地西他濱逆轉肺腺癌厄洛替尼耐藥的作用及機制
2021-10-14 08:58:46胡春生秦紅霞楊東林黃玖紅
中國藥理學通報 2021年10期
關鍵詞:耐藥
胡春生,唐 艷,秦紅霞,楊東林,黃玖紅
[重慶文理學院藥學院(創新靶向藥物國際研究院),創新靶向藥物國家地方聯合工程中心,重慶 402160]
表皮生長因子受體(epidermal growth factor receptor,EGFR)在非小細胞肺癌發生、發展過程中起重要作用。我國約40%的肺腺癌患者發生EGFR突變,此類患者對酪氨酸激酶抑制劑(tyrosine kinase inhibitorsn,TKI)具有良好的反應性,因此臨床上已將EGFR-TKI作為靶向藥物治療非小細胞肺癌(non-small cell lung cancer,NSCLC)患者EGFR陽性突變的一線治療方案。然而,大部分患者在接受第一、二、三代EGFR-TKI治療以后,約10-12個月出現耐藥現象,制約了EGFR-TKI在臨床上繼續使用[1]。
近年來,研究發現表觀遺傳機制在基因轉錄活性調控方面起重要作用。DNA 甲基化異常與多種疾病的發生密切相關,目前研究認為,CpG島甲基化在細胞分化、基因表達調控、基因突變、基因印記等多方面起重要作用。DNA甲基化在腫瘤中處于異常狀態,導致腫瘤轉移、耐藥等。研究表明,特定基因啟動子區的超甲基化將引起EGFR-TKI耐藥[2]。地西他濱(decitabine,DE)是DNA甲基轉移酶抑制劑,常常用于治療骨髓增生異常綜合征[3],但對逆轉肺腺癌EGFR-TKI耐藥研究尚少,且其作用機制仍不十分清楚。因此,本文考察了地西他濱對肺腺癌HCC827厄洛替尼(erlotinib,ER)耐藥細胞的增殖、遷移、侵襲抑制及細胞周期阻滯的作用,并探討其主要作用機制。
1 材料與方法
1.1 實驗材料及儀器
1.1.1細胞株 人肺腺癌細胞株HCC827/ER耐藥細胞,采用厄洛替尼低濃度梯度遞增誘導法由重慶文理學院藥學院建立并保存,采用MTT法檢測其耐藥倍數為350倍。
1.1.2試劑 地西他濱(S1200,Selleck)、厄洛替尼(S1023,Selleck);RPMI 1640培養基(SH30809.01,HyClone);胎牛血清(NTC-HK022,Natocor);青鏈霉素(SV30010,HyClone);胰蛋白酶(2046781,Gibco);甲基化陽性和陰性對照基因組(3522和3521,Takara);PCR試劑(RR001,TaKaRa);基因組DNA提取試劑盒(DP304,天根生化科技有限公司)。
1.1.3儀器 細胞培養箱(Thermo Model 3111);熒光顯微鏡(OLYMPUS IX73);多功能酶標儀(BioTek Cytation 5);凝膠成像系統(天能Tanon-5200 Multi);Western blot條帶激光掃描儀(LI-COR Odeyssey)。
1.2 實驗方法
1.2.1MTT法測定細胞增殖 將對數生長期的HCC827/ER細胞用胰酶消化下來,調整細胞濃度為6×107個·L-1,向96孔板中加入100 μL細胞懸液,CO2培養箱中孵育過夜使細胞貼壁;向每孔培養基中加入終濃度為0、0.05、0.5、1、10、25、50 μmol·L-1ER及0、5、10 μmol·L-1DE處理48 h;每孔中加入10 μL MTT試劑后繼續培養4 h;棄掉上清,加入200 μL DMSO,搖床80 r·min-1振搖30 min;通過酶標儀測定570 nm吸光值(optical density,OD值),細胞增殖率/%=(OD實驗-OD空白)/(OD對照-OD空白)×100%。
1.2.2BrdU熒光染色 將HCC827/ER細胞接種于12孔板中,將細胞分成ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+4組(ER-、DE-表示加入DMSO,ER+表示加入5 μmol·L-1ER,DE+表示加入10 μmol·L-1DE),將BrdU加入細胞培養液中,終濃度為5 mg·L-1,繼續培養48 h。吸棄培養基,4%多聚甲醛室溫固定30 min,PBS洗滌2次,用0.1%TritonX-100和10%山羊血清封閉1 h,棄掉封閉液,加入兔抗BrdU一抗4 ℃孵育過夜,PBS漂洗后加入山羊抗兔二抗(Cy3標記),避光室溫孵育2 h,PBS洗滌后加入DAPI染細胞核,顯微鏡下觀察、拍照。每組隨機拍5個視野,計算BrdU表達陽性率。
1.2.3克隆形成 將HCC827/ER細胞按1 000個/孔接種于6孔板中,將細胞分成ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+4組,繼續培養14 d。加入4%多聚甲醛固定細胞,PSB洗滌,然后加入1 mL結晶紫溶液室溫染色2 min,吸棄染液并用ddH2O洗滌5次。置于掃描儀中拍攝圖片,并進行克隆數分析。
1.2.4細胞劃痕和侵襲 細胞劃痕:使用對數生長期HCC827/ER細胞,接種于12孔板中繼續培養待細胞鋪滿細胞板時,用細胞劃痕工具沿孔中線劃線,于劃線后0、12、24 h顯微鏡下觀察拍照。細胞侵襲:將200 μL無血清培養基稀釋的對數期細胞加入附有Matrigel的Transwell小室,下室加入800 μL完全培養基,并按ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+分組加入相應藥物繼續培養24 h,取出Transwell小室,棄去培養基,并用無菌棉簽擦拭掉上室底細胞,4%多聚甲醛室溫固定,結晶紫染色后,置于顯微鏡下觀察計數。
1.2.5DNA的硫化修飾及甲基化特異性PCR 收集HCC827/ER細胞,利用基因組DNA提取試劑盒提取DNA并定量。取1 μg基因組DNA,加入去離子水補足20 μL,采用甲基化DNA檢測試劑盒對DNA進行處理,簡要步驟如下,向上述DNA樣品中加入2.2 μL的M-Dilution buffer,42 ℃水浴30 min,繼續加入220 μL CT conversion reagent混勻,80 ℃避光孵育60 min,再加入480 μL M-buffer PA,然后通過DNA吸附柱純化修飾后的DNA,最后溶于20 μL去離子水中。以修飾后的DNA為模板,LMNA甲基化引物(M):UP 5′-TTTACGTTTGTTAGGAGTA AGTCGA-3′,Down 5′-ATAACCGACAAATTAACAAC GCT-3′。LMNA非甲基化引物(U):UP 5′-TTTATG TTTGTTAGGAGTAAGTTGA-3′,Down 5′-CCATAACC AACAAATTAACAACACT-3′,進行甲基化特異性PCR。PCR條件為:95 ℃ 預變性5 min;95 ℃變性30 s、58 ℃退火30 s、72 ℃延伸30 s,共35個循環;1%瓊脂糖凝膠電泳,80 V,30 min。凝膠置于凝膠成像系統中拍照。
1.2.6Western blot分析蛋白表達 使用RIPA裂解液(含蛋白酶抑制劑)分別提取ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+4組HCC827/ER細胞的總蛋白,并用BCA蛋白檢測試劑盒進行蛋白定量。上樣20 μg總蛋白進行10% SDS-PAGE膠電泳;采用濕轉方法(100 V,120 min)將PAGE膠上的蛋白轉印至PVDF膜上,然后5%脫脂牛奶室溫封閉2 h,TBST洗滌5次,每次2 min;laminA/C、CyclinB1、CyclinA2、CyclinD1、CDC2、CDK4、CDK6一抗4 ℃孵育過夜,TBST洗滌5次,每次2 min;對應IRDye標記二抗稀釋比例為1 ∶10 000,室溫孵育1 h,TBST洗滌5次,每次2 min;采用LI-COR Odeyssey激光掃描儀檢測目的條帶并分析檢測結果。
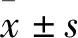
2 結果
2.1 地西他濱增強HCC827/ER細胞對厄洛替尼的敏感性MTT法檢測結果顯示,1、2.5、5、10、20、30、50、100 μmol·L-1的地西他濱處理HCC827/ER細胞48 h的抑制率都在20%左右,且無劑量依賴效應(Fig1A),因此選擇5、10 μmol·L-1地西他濱用于聯合用藥實驗。當地西他濱聯合不同濃度厄洛替尼(0.05、0.1、0.5、1、5、10、25、50 μmol·L-1)處理耐藥細胞,其細胞抑制率都在50%左右,增加HCC827/ER細胞對厄洛替尼的敏感性,且無劑量依賴效應(Fig1B)。利用Compusyn軟件計算聯合用藥的聯合指數(combination index,CI值),結果顯示CI<1(協同),結果表明地西他濱增強厄洛替尼抑制HCC827/ER耐藥細胞增殖的作用。
2.2 地西他濱聯合厄洛替尼抑制HCC827/ER細胞遷移和侵襲細胞劃痕實驗結果顯示(Fig2A、2B、2C),劃痕12 h后,細胞傷痕愈合率在ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+組分別為(57.3±4.2)%、(49.0±6.6)%、(34.7±2.1)%、(17±2.5)%;劃痕24 h后,各組細胞傷痕愈合率分別為(96.7±5.8)%、(76.3±1.5)%、(57.3±2.1)%、(45.7±1.5)%。以上結果表明,地西他濱具有抑制HCC827/ER細胞遷移的作用。當地西他濱與厄洛替尼聯合使用時可顯著抑制HCC827/ER遷移,與單獨使用地西他濱相比差異具有顯著性。另外,通過細胞侵襲實驗(Fig2D、2E),在ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+組HCC827/ER細胞穿過Transwell膜的數量分別為501±9、339±12、253±20、63±15。與DMSO組相比,單獨使用地西他濱對HCC827/ER細胞侵襲能力具有一定的抑制作用,與厄洛替尼聯合使用后顯著抑制HCC827/ER細胞侵襲(P<0.01)。

Fig 1 Decitabine enhanced anti-cancer effect of erlotinib to lung adenocarcinoma HCC827/ER n=6)A:The effect of decitabine on HCC827 and HCC827/ER cell viability.**P<0.01 vs DMSO group;B:The effect of decitabine in combination with erlotinib on cell viablility in HCC827/ER cells.**P<0.01 vs ER group;##P<0.01 vs only 5 μmol·L-1 or 10 μmol·L-1 DE group.

Fig 2 Effect of decitabine in combination with erlotinib on cell migration and invasion in HCC827/ER cells n=3)A:Decitabine,erlotinib alone or both together on the migration in HCC827/ER cell was detected by wound healing assay (×10);B,C:The percentage of wound healing area was analyzed at 12 h and 24 h after scratch;D:Decitabine,erlotinib alone or both together on the invasion in HCC827/ER cell was detected by Transwell(×20);E:The number of invasion cells was analyzed.*P<0.05,**P<0.01 vs ER-/DE-group.

Fig 3 Effect of decitabine in combination with erlotinib on cell proliferation ability and clonogenicity in vitro n=3)A,B:The cell proliferation ability was detected by BrdU assay(×20);C,D:Colony formation was performed with HCC827/ER cell for two weeks.**P<0.01 vs ER-/DE- group;##P<0.01 vs ER+/DE- or ER-/DE+ group.
2.3 地西他濱聯合厄洛替尼抑制HCC827/ER細胞增殖及克隆形成通過BrdU參入實驗(Fig3A、3B),BrdU/DAPI的比率在ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+組分別為0.64±0.08、0.39±0.01、0.53±0.19、0.20±0.04。以上結果表明,單獨應用5 μmol·L-1厄洛替尼可抑制HCC827/ER細胞增殖,聯合地西他濱后可顯著抑制HCC827/ER細胞增殖(P<0.01),然而單用地西他濱(10 μmol·L-1)對耐藥細胞增殖抑制不明顯。進一步通過克隆形成實驗結果顯示(Fig3C、3D),在ER-/DE-、ER+/DE-、ER-/DE+、ER+/DE+組中克隆數分別為216±15、101±9、129±11、52±8。與厄洛替尼(5 μmol·L-1)和地西他濱(10 μmol·L-1)單藥組相比,聯合用藥組細胞細胞集落數量和大小形態均小于單藥處理組。地西他濱聯合厄洛替尼對HCC827/ER細胞克隆形成率明顯下降,表明兩藥聯合對HCC827/ER細胞的增殖和成瘤能力具有明顯抑制作用。
2.4 地西他濱促進核纖層蛋白laminA/C表達甲基化特異性PCR檢測了laminA/C基因啟動子區甲基化情況(Fig4),在HCC827/ER細胞中laminA/C基因啟動子區甲基化水平明顯高于HCC827親本細胞,Western blot結果顯示在蛋白水平上,HCC827/ER細胞中laminA/C蛋白表達低于HCC827細胞,這表明啟動子區高甲基化抑制了laminA/C的蛋白表達。向HCC827/ER細胞中加入地西他濱后,抑制了laminA/C啟動子區甲基化,促進laminA/C蛋白表達,從而增強了耐藥細胞對厄洛替尼的敏感性。

Fig 4 Effect of decitabine in combination with erlotinib on methylation of laminA/C promoter and protein expressionA:Expression of laminA/C protein significantly decreased in HCC827/ER cells compared to HCC827 cells;B:Methylation analysis was detected by MSP.“+”:Hypermethylation of the genome,“-”:Low methylation of genome,“M”:Methylation,“U”:Unmethylation;C:LaminA/C protein was detected in HCC827/ER cells treated with decitabine or erlotinib.

Fig 5 Effect of decitabine in combination with erlotinib on cell cyclin related protein expression
2.5 地西他濱聯合厄洛替尼對細胞周期蛋白的抑制作用如Fig5所示,在聯合地西他濱和厄洛替尼的HCC827/ER細胞中,細胞周期蛋白CyclinA2、CyclinB1、CyclinD1表達下調,同時協同蛋白CDC2、CDK4、CDK6表達也下調;周期抑制P21蛋白表達上調。結果表明聯合地西他濱與厄洛替尼可通過細胞周期阻滯,抑制細胞增殖,促進HCC827/ER耐藥細胞死亡。
3 討論
本課題組研究發現,核纖層蛋白laminA/C在肺腺癌HCC827/ER厄洛替尼耐藥細胞株中的表達顯著低于親本細胞HCC827。在此基礎上,通過甲基化特異性PCR發現耐藥細胞中laminA/C啟動子區甲基化修飾高于親本細胞,且地西他濱能夠抑制laminA/C啟動子區甲基化修飾,促進laminA/C蛋白表達,逆轉HCC827/ER細胞對厄洛替尼的耐藥。以上結果表明超甲基化修飾抑制laminA/C蛋白表達,從而促進細胞厄洛替尼耐藥。本研究通過深入研究去甲基化酶抑制劑地西他濱逆轉肺腺癌厄洛替尼的耐藥及機制,以期為肺腺癌患者EGFR-TKI耐藥后精準治療提供理論基礎。
非小細胞肺癌EGFR-TKI獲得性耐藥機制涉及多基因、多途徑的復雜調控過程,部分耐藥機制尚不明確。已有大量文獻報道EGFR-TKI耐藥機制主要集中在以下幾個方面[4-8]:(1)EGFR基因的二次突變(T790M突變、C797S突變等),其中第一、二代EGFR-TKI發生T790M突變。針對T790M突變的第三代EGFR-TKI藥物在使用10個月左右患者發生C797S突變,病情再次出現進展;(2)c-MET及HER2原癌基因擴增,該類基因擴增可導致下游PI3K/AKT信號通路異常活化引起腫瘤對EGFR-TKI耐藥;(3)K-Ras基因突變,其基因的活化突變會導致患者對EGFR-TKI耐藥;(4)PTEN功能缺陷,其抑癌作用喪失,導致EGFR-TKI無法下調凋亡基因,從而引起腫瘤耐藥;(5)上皮間質轉化,在耐藥發生過程中,伴隨著上皮細胞向間質細胞轉化的形態學改變,同時伴有上皮標志物減少和間質標志物增多。本課題組前期研究和相關文獻報道顯示,肺腺癌HCC827細胞EGFR-TKI耐藥后發生EMT轉化,主要表現E-cadherin表達降低,N-cadherin和vimentin表達上調[9]。同時,本研究結果顯示,HCC827/ER耐藥細胞中核纖層蛋白laminA/C表達顯著低于親本細胞株,該發現提示laminA/C可能參與EMT過程介導EGFR-TKI耐藥。
本研究發現,利用地西他濱處理HCC827/ER耐藥細胞以后,提高核纖層蛋白laminA/C表達,可以逆轉HCC827/ER對厄洛替尼的耐藥性,降低耐藥細胞增殖、遷移侵襲能力。laminA/C的研究主要集中在早老癥上,但近年來研究發現腫瘤中的laminA/C表達量明顯低于癌旁組織或正常組織,在腫瘤的發生、轉移和侵襲過程中發揮著重要作用[10-13]。Nardella和Cicchillitti課題組都發現laminA/C的低表達增加腫瘤細胞的惡性程度[11,14]。Zhang等[15]通過上調乳腺癌細胞laminA/C表達引起細胞骨架排列改變促進懸浮乳腺癌細胞快速貼壁,表明laminA/C對腫瘤轉移具有抑制作用。既往的研究發現,在非小細胞肺癌患者的基因組中存在高甲基化修飾。因此,非小細胞肺癌PC9/GR耐藥細胞株用去甲基化藥物5-Aza-cdR處理以后,可恢復對吉非替尼的敏感性,這可能抑制EGFR啟動子區甲基化水平有關[16]。另外,聯合使用去甲基化藥物5-Aza-cdR可以增強肺腺癌原發耐藥細胞株H1650、H1299對吉非替尼的敏感性[17]。laminA/C在肺腺癌中呈現表達降低的趨勢,這與其基因啟動子區高甲基化相關。本研究發現在耐藥細胞中laminA/C啟動子區甲基化水平高于親本細胞,抑制laminA/C蛋白表達導致HCC827細胞對厄洛替尼產生耐藥。利用地西他濱處理耐藥細胞后可抑制laminA/C啟動子區甲基化水平,增加laminA/C蛋白表達,逆轉HCC827/ER細胞對厄洛替尼的耐藥性,但laminA/C在調控肺腺癌EGFR-TKI耐藥中的機制還需深入研究。
EGFR受體被配體激活后,引起下游信號通路PI3K/AKT、Ras/MEK/MAPK等調控細胞增殖與分化、遷移與侵襲等[18]。HCC827/ER耐藥細胞中,細胞周期蛋白相關蛋白CyclinA2、CyclinB1、CyclinD1、CDK4、CDK6表達上調,促進耐藥細胞增殖;單獨使用厄洛替尼或地西他濱處理HCC827/ER耐藥細胞,其相關周期蛋白表達與DMSO對照組相比無明顯差異,但地西他濱聯合厄洛替尼可顯著抑制周期相關蛋白的表達,促進周期抑制P21蛋白表達,使耐藥細胞周期受到阻滯,引起細胞死亡。
綜上所述,本研究發現地西他濱可逆轉肺腺癌HCC827/ER細胞對厄洛替尼的耐藥性,主要通過抑制laminA/C啟動子區DNA甲基化水平,促進laminA/C蛋白表達,對細胞周期進行調控,抑制耐藥細胞增殖、遷移和侵襲,為非小細胞肺癌EGFR-TKI耐藥后用藥提供理論依據。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
