要是微生物作用的過(guò)程,它們?cè)诜纸膺^(guò)程中對(duì)N、P等元素濃度有很高的要求,因此,當(dāng)?shù)蚵湮锖@些養(yǎng)分濃度較高時(shí),微生物群落生長(zhǎng)繁殖加速,活動(dòng)加劇,分解速度加快[36,51]。這也可能是E+紫花針茅凋落物在分解過(guò)程中具有更低的纖維素殘留率的原因之一(圖4c),有研究表明,內(nèi)生真菌在強(qiáng)化還田秸稈中有較廣泛的應(yīng)用[52],即植物內(nèi)生真菌有降解纖維素的能力[53],由此可以推測(cè),在本研究中,內(nèi)生真菌也可能通過(guò)促進(jìn)紫花針茅凋落物中纖維素的降解提高凋落物分解速率。然而隨著分解時(shí)間的延長(zhǎng),這種優(yōu)勢(shì)有減弱的趨勢(shì),這可能主要是由于此時(shí)E+與E-紫花針茅凋落物中纖維素的殘留率均到了較低的水平,因此內(nèi)生真菌的作用逐漸不明顯。
4 結(jié)論
內(nèi)生真菌可以增強(qiáng)宿主植物紫花針茅的凋落物分解速率,縮短其分解周期,同時(shí),隨著分解時(shí)間的延長(zhǎng),內(nèi)生真菌不同程度地促進(jìn)了紫花針茅凋落物中N元素、木質(zhì)素和纖維素的降解。因此,內(nèi)生真菌對(duì)于加快紫花針茅向草地生態(tài)系統(tǒng)的養(yǎng)分輸入起到積極作用。然而,本研究只觀測(cè)了短期內(nèi)凋落物分解的過(guò)程,更長(zhǎng)期的凋落物分解變化以及分解環(huán)境條件、微生物活動(dòng)和內(nèi)生真菌次生代謝物等因素的影響仍需要進(jìn)一步研究。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
