貴州省兩個藍莓品種組培苗和扦插苗干旱脅迫響應
2021-10-06 17:24:20周熒,王頔,聶飛*
南方農業 2021年20期
周 熒,王 頔,聶 飛*
(1.貴州省生物研究所,貴州貴陽 550009;2.湄潭縣林業局,貴州遵義 564100)
藍莓為叢生灌木,生產上一般采用組培和扦插2 種繁殖方式。藍莓為淺根系植株,無主根,根系纖細無根毛[1]。藍莓喜酸性疏松土壤,因此選擇抗旱性強的品種對藍莓苗木在不同生態區適應性極其重要。
貴州藍莓種植地多為山地,喀斯特地貌,小氣候類型多,主要土壤類型為黃壤、紅壤、黃棕壤、棕壤、石灰土和水稻土這幾大類,土壤保水能力差,土壤暫時性干旱頻繁發生[2-3]。干旱脅迫是植物常見的逆境脅迫之一,不僅制約植物的生長發育,并且對植物的結構產生影響[4]。植物對干旱脅迫的響應過程和受脅迫程度與植物自身的抗性相關,植物會從表型、生理功能、分子結構、生化代謝、形態適應、生長發育、生物量及產量等多種形式表現[5-7]。因此,從植物生理學及形態特征出發,是開展干旱脅迫下植物生理生化反應及葉片解剖結構的系統研究的重要方法。
1 材料與方法
1.1 材料
本試驗材料為貴州瑞藍果業有限公司麻江縣種苗繁育基地“萊克西”和“燦爛”2 個藍莓品種的移栽后1.5年生組培和扦插容器苗。“燦爛”組培苗用CLZ 表示,“燦爛”扦插苗用CLQ 表示,“萊克西”組培苗用LKXZ 表示,“萊克西”扦插苗用LKXQ 表示。
1.2 試驗方法
1.2.1 干旱脅迫
2017 年8 月,選擇生長健壯、枝葉均勻移栽后1.5年生幼苗進行水分脅迫處理,脅迫方式為土壤干旱漸進脅迫。每個藍莓品種各選擇長勢較一致的移栽幼苗25 株進行脅迫處理。先對試驗容器苗進行充分澆水3 d 后停止澆水,使其進行自然干旱,分別于停止澆水后第7 d、第14 d、第21 d、第28 d 隨機選取植株功能葉片,測定葉片相對含水量、葉片相對電導率、葉綠素含量、丙二醛(MDA)含量、脯氨酸(Pro)含量,每次采樣時對土壤進行含水量的測定,每個指標測定3 次重復。
1.2.2 葉片切片制作
在脅迫第7 d 和第28 d 分別選取“燦爛”“萊克西”2個藍莓品種組培和扦插的1.5 年生苗作為試驗材料,分別將采集的大小一致無病蟲害、無機械損傷的葉片跨中脈截取0.5 cm×0.5 cm 作為材料,用FAA 固定液固定24 h以上,梯度酒精和二甲苯進行脫水和透明,進行常規石蠟切片制作,切片厚度8~10 μm,番紅-固綠染色,樹脂膠封片,Nikon eclipse-e200 顯微鏡下觀察、隨機選取30 個視野進行拍照測定。
1.3 指標測定
1)土壤含水量。采用土壤烘干法測定土壤含水量。
2)葉片相對電導率測定。參考陳文榮等的改進方法采用佑科DDS-307A 型電導率儀測定[8],將待測葉片擦洗干凈,每個樣品稱取0.1 g 放入用20 mL 去離子水,用注射器抽真空,搖床振蕩3 h,靜置1 h,測定初電導率及所用去離子水的電導空白,沸水浴15 min 殺死組織細胞,靜置1 h,測終電導率。相對電導率的計算公式為(1):
3)葉片相對含水量。取鮮葉稱鮮葉質量,在蒸餾水中用注射器抽取真空,浸泡24 h 后稱其飽和鮮樣質量,之后在100~105 ℃下烘干,稱其干樣質量。相對含水量的計算公式為(2):

4)葉綠素、丙二醛(MDA)及游離脯氨酸(Pro)含量。葉綠素含量采用丙酮乙醇1 ∶1 浸泡提取比色,丙二醛(MDA)含量采用硫代巴比妥酸法,游離脯氨酸(Pro)采用酸性茚三酮法[9]。
1.4 數據處理
將試驗數據采用Excel 2013 進行錄入,采用spss 18.0 進行數據的處理,多重比較分析,單因素方差分析、隸屬函數平均值法綜合評價苗木抗旱性。
隸屬函數平均值法中,若為正相關,則:

若為負相關,則:

式中:i表示某個品種;j表示某項指標;Uij表示i種類j指標的抗旱隸屬函數值;Xij表示i種類j指標的測定值;Xjmin表示所有種類j指標的最小值;Xjmax表示所有種類j指標的最大值。
權重應用客觀賦權法進行計算:

其中Ij是一個無量綱數,表示某評價指標在干旱脅迫下的測定值相對于對照組的比值。Cj是第j個指標正常組的測定值。Sj是第j個指標在某個脅迫處理下所測定的平均值。
如果是負相關,則計算式為Ij=Sj/Cj。最后通過歸一化,計算出每個評價指標的權重:Wj=Ij/∑Ij。綜合評價值:D=∑(Uij×Wj)。
2 結果與分析
2.1 干旱脅迫下土壤含水量的變化
不同干旱處理時間土壤含水量變化如圖1 所示。由圖1 可知(小寫字母代表顯著性p<0.05,下同),不同脅迫時間土壤相對含水量差異顯著(p<0.05);處理第7 d,土壤含水量均值為84.72%±1.14%(正常生長),第28 d 的土壤含水量均值為24.78%±0.85%(重度干旱)。隨著干旱脅迫的天數增加,土壤含水量下降顯著,從第7 d 到第14 d 時下降最明顯,整體下降33.30%。

圖1 不同干旱處理時間土壤含水量變化
2.2 干旱脅迫下藍莓葉片葉綠素含量的變化
不同干旱時間葉綠素含量變化如圖2 所示。由圖2可知,隨著干旱脅迫增加,葉綠素含量逐漸降低。“萊克西”藍莓在第14 d 葉綠素含量下降明顯,扦插苗和組培苗比第7 d 分別下降了32.52%、51.14%;“萊克西”藍莓組培苗從第14 d 后變幅很小,“萊克西”藍莓扦插苗在干旱脅迫期間變幅最大,第28 d 葉綠素含量最小為(1.16±0.02)mg·g-1,比第7 d 下降了67.48%。“燦爛”藍莓在第21 d 明顯下降;“燦爛”藍莓組培苗在后期變化緩慢,“燦爛”藍莓扦插苗整體變化較明顯。
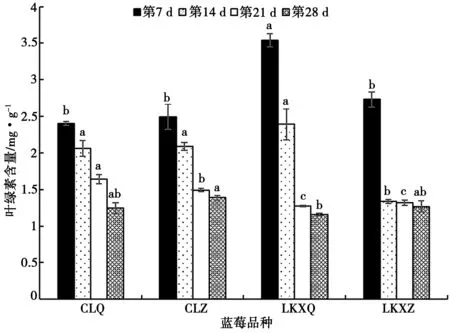
圖2 不同干旱時間葉綠素含量變化
2.3 干旱脅迫下藍莓葉片丙二醛含量的變化
不同干旱時間葉片丙二醛(MDA)含量變化如圖3所示,隨著干旱程度的增加,丙二醛含量呈增加趨勢。干旱前期,“萊克西”藍莓和“燦爛”藍莓中MDA 含量差異顯著(p<0.05),“萊克西”藍莓中MDA 含量變幅較小,組培苗和扦插苗中MDA 含量的變幅分別僅為3.56%、7.78%;干旱后期,“燦爛”藍莓中MDA 含量的變幅明顯弱于“萊克西”藍莓,組培苗和扦插苗的變幅分別為11.32%、5.43%。在干旱脅迫時MDA 含量相差最大的是“燦爛”藍莓組培苗,為58.54 nmol·g-1,其次是“燦爛”藍莓扦插苗,變化為44.89 nmol·g-1,“萊克西”藍莓的扦插苗和組培苗變化差距不大,分別為42.07 nmol·g-1、42.88 nmol·g-1。

圖3 不同干旱時間葉片MDA 含量變化
2.4 干旱脅迫下脯氨酸含量變化
不同干旱時間葉片脯氨酸含量變化如圖4 所示。由圖4 可知,隨著干旱脅迫的增加,葉片脯氨酸含量逐漸增加,但增加幅度不一致。4 種類型的藍莓苗初始和結束時的脯氨酸含量差異明顯(p<0.05),初始時脯氨酸含量“萊克西”藍莓扦插苗最大,其次是“燦爛”藍莓扦插苗,脅迫第28 d 時脯氨酸含量從大到小依次是“萊克西”藍莓扦插苗、“萊克西”藍莓組培苗、“燦爛”藍莓扦插苗、“燦爛”藍莓組培苗,且4 種幼苗脯氨酸差異顯著(p<0.05)。“萊克西”藍莓扦插苗在第7 d 時含量最小,但是在第14 d 后脯氨酸含量均大于其他3 種幼苗,且第28 d 的含量是第7 d 的1.86 倍,說明“萊克西”藍莓扦插苗對干旱脅迫反應強烈。

圖4 不同干旱時間葉片脯氨酸含量變化
2.5 干旱脅迫下葉片相對含水量變化
不同干旱時間葉片相對含水量變化如圖5 所示,隨著水分脅迫程度的加強,藍莓葉片的相對含水量逐漸降低。脅迫初期葉片相對含水量最大的是“萊克西”藍莓組培苗,含量為82.80%,最小的是“燦爛”藍莓組培苗,含量為70.18%。脅迫第21 d 以前葉片相對含水量4 種幼苗均變化幅度較小,第28 d 時變化幅度較大,變化幅度最大的是“萊克西”藍莓扦插苗,最小的是“燦爛”藍莓扦插苗。
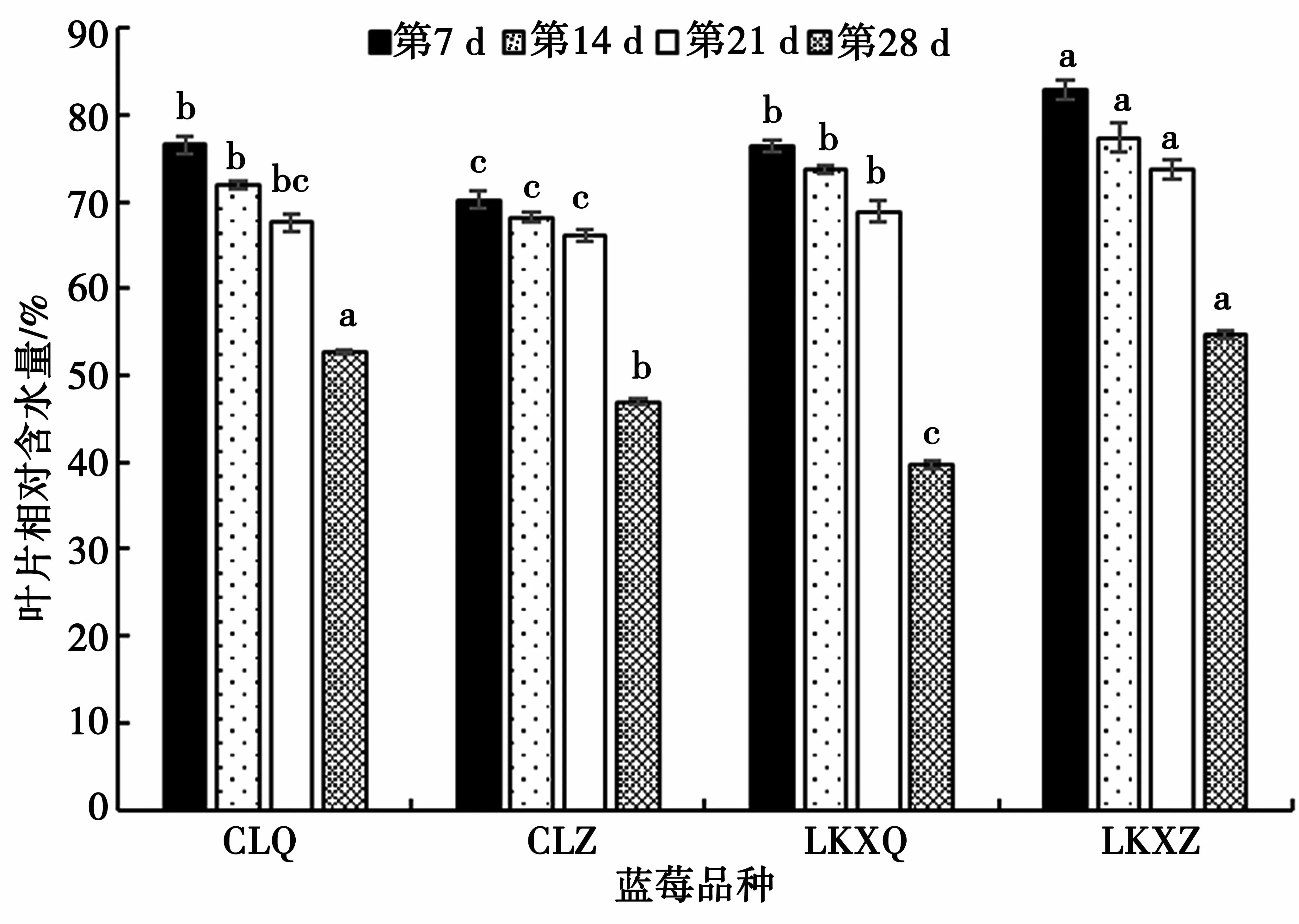
圖5 不同干旱時間葉片相對含水量變化
2.6 干旱脅迫下葉片相對電導率變化
干旱程度的加劇會造成植物的膜傷害,質膜透性增大,電導率值即可反映傷害程度,從而鑒定出植物的抗旱能力。隨著干旱時間的增加葉片相對電導率呈上升趨勢,如圖6 所示。脅迫初期,“燦爛”藍莓扦插苗和組培苗變幅明顯,第14 d 葉片相對電導率分別比第7 d 上升了1.97 倍和1.31 倍;在脅迫后期,“萊克西”藍莓扦插苗和組培苗相對電導率變化較明顯,第28 d 比第21 d上升了61%和87%。可見,4 種藍莓幼苗在不同脅迫時間受脅迫程度不一樣。

圖6 不同干旱時間葉片相對電導率變化
2.7 干旱脅迫下藍莓抗旱性評價
隸屬函數法是目前應用最廣的多指標評價方法[10-11],通過對4 種苗木各項理化指標進行隸屬函數綜合評價,由表1 可知,抗旱性由強到弱的是“燦爛”藍莓組培苗、“萊克西”藍莓組培苗、“萊克西”藍莓扦插苗、“燦爛”藍莓扦插苗。

表1 兩種繁殖方式下兩個品種抗旱性綜合評價
2.8 干旱脅迫下藍莓葉片結構變化
對干旱脅迫下藍莓葉片進行石蠟切片比較,結果表明,“燦爛”藍莓和“萊克西”藍莓的組培苗的上表皮厚度值、柵欄組織厚度值、葉片厚度值、柵海比值均高于扦插苗,見表2。

表2 葉片結構指標
3 結論與討論
隨著干旱程度加深,不同繁殖方式下4 種藍莓幼苗葉綠素含量、葉片相對含水量不斷下降,丙二醛含量、脯氨酸含量、葉片相對電導率不斷增加。在干旱脅迫下,幼苗在生理上首先表現為葉片水分虧缺;在生化方面水分脅迫導致活性氧等自由基積累,引起膜脂過氧化,膜脂過氧化產物MDA 含量和滲透調節物質脯氨酸含量增加,幼苗迅速啟動保護酶系統和積累滲透調節物質;在結構上由于水分虧缺引起細胞脫水,導致細胞膜損傷,質膜相對透性增加[12-14]。本試驗結果表明,4 種苗木在干旱脅迫下,土壤含水量降低,導致植株根系吸水困難,植株組織水分虧缺,葉片相對含水量降低,“萊克西”藍莓扦插苗葉片相對含水量降低最明顯,“燦爛”藍莓組培苗的葉片相對含水量變化最小,葉片相對含水量變化小,說明“燦爛”藍莓組培苗在干旱脅迫下能通過維持組織含水量來抵御干旱。此外,葉綠素含量是植物對干旱脅迫響應敏感的指標,大量的研究結果表明,干旱脅迫通過抑制葉綠素合成,并加速其分解,導致葉綠素含量直線下降[12,15]。本試驗結果表明,干旱脅迫下葉綠素均顯著下降,但在脅迫后期,“燦爛”藍莓和“萊克西”藍莓的組培苗均變化不明顯,這種變化可能是植株在受到脅迫后引起膜脂過氧化產生MDA 和脯氨酸阻礙了葉綠素的合成。
植物器官的形態結構與植物生理功能密切相關,生理變化間接導致形態結構的變化,表皮是葉片結構的基礎,植物解剖結構中表皮越厚,其抗旱、隔熱、持水能力一般均較強,對于干旱逆境的抵抗力越強[16-17]。本試驗中“燦爛”藍莓和“萊克西”藍莓兩種苗木,均是扦插苗的上表皮厚度大于組培苗葉小而厚,柵欄組織發達,柵欄組織與海綿組織比值高,角質層及上皮層厚,葉肉細胞小而排列緊密,氣孔下陷、表皮毛發達等都是抗旱性強的標志[18-21]。本試驗中,從葉片厚度來看,“燦爛”藍莓葉片厚度明顯厚于“萊克西”藍莓,一般認為葉片小而厚是植物抗旱的特征,細小的葉形可以減少水分蒸發的面積,葉片越厚,儲水能力越強[22]。隸屬函數的抗旱性綜合評價得出的結果與此一致,但在植物抗旱性綜合評價中不能用單一的指標進行評定,因為植物的抗旱性與植物本身的結構和外界環境有關,單一的指標評定很難反應客觀結果,可采用多種指標綜合評價[19]。
