小麥旗葉與幼苗性狀的QTL分析
2021-09-23 09:18:56李興茂王淑英倪勝利
麥類作物學報 2021年5期
李興茂,王淑英, 倪勝利
(甘肅省農業科學院旱地農業研究所,甘肅蘭州 730070)
相對于苗期活力低的品種,苗期活力高的品種其籽粒產量平均提高12%,有明顯的節水增產效果[1-2]。高幼苗活力與高生物量的積累相關,在高溫、高CO2濃度等脅迫條件下,籽粒產量也顯著提高[3-4]。干旱環境下根系活力大的品種,在水分脅迫條件下產量的損失最小[5]。苗期活力與萌發期胚芽鞘長度、根系活力及莖葉活力密切相關。胚芽鞘長的品種有利于雨養環境下小麥深層水份的利用,具有耐深播的優點[6]。較高的根系活力,可增強對營養的吸收和生物量的積累,對提高小麥苗期抵抗水分脅迫[7]和后期抵抗干旱脅迫均有重要的影響。
前人在1A、6A、6D和7D染色體上定位到控制胚根長度的QTL,在2D和6D染色體上定位到控制根莖比率的QTL[8-10];在1A、2A、2B、4B、5A、5D、6A 和7B染色體上定位到控制根長的QTL,在5D染色體上定位到控制根、莖干重的QTL,在1D染色體上定位到控制莖稈長度的QTL[9,11-12],然而,很多QTL存在顯著的上位性效應[10,13]。Manschadi等[14]研究發現,幼苗期根生長角度較窄的品種,其成株期同樣具有較窄的根生長角度。Spielmeyer等[15]研究發現,在6A染色體上控制幼苗葉片寬度主效QTL緊密連鎖的分子標記NW3106,也與胚芽鞘長度和株高QTL緊密連鎖。Bennett等[16]研究發現,在3B染色體上控制苗期活力的QTL,也與籽粒產量、葉寬和冠層溫度等性狀有關。
小麥苗期性狀相關的QTL定位報道較多,但苗期性狀及旗葉性狀的關系還不清楚。本研究利用京冬8號/矮抗58的207個RIL群體為材料,針對冬小麥幼苗期性狀和開花期旗葉性狀進行QTL研究,探討小麥苗期性狀與后期旗葉性狀的關系,以期發掘出穩定調控小麥幼苗性狀相關的QTL,為今后分子標記輔助選擇育種提供 依據。
1 材料與方法
1.1 供試材料
試驗材料為京冬8號/矮抗58的207個RIL群體及其親本。其中,京冬8號攜帶矮稈基因Rht8,矮抗58攜帶矮稈基因Rht8和Rht2[17]。
1.2 試驗設計
室內試驗于2016年10月在實驗室進行,將發芽紙在121℃下滅菌30 min,每個材料選取無損傷且大小均勻的種子200粒,用1.0%次氯酸鈉消毒4 min,沖洗干凈后,置于恒溫光照培養箱(25 ℃)培養,重復3次,待發芽7 d后,測定發芽數、苗長、根長、苗鮮重、根鮮重、苗干重、根干重等指標。鮮重根冠比=根鮮重/苗鮮重;干重根冠比=根干重/苗干重。
田間試驗于2015年在甘肅省慶陽市鎮原試驗站進行,種植RIL群體及其親本,行長2 m,重復2次。于2016年5月14日小麥揚花期,測定旗葉長度和寬度,計算旗葉面積和旗葉長寬比。參照Li等[18]和李興茂等[19]的方法測定產量、植被指數、植被覆蓋度、葉片衰老速率、千粒重等 性狀。
1.3 數據分析
用Excel進行數據整理,每個性狀3次重復的平均值為表型值。從1 208對引物中進行雙親多態性篩選,篩選出391對差異引物。依據GrainGenes 2.0圖譜提供的標記位置,利用Try命令進行調整、構建包含149對引物的遺傳圖譜。利用IciMapping 4.1軟件中的雙親群體QTL定位功能,基于逐步回歸的完備復合區間作圖法,以 LOD>2.5作為閾值進行基因定位分析,分析方法參見文獻[18-19]。
2 結果與分析
2.1 RILs群體及親本的苗期與旗葉性狀
從表1可以看出,幼苗期性狀中,苗長、苗干重、根干重、鮮重根冠比和干重根冠比在親本間差異較大,除鮮重根冠比和干重根冠比外,京冬8號的其他6個性狀均小于矮抗58。旗葉性狀中,京冬8號的旗葉長、寬、面積、長寬比4個性狀的表型值均高于矮抗58。群體中苗長、根長、苗鮮重、根鮮重和苗干重的平均值都大于雙親,而旗葉長、寬和面積的群體平均值介于雙親之間。群體內所有研究性狀均表現出超親分離現象。表明這些性狀是由多基因控制的數量性狀,適合進行QTL定位。所有性狀在群體內基因型間存在顯著 差異。
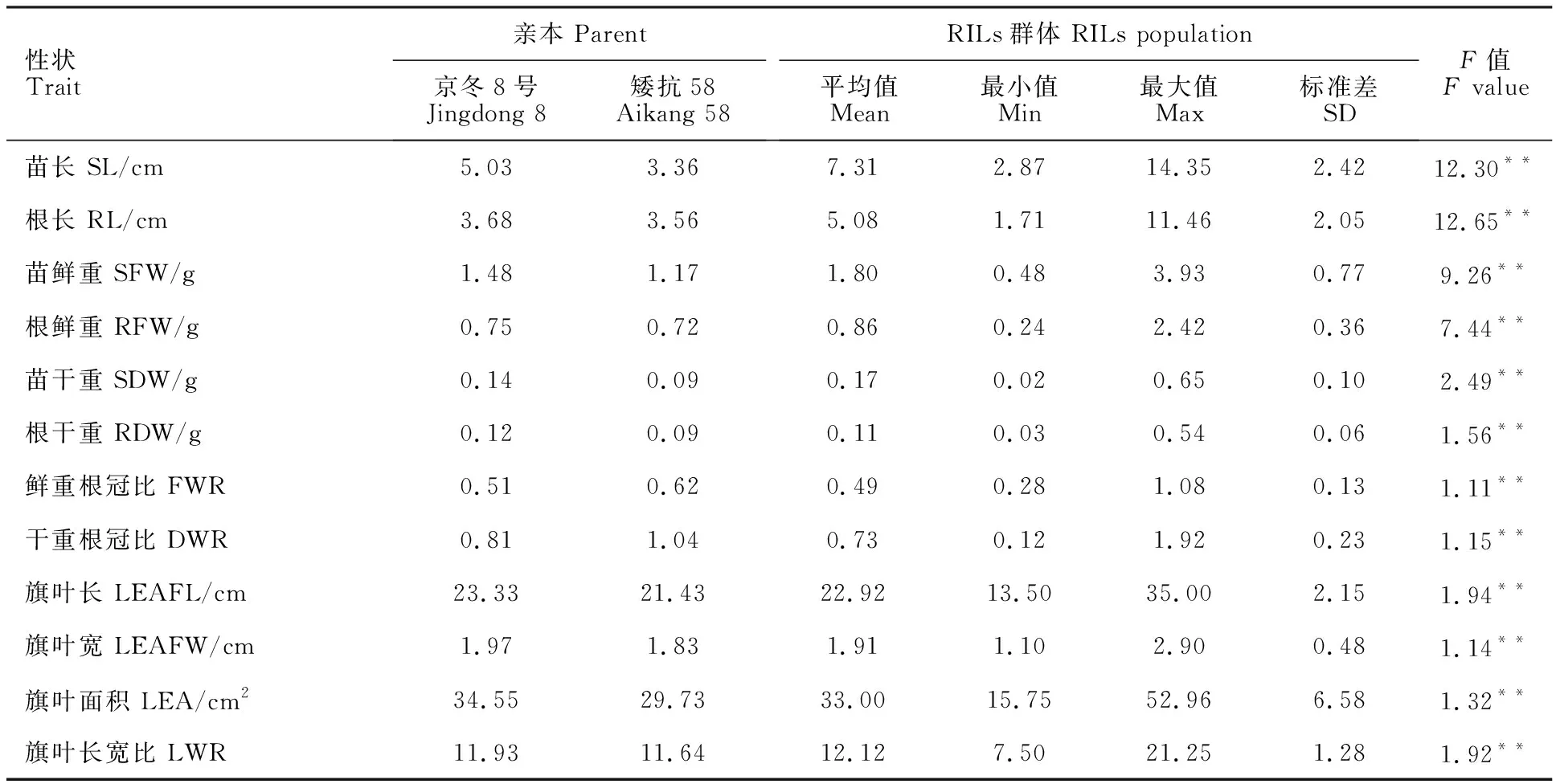
表1 RIL群體及其親本的苗期和旗葉性狀Table 1 Phenotypic performance for the investigated traits of the RILs and their parents
2.2 旗葉相關性狀的QTL定位結果
從表2可以看出,共檢測到10個控制旗葉性狀的QTL,其中4個QTL與旗葉面積有關,分布在1A和5D染色體上,表型貢獻率為1.98%~7.69%;1個QTL與旗葉長有關,分布在1A染色體上,表型貢獻率為9.24%;1個QTL與旗葉寬有關,分布在5D染色體上,表型貢獻率為 6.54%;共檢測到4個控制旗葉長寬比的QTL,均位于D染色體組上,分別分布在2D、3D、4D和5D染色體上,表型貢獻率為2.73%~9.89%,其中,2D和5D染色體上的QTL加性效應來自矮抗58,3D和4D染色體上的QTL來自京冬8號。其中,1A染色體上與旗葉面積QTL緊密連鎖的分子標記wmc550也與旗葉長QTL緊密連鎖,遺傳距離均為0.1 cM,加性效應均來自親本矮抗58;5D染色體上與旗葉面積QTL緊密連鎖的分子標記gwm182也與旗葉寬QTL連鎖,但遺傳距離不同,分別為0.4和2.4 cM,其加性效應均來自親本京冬8號。有6個QTL的表型貢獻率大于5%,為主效QTL,分別為1A染色體上控制旗葉面積和旗葉長的QTL、4D染色體上控制旗葉長寬比的QTL、5D染色體上控制旗葉面積、旗葉寬和旗葉長寬比的QTL。除5D染色體上控制旗葉面積和旗葉寬的QTL表現為部分顯性效應外,與其他性狀相關的QTL均表現為超顯性效應。
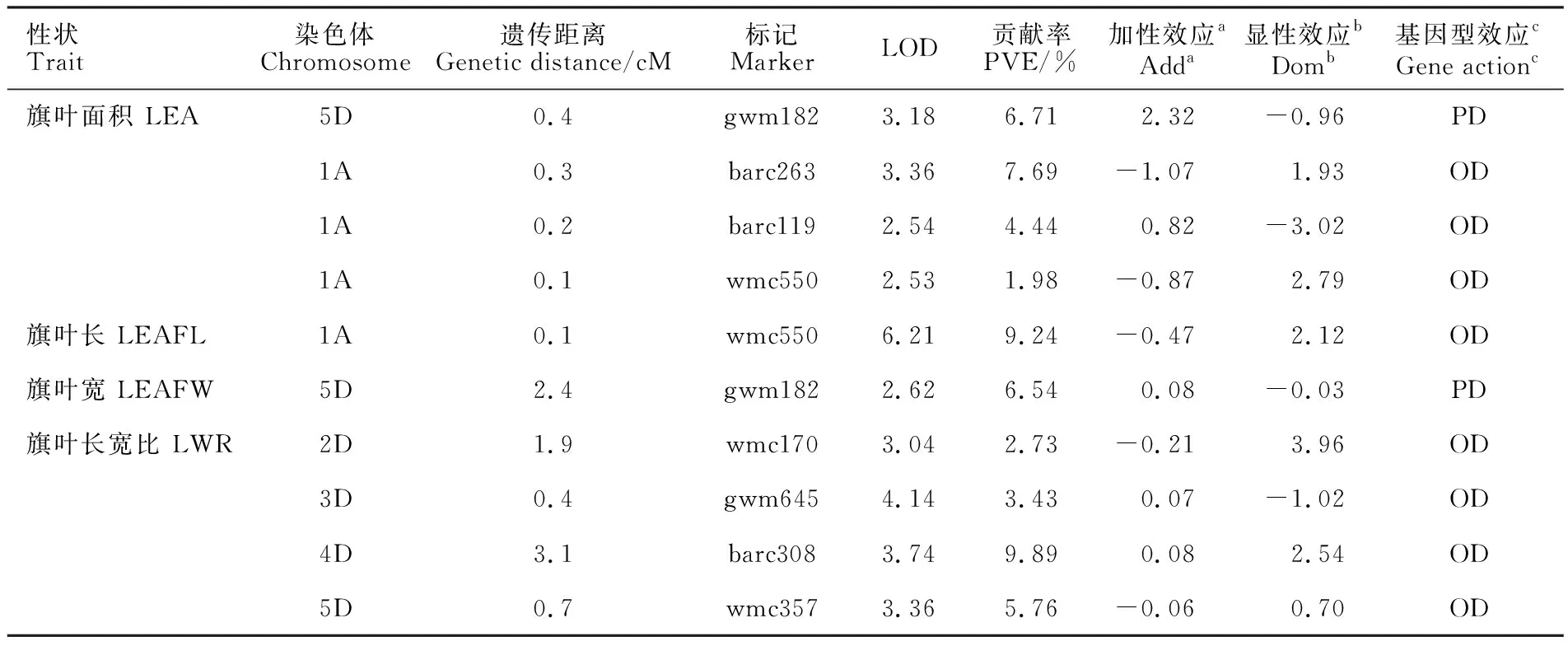
表2 旗葉相關性狀的QTLTable 2 QTLs for flag leaf traits
2.3 幼苗期相關性狀的QTL定位結果
從表3可以看出,共檢測到22個控制幼苗性狀的QTL,其中7個QTL與根長有關,分布在1A、2D、3D、4D(2)、5A和7A染色體上,表型貢獻率為2.53%~4.84%;1個QTL與幼苗根鮮重有關,分布在2D染色體上,表型貢獻率為 1.31%;2個QTL與幼苗根干重有關,分布在4D和7A染色體上,表型貢獻率分別為1.14%和 4.38%;3個QTL與幼苗苗長有關,分布在1A、4A和4D染色體上,表型貢獻率為3.10%~ 10.52%;2個QTL與幼苗鮮重有關,分布在2D和5A染色體上,表型貢獻率分別為4.91%和 1.23%;3個QTL與幼苗干重有關,分布在1A、2D和7A染色體上,表型貢獻率為2.32%~ 3.32%;2個QTL與鮮重根冠比有關,分布在4D和7A染色體上,表型貢獻率分別為1.88%和 1.90%;2個QTL與干重根冠比有關,分布在2D和7A染色體上,表型貢獻率分別為2.78%和 2.74%。1A染色體上與根長QTL緊密連鎖的分子標記wmc84也與苗長和苗干重QTL緊密連鎖,但遺傳距離不同,分別為4.9、3.0和4.0 cM,除根長QTL加性效應來自親本京冬8號外,其他兩個QTL均來自矮抗58。2D染色體上與根長QTL緊密連鎖的分子標記wmc170也與根鮮重、苗鮮重、苗干重和干重根冠比QTL緊密連鎖,遺傳距離分別為5.8、0.9、6.0、0.9和3.0 cM,除干重根冠比QTL的加性效應來自親本矮抗58外,其他性狀均來自親本京冬8號。4D染色體上與根長QTL緊密連鎖的分子標記barc308與鮮重根冠比QTL緊密連鎖,遺傳距離分別為11.0和0.2 cM;與根干重QTL緊密連鎖的分子標記barc98與苗長QTL緊密連鎖。5A染色體上與根長QTL緊密連鎖的分子標記gwm156也與苗鮮重QTL緊密連鎖,遺傳距離分別為3.0和5.0 cM。7A染色體上與根長QTL緊密連鎖的分子標記barc1136也與根干重、苗干重、鮮重根冠比和干重根冠比QTL緊密連鎖,遺傳距離分別為6.0、2.0、7.0、2.0和1.0 cM。Rht2矮稈基因位于4D染色體上,但與4D染色體上控制根長、根干重、苗長和鮮重根冠比的QTL距離均較遠。除1A和4D染色體上控制苗長的QTL表型貢獻率大于5%外,其他與幼苗期相關性狀的QTL表型貢獻率均較低,且除3D染色體上控制根長的QTL表現為部分顯性效應外,其他QTL均表現為超顯性效應。
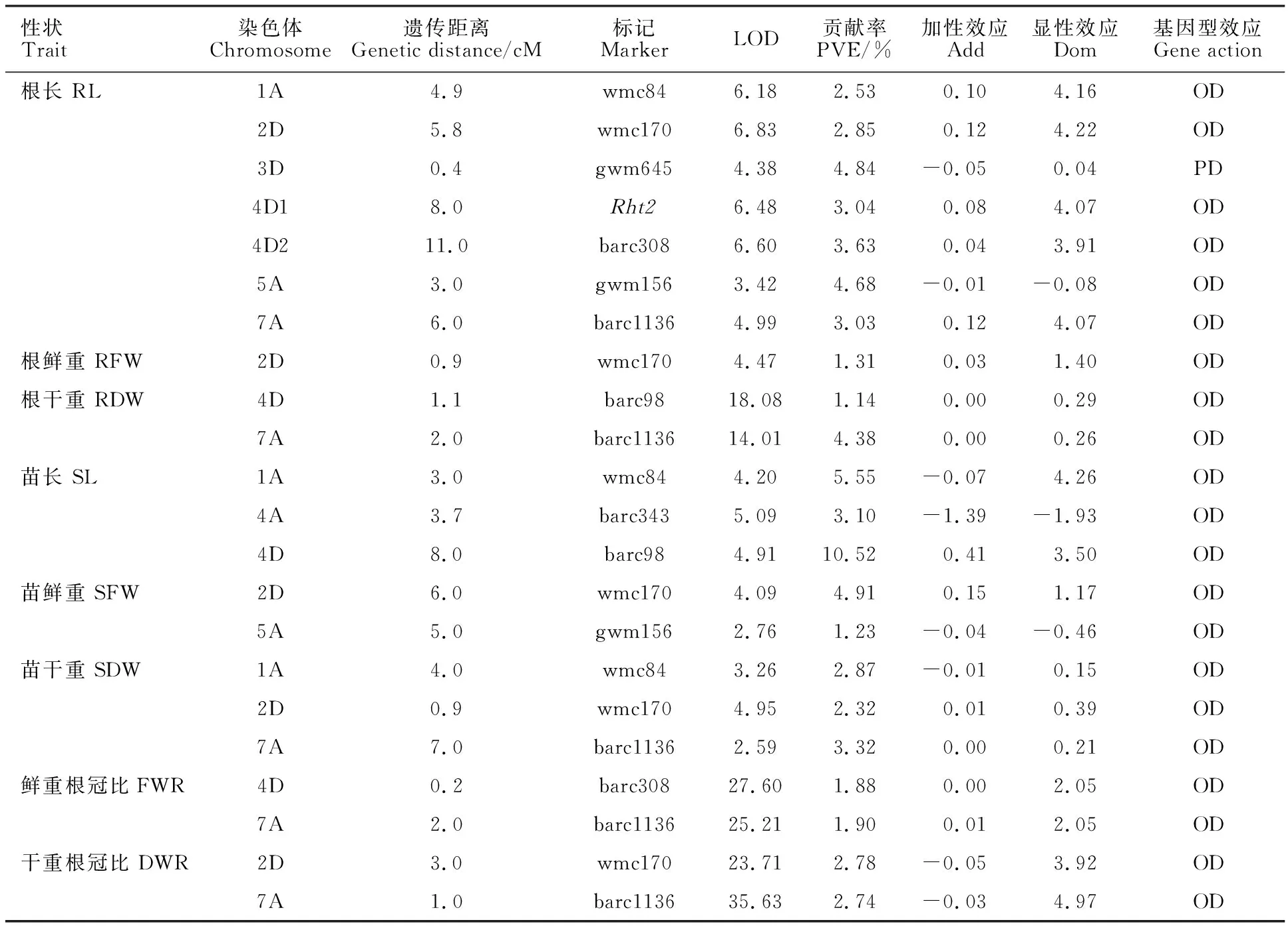
表3 幼苗期相關性狀的QTLTable 3 QTLs for seedling traits
3 討 論
本研究發現,2D染色體上與旗葉寬QTL緊密連鎖的分子標記wmc170與幼苗根長、根鮮重、苗鮮重、苗干重和干重根冠比QTL緊密連鎖;4D染色體上與旗葉長寬比QTL緊密連鎖的分子標記barc308與幼苗根長、鮮重根冠比QTL緊密連鎖,其他旗葉性狀QTL緊密連鎖的分子標記沒有發現與幼苗期性狀QTL緊密連鎖的現象。袁倩倩等[9]在1A染色體側翼標記wmc550附近檢測到控制根長的QTL,而本研究在1A染色體側翼標記wmc550附近檢測到控制旗葉面積和旗葉長的QTL。Li等[20]在1A染色體側翼標記wmc84附近檢測到控制抽穗期、植被歸一化指數和植株衰老速率的QTL,而本研究在1A染色體側翼標記wmc84附近檢測到控制根長、苗長和苗干重的QTL,遺傳距離不同。Landjeva等[21]在5D染色體側翼標記gwm182附近檢測到控制苗長和根莖比率的QTL,而本研究在5D染色體側翼標記gwm182附近檢測到控制旗葉面積和旗葉寬的QTL,但遺傳距離不同。因此,與幼苗期性狀相關的QTL存在與苗期活力、后期生長發育性狀和產量性狀等QTL共同的連鎖分子標記,這些標記可有效用于小麥分子標記輔助育種。盡管本研究檢測到的QTL都是微效QTL,但驗證了這些QTL位點與種子苗期活力性狀有關,對苗期性狀與小麥旗葉性狀的深入研究有參考價值。
本研究在4D染色體上定位到矮稈基因Rht2,朱 浩等[22]研究表明,Rht2基因不僅影響根長,而且與產量和千粒重有關。但本研究檢測到Rht2與控制根長的QTL距離較遠。Tian等[23]在6A染色體上定位到矮稈基因Rht24,可增加春季苗期活力和千粒重,但與幼苗期活力性狀無關。因此,Rht24基因彌補了大多數矮稈基因降低小麥初期生長活力的缺陷,對研究耐旱相關性狀遺傳檢測及分子標記輔助選育奠定了基礎。
本試驗采用室內苗期性狀研究方法,占用空間小,一次可以測試批量樣品,有利于對遺傳群體的多樣本開展多重復研究。然而,統計分析的工作量大,不能及時完成所有品系的根系性狀測量,可能造成品系間測量誤差,因此,本研究在測定時,每個重復間適當增加了試驗間隔,盡可能減少誤差。今后采用數碼拍照后,利用適合于大群體表型研究的根系分析系統軟件進行處理,可以分析更多的幼苗指標,還可進一步減少試驗誤差。
