煙草NtMHX1和NtMHX2基因的克隆及不同金屬離子脅迫下的表達模式分析
2021-09-26 07:30:42張吉順張孝廉楊慧賈蒙驁張婕趙德剛
中國煙草學報 2021年4期
張吉順,張孝廉,楊慧,賈蒙驁,張婕,趙德剛
1 貴州大學生命科學學院/農業生物工程研究院,貴州省農業生物工程重點實驗室,山地植物資源保護與種質創新省部共建教育部重點實驗室,貴陽 550025;
2 貴州省煙草科學研究院,煙草行業分子遺傳重點實驗室,貴陽 550081;
3 貴州省農業科學院農業部DUS中心貴陽分中心,貴陽 550006
鎂是植物生長發育所必需的金屬元素,參與植物體的多個生理代謝過程。煙草(Nicotiana tabacum)是以收獲葉片為主的經濟作物,鎂元素不僅影響烤煙的農藝性狀,還與烤煙品質有著密切的關系[1]。烤煙在缺鎂和高鎂條件下,葉片葉綠素含量均會顯著降低[2]。很多研究都證明了,施鎂可以有效地提升煙葉的產質量并較好地協調煙葉中的化學成分,提升煙葉吸食品質[1,3-5]。
植物中鎂的吸收、運輸和分配受到不同基因家族的調控,目前已發現有4個家族蛋白參與了鎂離子的運輸[6-9],包括:CroA亞家族的部分成員、SV(slow-vacuolar)通道蛋白、環核苷酸相聯的離子通道AtCNGC10、MHX蛋白。其中MHX蛋白屬于CaCA(Ca2+/Cation Antiporter)亞家族,對植物鎂離子平衡具有重要作用。擬南芥AtMHX主要負責質子和Mg2+、Zn2+和Cd2+的交換[10-11],AtMHX蛋白將金屬離子扣留在液泡中,并將液泡中的質子釋放到細胞質中。在橡膠樹中也發現MHX具有交換質子和Mg2+、Zn2+、Cd2+離子的功能[12-13]。
Gaash等[14]對擬南芥(Arabidopsis thaliana)、馬 鈴 薯(Solanum tuberosum)、番 茄(Solanum lycopersicum)、葡萄(Vitis vinifera)、大豆(Glycine max)、 水 稻(Oryza sativa)、 小 麥(Triticum aestivum)、玉米(Zea mays)和小立碗蘚(Physcomitrella patens)等植物中MHX蛋白進行進化和結構模型分析,發現MHX蛋白是植物所特有,且大部分植物都只有一個MHX同源基因。目前,煙草中MHX蛋白的序列信息和功能尚不明確,本文對栽培煙草中的MHX基因進行克隆,對其編碼蛋白進行了生物信息學分析,并檢測了不同金屬離子處理下該基因的表達模式變化,為MHX基因在煙草中的作用及分子機制研究提供了參考。
1 材料與方法
1.1 材料
試驗所用材料為栽培煙草(Nicotiana tabacum L.)K326。采用漂浮育苗法育苗,溫室培養至5~6片真葉時將幼苗移栽至含有基質的花盆中培養,分別取幼苗期的根、莖、葉和開花期花,液氮迅速冷凍,于-80℃保存備用。將5~6片真葉期的煙苗移栽至1/4濃度的Hogland營養液中培養14 d,每7 d更換一次新的營養液,分別在1/4濃度的Hogland營養液中添加100 mmol/L的NaCl、100 mmol/L的KCl、100 mmol/L的CaCl2、50 mmol/L的ZnSO4,50 mmol/L的MnCl2,50 mmol/L的MgSO4、50 mmol/L的CuSO4和50 μmol/L的CdCl2進行不同金屬離子的脅迫處理,并于添加金屬離子的0 d、1 d、2 d、4 d和6 d取煙草葉片,以不額外添加金屬離子的1/4濃度的Hogland營養液中培養的煙苗做為平行對照,同步取樣,液氮速凍后置于-80℃冰箱保存備用,每個處理設置3次重復。
1.2 NtMHX基因的克隆及測序
以擬南芥MHX基因全長CDS序列(登錄號:AT2G47600)為信息探針,利用NCBI煙草數據庫比對,獲得煙草MHX基因同源序列,選擇一致性最高的序列,利用Primer Primer 5.0設計CDS全長克隆引物MHX-F和MHX-R(表1),分別以煙草‘K326’的cDNA為模板,使用NEB公司的KOD Fx NEO高保真酶進行煙草MHX基因的CDS序列的PCR擴增。利用天根膠回收試劑盒對PCR產物進行膠回收后,連入pGEM-T載體,轉化大腸桿菌DH5α,挑取陽性克隆送去上海生工公司測序。
1.3 生物信息學分析
在NCBI網 站(https://www.ncbi.nlm.nih.gov/)上BLASTp進行蛋白預測,利用Specialized BLAST (CDD search)進行保守域分析。選擇其他物種中相似性較高的MHX序列16條用MEGA5.0構建系統進化樹,采用鄰接法(Neighor Joining Method)作圖,重復次數為1000。在蛋白生物信息學網站(http://www.expasy.org/)對NtMHX蛋白進行理化性質、親水性/疏水性、跨膜結構、二級結構及三級結構進行預測和分析。
1.4 NtMHX基因的表達分析
根據NtMHX1和NtMHX2的CDS序列設計熒光定量特異引物2對(表1),同時選用煙草Actin基因(NTU60495)為內參基因,并設計定量PCR引物Ref-RT-F和Ref-RT-R (表1)。采用SYBR Green法在ABI Vii7實時熒光定量PCR儀上進行Real Time-PCR實驗,設置3次生物學重復和3次實驗重復,PCR反應體系和反應程序參照TaKaRa定量PCR試劑盒說明書。對溶解曲線、擴增效率和相關系數進行分析,選取特異性好、擴增效率接近1的引物進行實驗,分析NtMHX1和NtMHX2基因在煙草組織及不同金屬離子處理后各時間點的表達情況。將根中和金屬離子處理0 d的葉片的相對表達量分別設置為1,其余組織中相對表達量按照2-??CT法進行計算。
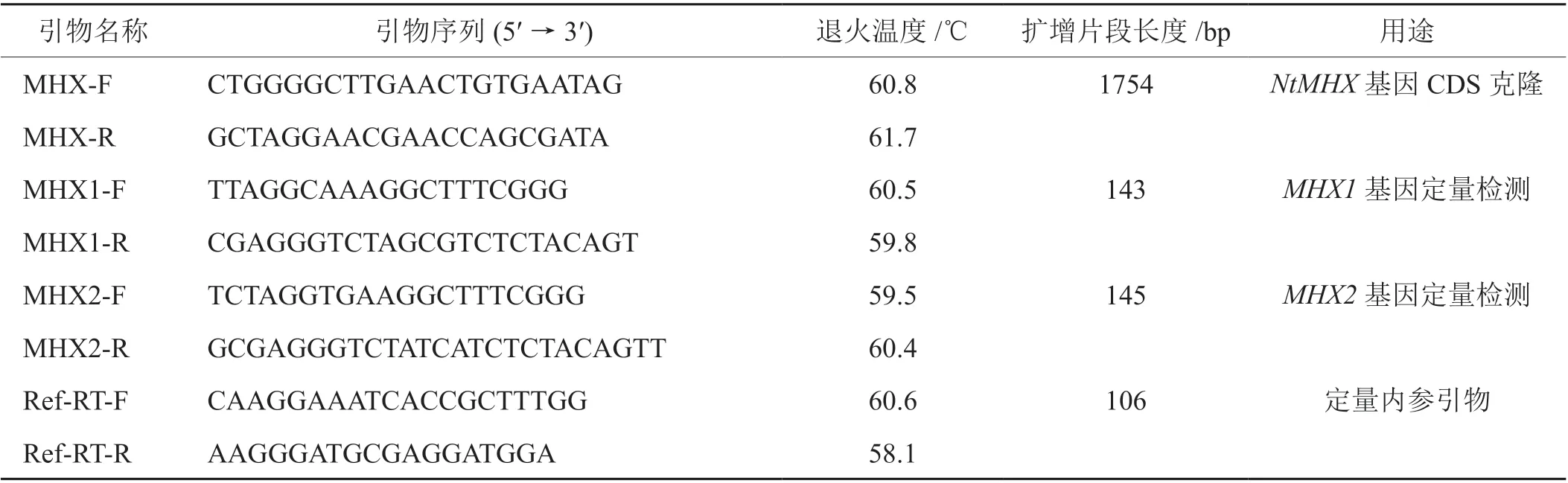
表1 本文中使用引物的序列信息Tab. 1 Sequence information of primers used in this research
2 結果分析
2.1 NtMHX基因克隆
以栽培煙草K326葉片總cDNA為模板,MHX-F和MHX-R為引物,擴增獲得1800 bp左右的片段(圖1)。經測序分析,共獲得2個NtMHX基因CDS序列,一條1623 bp,命名為NtMHX1;一條1641 bp,命名為NtMHX2,序列一致性為96.4%,并擴增到了NtMHX2的一個選擇性剪切體,長度為1620 bp,缺失了21 bp(844-864 bp)。選擇NtMHX1和1641 bp的NtMHX2序列編碼的蛋白進行后續的生物信息學分析和進化分析。
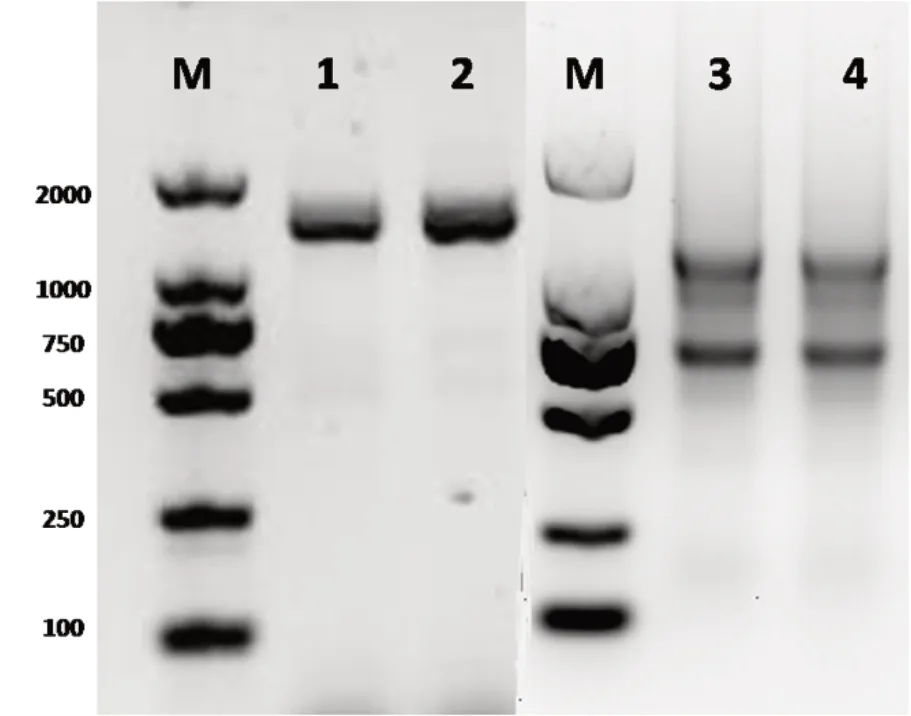
圖1 煙草葉片RNA和NtMHX基因PCR產物電泳圖Fig.1 Electrophoretogram of PCR products of RNA and NtMHXgenes in tobacco leaves
2.2 NtMHX蛋白的生物信息學分析
2.2.1 NtMHX蛋白質理化性質及親/疏水性
利用在線軟件ExPASy-ProtParam對NtMHX1蛋白物理化學性質進行分析,結果顯示,該蛋白質分子式為C2822H4291N689O758S12,由540個氨基酸組成,相對分子量為60382.99 Da,理論等電量(pI)為5.66,屬于酸性蛋白質,由20種氨基酸組成,其中Leu含量最多,占總數的11.7%,而Met的含量最少,占總數的0.7%;該蛋白為穩定蛋白,不穩定系數為38.07(<40),脂肪族氨基酸指數為111.59,總親水性平均值為0.408。
NtMHX2蛋白物理化學性質分析結果顯示,該蛋白質分子式為C2850H4333N693O766S13,由546個氨基酸組成,相對分子量為60977.72 Da,理論等電量(pI)為5.63,為酸性蛋白質。由20種氨基酸組成,其中Leu含量最多,占總數的11.2%,而Met的含量最少,占總數的0.9%。該蛋白為穩定蛋白,不穩定系數為33.16(<40),脂肪族氨基酸指數為111.25,總親水性平均值為0.414。
利用在線軟件ProtScale進一步對蛋白質的親水疏水性進行分析,結果顯示,組成NtMHX1蛋白的氨基酸大部分是疏水性氨基酸,其中,第527位分值最高,為3.222,第235位分值最低,為-2.633,說明該蛋白為疏水性蛋白。組成NtMHX2蛋白的氨基酸大部分是疏水性氨基酸,其中,第533位分值最高,為3.222,第235位分值最低,為-2.367,說明該蛋白為疏水性蛋白。
2.2.2 分析NtMHX蛋白跨膜域及亞細胞定位
利用在線軟件TMHMM 2.0分析預測NtMHX蛋白跨膜區域,結果(圖2)顯示,NtMHX1和NtMHX2蛋白均有11個跨膜區域,是跨膜蛋白。
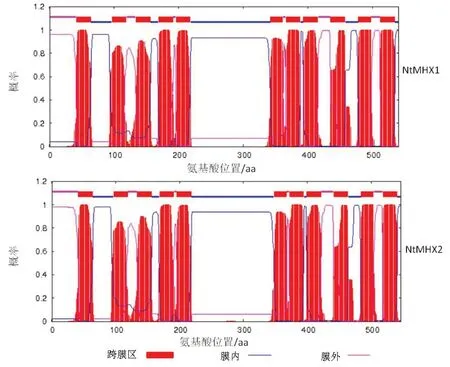
圖2 NtMHX1和NtMHX2蛋白的跨膜結構域預測分析Fig.2 Prediction of the transmembrane domains of NtMHX1 and NtMHX2 proteins
利用在線軟件SignalP 4.0分析NtMHX信號肽分布,結果顯示,NtMHX1和NtMHX2蛋白均沒有信號肽,為非分泌蛋白。
使用Plant-mPLoc軟件預測NtMHX蛋白的亞細胞定位,推測NtMHX1和NtMHX2蛋白定位于細胞膜上。
2.2.3 分析NtMHX蛋白磷酸化位點
通過在線軟件NetPhos 3.1預測NtMHX蛋白氨基酸上的磷酸化位點(圖3)發現,NtMHX1蛋白氨基酸上共有45個磷酸化位點,其中包括26個Ser磷酸位點、12個Thr磷酸位點和7個Tyr磷酸位點。NtMHX2蛋白氨基酸上共有40個磷酸化位點,其中包括22個Ser磷酸位點、10個Thr磷酸位點和8個Tyr磷酸位點。
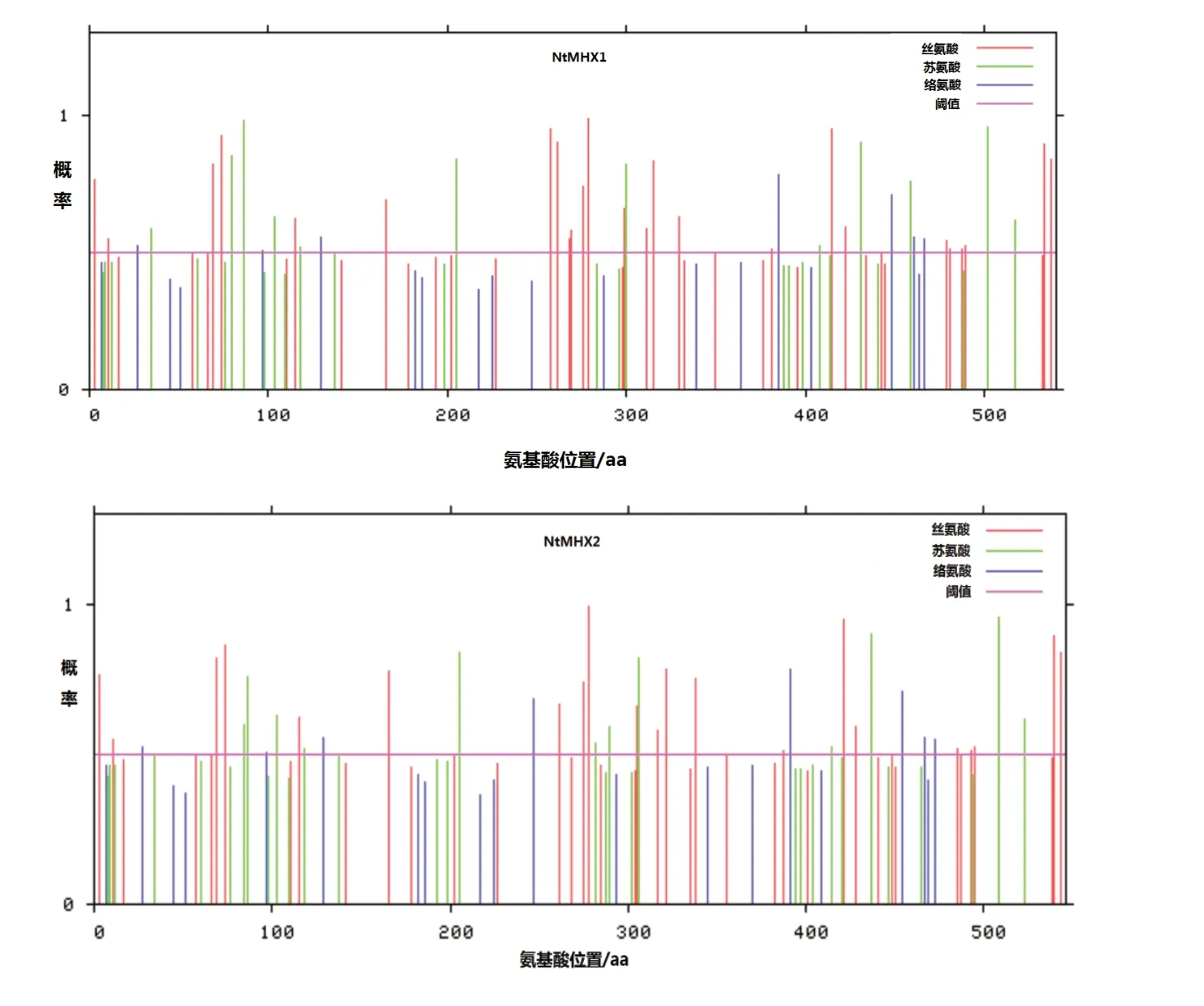
圖3 NtMHX1和NtMHX2蛋白的磷酸化位點預測Fig.3 Prediction of phosphorylation sites in NtMHX1 and NtMHX2 protein sequences
利用在線軟件NCBI-CD Search對NtMHX蛋白保守結構域進行預測(圖4),NtMHX1和NtMHX2均存在兩個CACa蛋白家族的保守結構域。
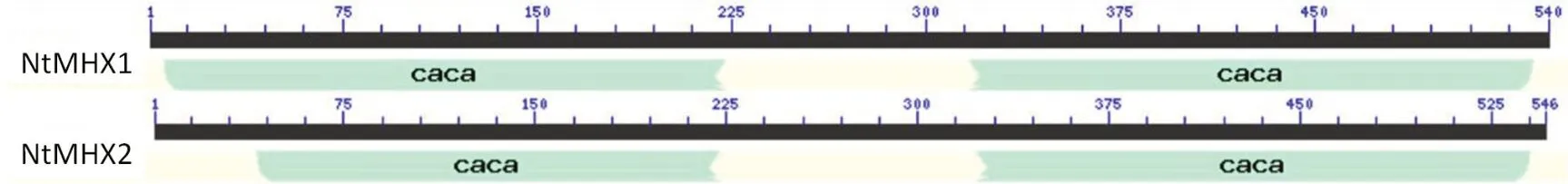
圖4 煙草MHX蛋白保守結構域預測Fig.4 Prediction of conserved domains of MHX proteins
2.2.4 分析MHX蛋白高級結構
通過在線軟件SOPMA對NtMHX蛋白二級結構進行預測,結果(圖5)顯示,NtMHX1蛋白的α螺旋占45.37%,延伸鏈占15.19%,β轉角占5.74%,無規則卷曲占33.70%。NtMHX2蛋白的α螺旋占44.14%,延伸鏈占16.48%,β轉角占5.13%,無規則卷曲占34.25%。使用SWISS-MODEL在線軟件對NtMHX蛋白的三級結構進行預測,結果如圖6,與二級結構預測結果一致,這兩個蛋白富含α螺旋和無規則卷曲。
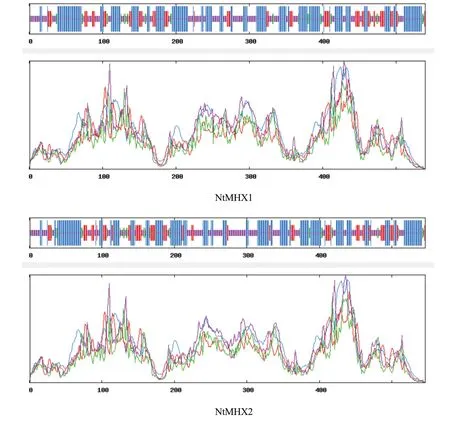
圖5 NtMHX1和NtMHX2蛋白二級結構預測Fig.5 Prediction for secondary structure of NtMHX1 and NtMHX2 proteins
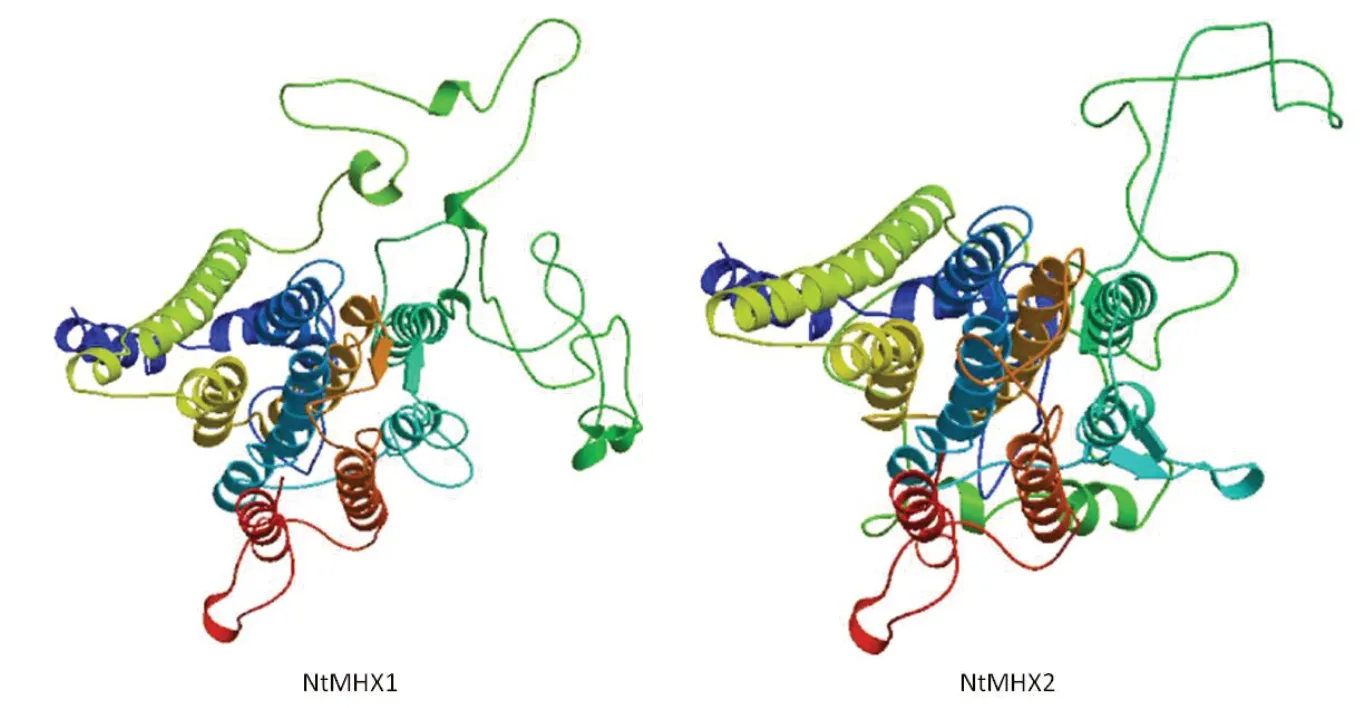
圖6 NtMHX1和NtMHX2蛋白三級結構預測Fig.6 Prediction for thridary structure of NtMHX1 and NtMHX2 proteins
2.3 NtMHX蛋白序列比對及進化分析
在NCBI數據庫中使用Protein Blast程序對NtMHX1和NtMHX2蛋白進行保守區域檢索,發現該蛋白包含2個CaCA結構域,屬于CaCA亞家族。經過蛋白同源檢索,發現來自不同植物物種的MHX蛋白有很高的相似性。NtMHX1與煙草野生種絨毛狀煙草(Nicotiana tomentosiformis)的一致性為99.81%,與林煙草(Nicotiana sylvestris)的一致性為96.67%,NtMHX2與林煙草的一致性為99.82%,與絨毛狀煙草的一致性為96.34%,推測NtMHX1來源于絨毛狀煙草,而NtMHX2來源于林煙草。
選擇部分序列相似性較高、功能注釋明確或有文獻報道的MHX蛋白序列,利用Clustral X進行序列比對(圖7),發現這兩個蛋白具有保守的11個跨膜結構域。煙草NtMHX1和NtMHX2蛋白與茄科植物番茄、馬鈴薯具有100%的序列覆蓋度,與野生番茄序列一致性最高,分別為92.78%和91.77%,與馬鈴薯MHX蛋白序列一致性分別為92.59%和91.77%。用MEGA5.0進行系統進化樹的繪制(圖8),結果可以看出,煙草和同屬茄科植物的馬鈴薯、番茄MHX序列處于同一分枝上。序列比對和聚類分析結果均表明煙草NtMHX1、NtMHX2蛋白與番茄和馬鈴薯的親緣關系最近。
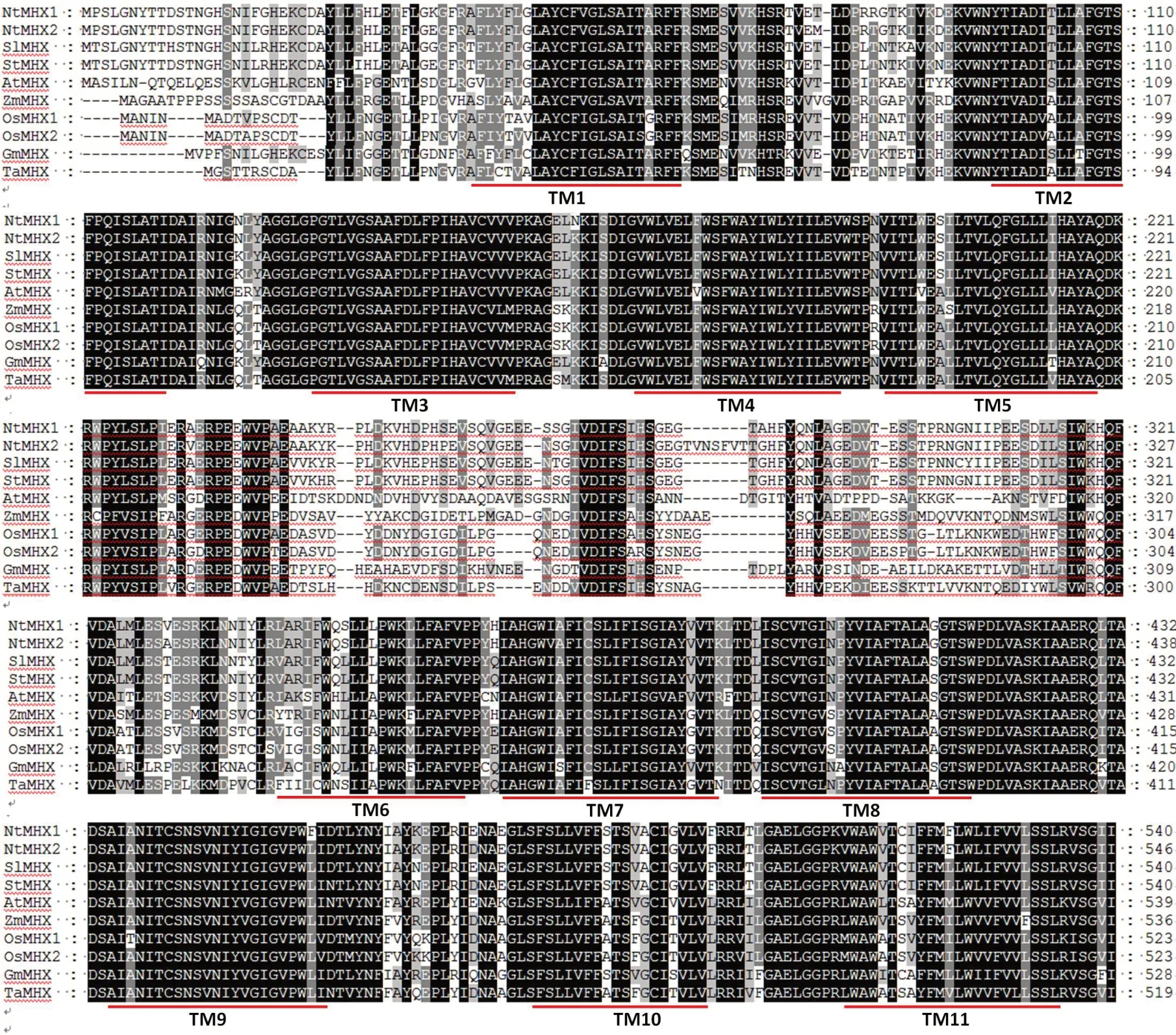
圖7 不同植物MHX蛋白序列同源性比較Fig.7 Homology comparison of MHX proteins from different plants
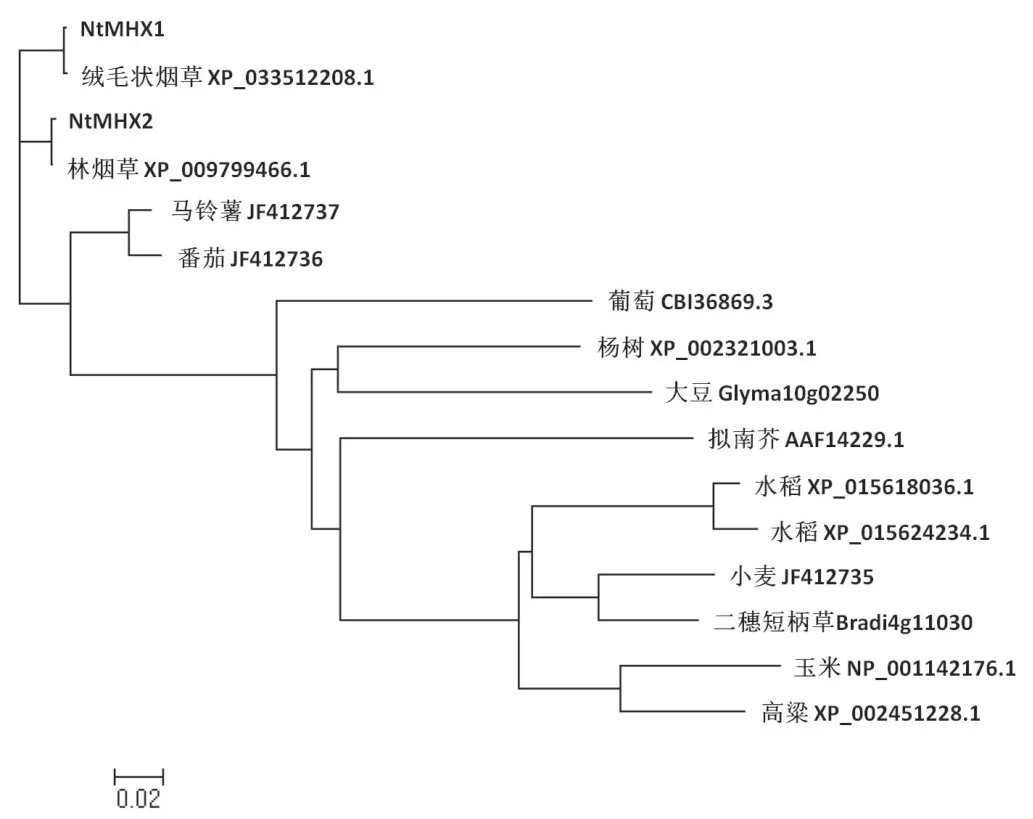
圖8 NtMHX1和NtMHX2的系統進化分析Fig.8 Phylogenetic analysis of NtMHX1 and NtMHX2
選擇部分序列相似性較高、功能注釋明確或有文獻報道的MHX蛋白序列,利用Clustral X進行序列比對(圖7),發現這兩個蛋白具有保守的11個跨膜結構域。煙草NtMHX1和NtMHX2蛋白與茄科植物番茄、馬鈴薯具有100 %的序列覆蓋度,與野生番茄序列一致性最高,分別為92.78 %和91.77 %,與馬鈴薯MHX蛋白序列一致性分別為92.59 %和91.77 %。用MEGA5.0進行系統進化樹的繪制(圖8),結果可以看出,煙草和同屬茄科植物的馬鈴薯、番茄MHX序列處于同一分枝上。序列比對和聚類分析結果均表明煙草NtMHX1、NtMHX2蛋白與番茄和馬鈴薯的親緣關系最近。
2.4 NtMHX基因的組織表達特性
分別從未處理的煙草的根、莖、葉、花中提取RNA進行熒光定量分析得到結果顯示(圖9),NtMHX1和NtMHX2基因在根中表達量最低,在莖中表達量最高。
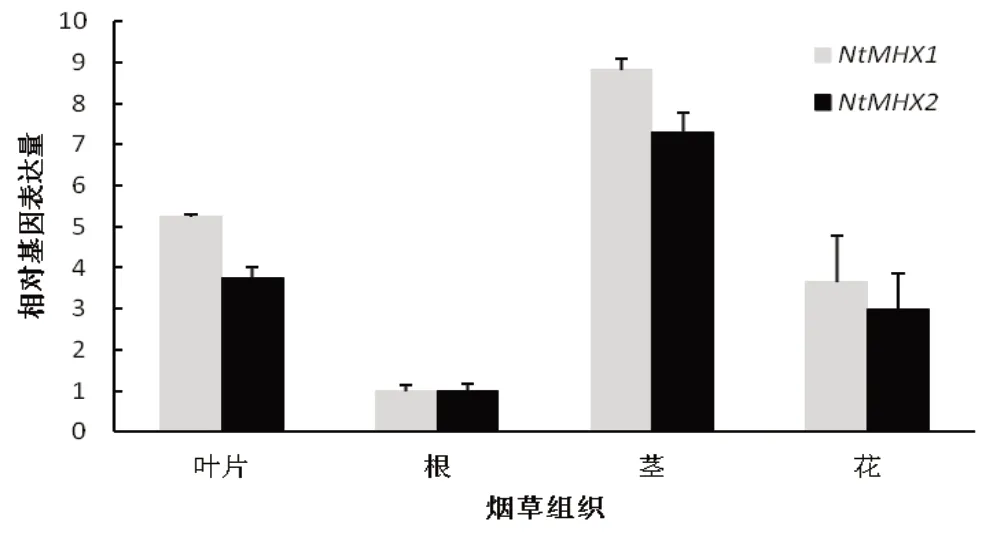
圖9 NtMHX1和NtMHX2基因的組織表達特性Fig.9 The relative expression level of NtMHX1 and NtMHX2 in different tissues of Nicotiana tabacum
2.5 金屬離子處理對NtMHX基因表達的影響
將苗期煙草經過不同離子處理后,檢測NtMHX1和NtMHX2基因在葉片中的相對表達量。結果(圖10)顯 示,50 mmol/L Mg2+處 理 后,NtMHX1和NtMHX2基因表達水平迅速升高并在1 d時達到最大值(分別為2.88和2.64)后逐漸下降,在6 d左右恢復至處理前水平。50 mmol/L Mn2+處理后,NtMHX1和NtMHX2基因轉錄水平呈現先升高后下降的趨勢,在處理后2天達到最大值(分別為3.80和3.90)。100 mmol/L Ca2+處理后,NtMHX1和NtMHX2基因的相對表達量迅速升高,在處理后2 d達到最大值(6.08和5.75)后逐漸減低。100 mmol/L K+處理后,NtMHX2基因轉錄水平先升高后降低,而NtMHX1基因表達水平呈現先升高后減低再升高的趨勢,兩個基因均再處理后2 d達到最大值,分別為5.66和6.34。100 mmol/L Na+處理后,NtMHX1和NtMHX2基因表達趨勢為先升高后減低,同樣在處理后2 d達到最大值(分別為6.84和6.02)。50 mM Zn2+處理后,NtMHX1和NtMHX2基因相對表達量先升高,在處理后2 d達到最大值(3.41和3.50)后減低。50 mmol/L Cu2+處理后,NtMHX1和NtMHX2基因轉錄本積累量先升高后減低,在處理后2 d達到最大值,分別為3.44和4.36。50 μmol/L Cd2+處 理 后,NtMHX1和NtMHX2基因轉錄水平迅速下降,處理后2 d的相對表達量為0.23和0.35,并在處理后4~6 d維持這一表達水平。
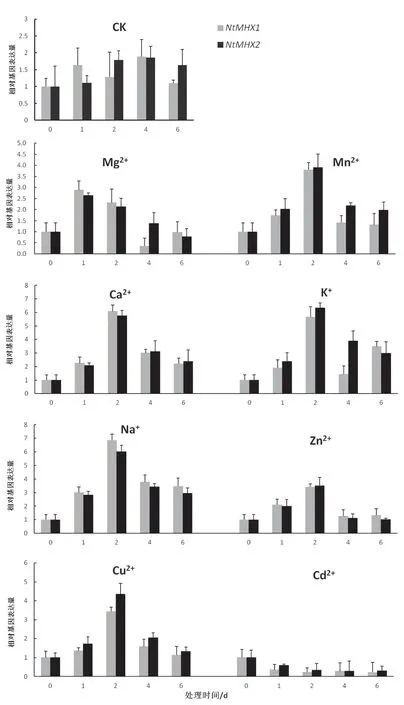
圖10 不同金屬離子對煙草葉片NtMHX1和NtMHX2表達水平的影響Fig.10 The effects of different metal ion treatments on the expression level of NtMHX1 and NtMHX2
3 討論
CaCA(Ca2+/cation antiporters)亞家族蛋白廣泛存在于不同的生物體內,從細菌到高等動植物都有該家族基因的報道。CaCA蛋白在維持植物體內離子平衡中發揮重要作用,進而影響了植物的生長發育[15]。根據蛋白結構和進化關系,CaCA亞家族可以分為CAX,CCX,NCL和MHX四個分支,其中MHX是植物中所特有的,且成員數量最少[14,16]。本文從煙草中克隆到2個MHX同源基因,命名為NtMHX1和NtMHX2,其中,NtMHX1來源于絨毛狀煙草,而NtMHX2來源于林煙草,并與茄科植物番茄和馬鈴薯的MHX蛋白位于同一分支上。
生物信息學分析發現,這兩個蛋白與其他植物中的MHX蛋白一樣,包含CaCA保守結構域,均有11個跨膜結構域,屬于跨膜蛋白,不含信號肽。亞細胞定位預測這兩個蛋白定位于細胞膜上,橡膠樹中該基因主要在液泡中表達[12-13],鼠耳芥(Arabidopsis Halleri)中AhMHX基因定位在液泡膜上[17]。與細胞質相比,液泡中的環境偏酸性,AtMHX蛋白將金屬離子扣留在液泡中,并將液泡中的質子釋放到細胞質中,本研究也發現煙草中的NtMHX蛋白等電點為5.66和5.63,為酸性蛋白。
AtMHX基因在擬南芥的維管組織,尤其是韌皮部的相對表達量較高[18],煙草NtMHX基因在根、莖、葉和花中都有表達,但莖中相對表達量最高,這與前人報道該基因主要在維管組織中表達的結果一致。鼠耳芥中AhMHX的基因表達量受植物體內Zn、Cd或者Mg含量狀態的影響不顯著[17]。本研究中不同金屬離子處理的表達模式分析顯示,Mn2+、Ca2+、K+、Na+、Zn2+處理后,NtMHX1和NtMHX2基因轉錄水平均先升高,后降低,并在處理2 d左右到達最大值,Cd2+處理后,NtMHX基因表達受到抑制,在2 d達到最低值。從不同金屬離子的響應時間及誘導水平來看,NtMHX基因對Mg2+的響應較早,Ca2+、K+、Na+離子處理對NtMHX基因的誘導水平要顯著高于Mg2+、Mn2+、Zn2+離子,只有Cd2+處理后,NtMHX基因的表達水平迅速下降,并維持低水平表達。分析發現Mn2+、Ca2+、K+、Na+、Zn2+是煙草生長發育所必需的營養元素,可以誘導NtMHX基因的表達,而有毒金屬元素鎘則可以抑制NtMHX基因的表達。推測NtMHX基因參與維持煙草體內金屬離子的平衡,其具體功能及機制有待進一步深入研究。
4 結論
(1)從栽培煙草中克隆到NtMHX1和NtMHX2基因,開放閱讀框全長分別是1623 bp和1641 bp,編碼540和546個氨基酸殘基,相對分子質量分別為60382.99 Da和60977.72 Da,均是包含11個跨膜結構域的酸性蛋白。
(2)進化分析表明,NtMHX1和NtMHX2與野生番茄和馬鈴薯的MHX蛋白高度相似。
(3)表達分析表明,NtMHX1和NtMHX2基因在煙草根、莖、葉、花中均有表達,且在莖中的表達水平最高。NtMHX基因的表達受到Mg2+、Mn2+、Ca2+、K+、Na+、Zn2+和Cu2+的誘導,并受到Cd2+的抑制,推測NtMHX基因在維持煙草體內金屬離子的平衡中發揮重要作用。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
