不同氮磷營養條件對海黍子生長及抗氧化能力的影響
2021-10-15 04:42:22王翔宇辛美麗詹冬梅吳海一
海洋漁業 2021年5期
關鍵詞:生長
呂 芳,王翔宇,辛美麗,詹冬梅,丁 剛,吳海一
(1.山東省海洋生物研究院,山東省大型海藻資源保護與應用工程技術研究中心,山東青島 266104;2.青島市大型海藻工程技術研究中心,山東青島 266104)
《2018年中國海洋生態環境狀況公報》顯示,我國近海海域水質主要超標要素為無機氮和活性磷酸鹽,呈富營養化狀態的海域面積達56 680 km2,氮、磷濃度在不同海域變化較大。由于大型海藻生產力很高,在生長過程中可大量吸收碳、氮、磷等生源要素,因此,栽培大型海藻是凈化養殖廢水、控制水域富營養化和保護生態環境的有效措施[1-3]。
海黍子(Sargassummuticum)屬褐藻門,馬尾藻科,是馬尾藻屬中一種重要的提取褐藻膠和多酚等活性物質的主要原料之一,具有巨大的潛在經濟價值。此外,海黍子還具有適應性廣、生長快、營養儲存庫大等特點,對海水中的富營養化物質和重金屬等有較強的生物吸附能力[4-5],具有重要的生態價值和較好的應用前景,從而成為修復富營養化水域和構建海藻場的研究熱點。氮、磷營養鹽是海藻細胞生長的重要限制性營養元素,對海藻的生長具有顯著影響[6-8],在一定的濃度范圍內會使藻類的生長速率升高,光合作用增加,但濃度過低或過高時,則對藻類的生長產生抑制作用。大型海藻對氮、磷營養鹽的吸收特性不僅因海藻種類存在差異,與海藻的生長環境和發育階段也顯著相關,盡管營養鹽濃度對海黍子生長的影響已有一些報道[5,9],但高濃度氮、磷對海黍子藻體內各種生化組分的生成和消耗以及抗氧化系統的調節等仍鮮見報道,而這方面的研究對弄清海黍子在人工增養殖過程中養殖環境的選擇及其在富營養化水域中進行生物修復所持續的時間和對環境的適應性等具有極其重要的意義。鑒于此,本研究通過實驗生態學方法,探討不同氮、磷營養條件對海黍子的生長、生理生化特性以及抗氧化能力的影響,旨在為豐富海黍子人工增養殖技術體系,進而為我國近海富營養化等環境污染的生物治理和可持續利用提供科學依據。
1 材料與方法
1.1 實驗材料
實驗用海黍子為2019年3月采集于山東榮成俚島近岸海域(37°25′N、122°59′E)潮下帶的野生群體。藻體采集后用保溫箱迅速運回實驗室,用海水反復清洗去除泥沙及雜質,置于循環水箱中充氣暫養,培養條件為溫度15℃,光照強度5 000 lx,鹽度30.5,光周期12L∶12D,5 d后用于實驗。
1.2 培養方法
實驗在1 L的三角瓶中進行,選取健康藻體,每瓶放置(4.0±0.1)g。培養液以天然海水(海水鹽度為基質,經0.45 μm濾膜過濾后,用NaNO3、NaNO2、(NH4)2SO4、KH2PO4加富氮、磷濃度,其中各試驗組中溶解無機氮(dissolved inorganic nitrogen,DIN)的濃度比設為5∶5∶1。實驗分組如下:T1(對 照 組,天 然 過 濾 海 水),T2P),T3(100μmol·L-1DIN+6.25μmol·L-1L-1DIN+37.5μmol·L-1每組設3個重復。培養條件同1.1,每隔一天更換培養液,實驗周期為15 d,在實驗的1、3、7、15 d分別取樣進行測定。
1.3 相對生長速率(relative growth rate,RGR)的測定
觀察藻體表觀性狀并稱量鮮質量,按下式計算相對生長速率:
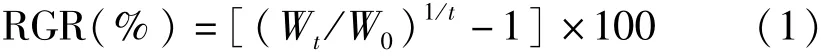
式(1)中,Wt為實驗中期或結束時鮮藻體質量(g),W0為實驗開始時鮮藻體質量(g),t為培養時間(d)。
1.4 氮、磷吸收速率測定
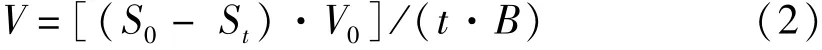
式(2)中,V為吸收速率[μmol·(g·h)-1],S0、St分別為取樣時間間隔的起始和結束時培養液中氮、磷的濃度,V0為每次取樣時間間隔起始的培養液體積(L),t為取樣的時間間隔(h),B為實驗開始時鮮藻體質量(g)。
1.5 光合色素[葉綠素a(chlorphyll a,Chl-a)和類胡蘿卜素(carotenoid,Car)]含量測定
光合色素的提取和測定參照呂芳等[10]的方法測定。取0.1 g新鮮藻體在液氮中研磨成勻漿狀,加入8 mL 80%丙酮,然后置于4℃黑暗處抽提24 h。于4℃、4 000 r·min-1離心10 min,棄沉淀,上清用80%丙酮定容至10 mL。以80%丙酮作為空白對照,分別測定665、652、510、480 nm波長處的吸光值。重復3次以上,計算平均值。按下式計算葉綠素a和類胡蘿卜素的含量:
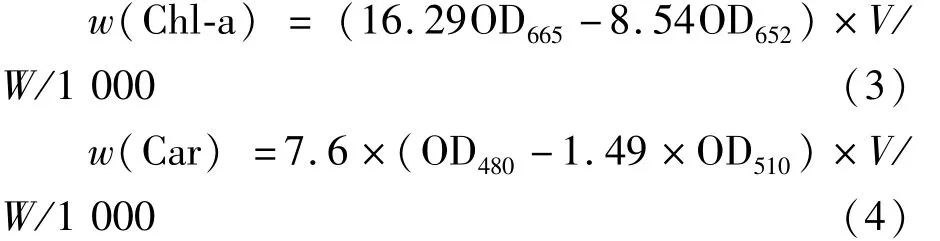
式(3)、式(4)中,V為浸提丙酮的體積(mL),W為鮮藻體質量(g),葉綠素a和類胡蘿卜素含量的單位為mg·g-1。
1.6 可溶性蛋白含量測定
可溶性蛋白采用南京建成蛋白檢測試劑盒-考馬斯亮藍法測定。取1.0 g新鮮藻體在液氮中研磨成勻漿狀,加蒸餾水后離心定容到10 mL,作為待測溶液。
1.7 抗氧化系統相關指標的測定
超氧化物歧化酶(superoxide dismutase,SOD)、過氧化氫酶(catalase,CAT)、過氧化物酶(peroxidase,POD)的活性和丙二醛(malondialdehyde,MDA)含量均采用南京建成生物工程研究所試劑盒測定。取1.0 g新鮮藻體在液氮中研磨成勻漿狀,加入4 mL提取液(50 mmol·L-1磷酸緩沖液,pH 7.0;0.1% Triton X-100;1% PVP),離心取上清作為待測溶液。
1.8 數據分析
采用SPSS 13.0進行數據統計分析,用單因素方差分析(one-way ANOVA)的Duncan法進行組間多重比較,當P<0.05時為有顯著性差異。結果采用Origin 9.0進行分析繪圖。
2 結果與分析
2.1 氮、磷濃度對海黍子生長的影響
比較不同氮、磷濃度對海黍子相對生長速率的影響發現,培養3 d時,藻體RGR隨著氮、磷濃度的增加,呈現先增加后減少的趨勢,除T6試驗組與對照組無顯著性差異外(P>0.05),T2、T3、T4、T5試驗組的RGR隨著氮、磷濃度的增加較對照組分別上升了30.7%、61.7%、47.0%和23.8%;培養7 d時,各試驗組的RGR均較3 d時有所下降,其中T2試驗組仍顯著高于對照組,T6試驗組則顯著低于對照組(P<0.05),而其余3組均與對照組無顯著差異(P>0.05);培養15 d時,僅T2試驗組的RGR與對照組無顯著差異,其他試驗組均顯著低于對照組,且隨著氮、磷濃度的增加呈逐漸下降的趨勢,其中T6試驗組RGR出現負增長,這可能是因為氮、磷濃度過高對海黍子產生了毒害作用,抑制了其生長代謝過程(圖1)。
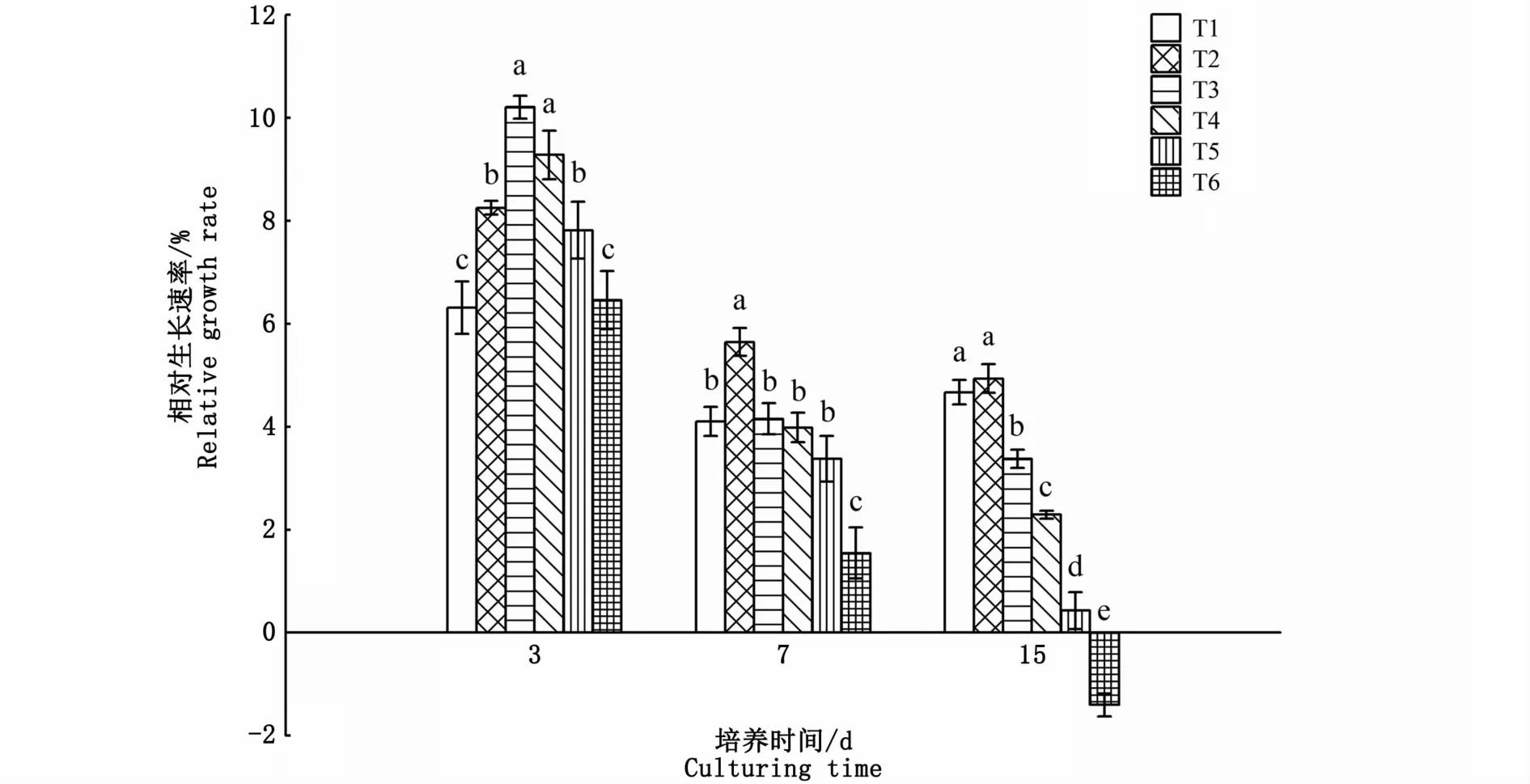
圖1 不同氮、磷濃度對海黍子相對生長速率的影響Fig.1 Effects of different N,P concentrations on relative growth rate of S.muticum
2.2 氮、磷濃度對海黍子吸收速率的影響
加富不同氮、磷濃度培養3 d后,海黍子對3種形態氮和磷的吸收速率見圖2。隨著氮、磷濃度的增加,藻體對3種形態氮和磷的吸收速率均呈逐漸升高的趨勢,且均較對照組存在顯著性差異(P<0.05);比較3種形態氮的吸收速率發現,海黍子對NH+4-N的吸收速率略高于NO-3-N,但兩者間差異不顯著(P>0.05),而對NO-2-N的吸收速率則顯著低于NH+4-N和NO-3-N。
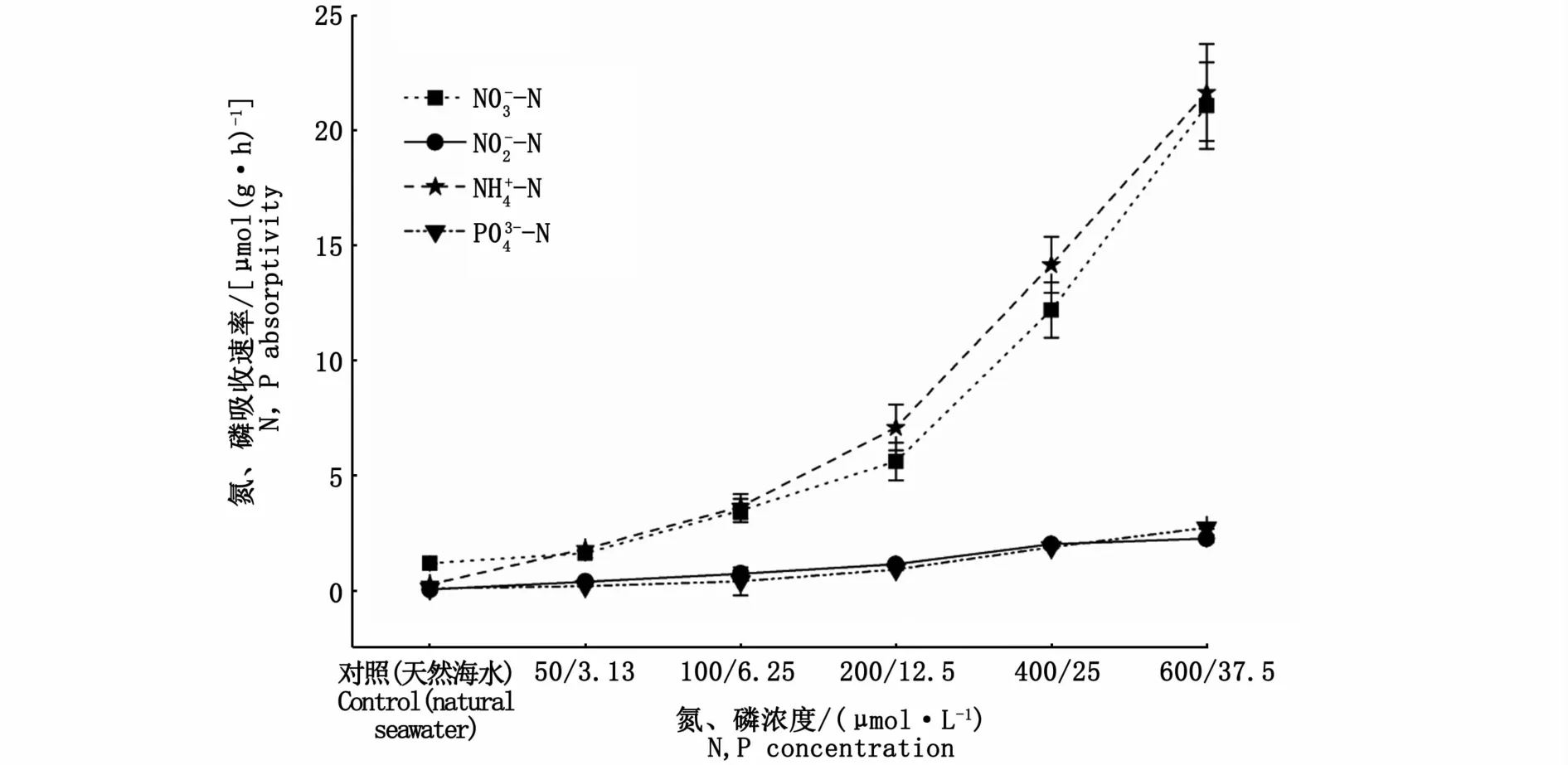
圖2 海黍子對不同形態氮和磷吸收速率的比較Fig.2 Comparison of different forms of N and P absorptivity rates in S.muticum
2.3 氮、磷濃度對光合色素含量的影響
海黍子光合色素的含量均受到氮、磷濃度和培養時間的影響。在3 d時,T3、T4、T5、T6試驗組葉綠素a的含量隨著氮磷濃度的升高均有增加;在7 d時,藻體葉綠素a的含量隨著氮、磷濃度的升高呈先上升后下降的趨勢,在T4試驗組中達到最大值,氮、磷濃度較低的4個試驗組藻體葉綠素a的含量顯著高于對照組(P<0.05);至15 d時,各試驗組葉綠素a的含量均有所下降,除T1試驗組與對照組無顯著差異外,其余4組則顯著低于對照組(P<0.05)(圖3-A)。
在培養3 d時,各試驗組類胡蘿卜素的含量均顯著升高,隨著氮、磷濃度的升高,T2、T3、T4、T5、T6試驗組分別比對照組高22.0%、26.1%、26.2%、39.5%和35.8%;培養7 d時,T3、T4、T5、T6試驗組類胡蘿卜素的含量仍顯著高于對照組,其中T4試驗組最高;培養15 d時,各試驗組變化各異,其中T2試驗組仍顯著高于對照組(P<0.05),T3試驗組與對照組差異不顯著(P>0.05),T4、T5、T6試驗組較對照組則顯著降低(圖3-B)。
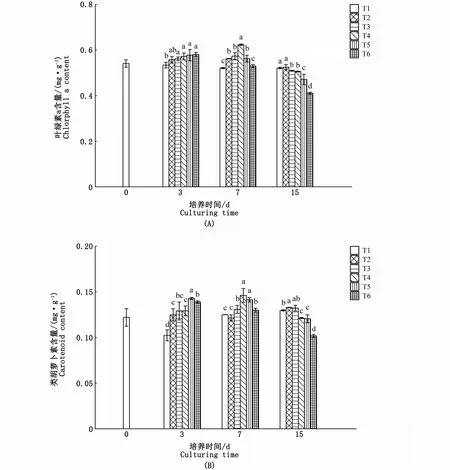
圖3 不同氮、磷濃度對海黍子光合色素含量的影響Fig.3 Effects of different N,P concentrations on photosynthetic pigments of S.muticum
2.4 氮、磷濃度對可溶性蛋白含量的影響
氮、磷濃度對海黍子可溶性蛋白含量的影響比較顯著,培養3 d時,T2、T3試驗組與對照組無顯著差異,而T4、T5、T6試驗組顯著升高,但3組間差異并不顯著(P>0.05);培養7 d時,各試驗組可溶性蛋白含量均顯著高于對照組,且隨著氮、磷濃度的升高呈先上升后下降的趨勢,其中T5試驗組最高;至15 d時,T2、T3、T4、T5、T6試驗組可溶性蛋白含量分別是對照組的1.16、1.25、1.25、2.06和1.61倍,差異顯著(P<0.05)(圖4)。
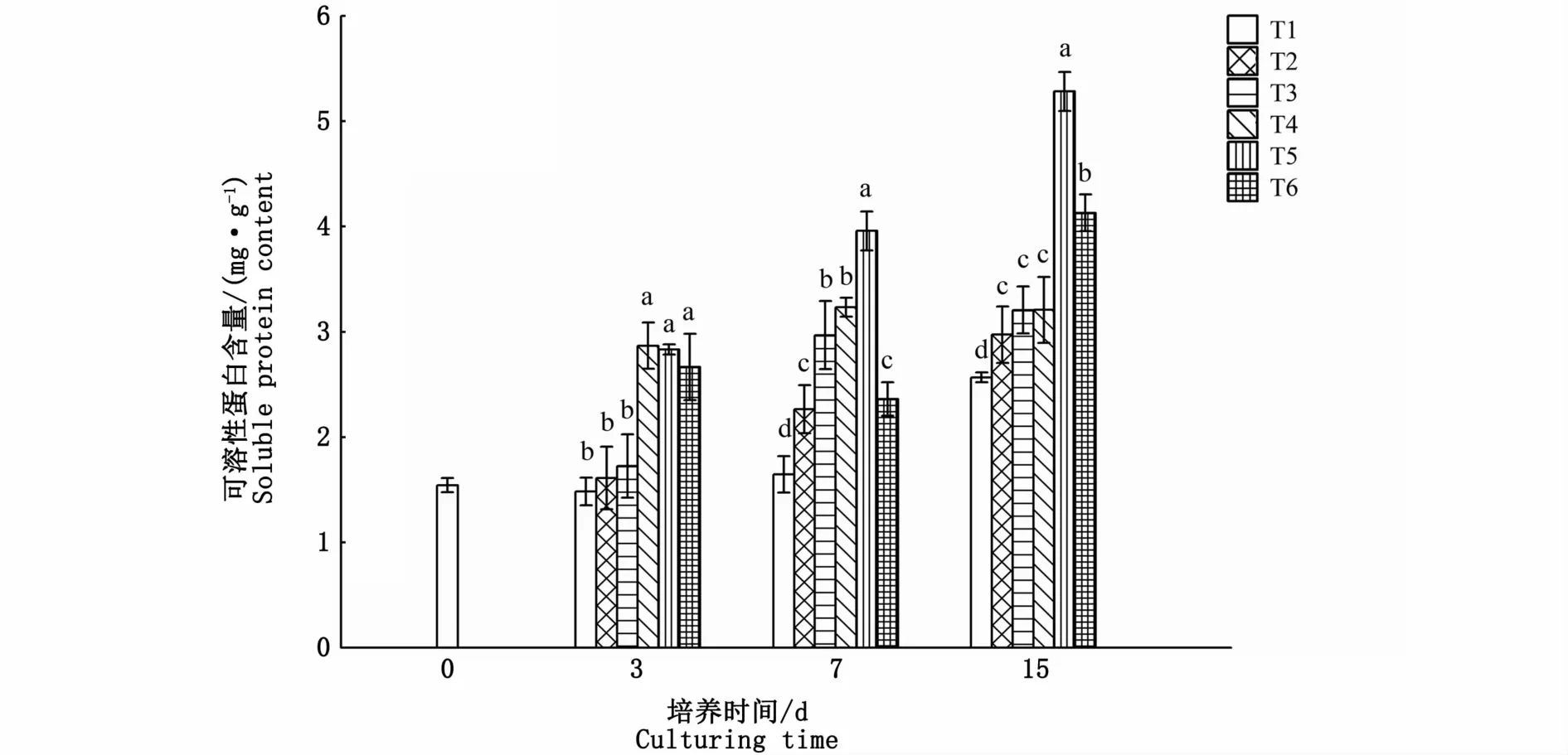
圖4 不同氮、磷濃度對海黍子可溶性蛋白含量的影響Fig.4 Effects of different N,P concentrations on soluble protein content of S.muticum
2.5 氮、磷濃度對SOD、CAT、POD活性和MDA含量的影響
各試驗組SOD活性的變化隨氮、磷濃度的升高呈現差異性,其中T2試驗組SOD活性在培養3 d時略有下降,7 d后與對照組無顯著性差異(P>0.05);T3試驗組在培養3 d時,其藻體SOD活性較對照組無顯著差異,7 d時,較對照組上升了13.8%,15 d又降至對照組水平;T4、T5、T6組在實驗期間SOD活性均保持在較高水平,15 d時分別比對照組高出15.3%、11.5%和12.8%(圖5-A)。
實驗期間,各試驗組藻體CAT活性隨氮、磷濃度的升高均呈先升高后下降的趨勢。T2試驗組CAT活性僅在培養7 d時略低于對照組,其他時間均與對照組無顯著差異(P>0.05);T6試驗組CAT活性在15 d內均顯著低于對照組,其中15 d時僅為對照組的52.2%;T3、T4、T5試驗組CAT活性則顯著高于對照組,至15 d時較對照組分別高10.2%、26.4%和56.8%(圖5-B)。
培養3 d時,POD活性試驗組T2、T3、T4藻體與對照組無顯著差異,而T5、T6組POD活性則顯著高于對照組(P<0.05);培養7 d時,藻體POD活性隨氮、磷濃度的升高呈逐漸上升的趨勢,其中T4、T5、T6組較對照組分別升高22.2%、47.9%和56.1%;至15 d時,除T6組POD活性略有下降外,其余各試驗組仍呈上升趨勢(圖5-C)。
MDA含量在實驗期間均隨著氮、磷濃度的增加呈逐漸上升的趨勢,除T2、T3組在培養3 d時MDA的含量略低于對照組外,其余各組MDA的含量在實驗期間較對照組均有不同程度的升高,尤其是T5、T6試驗組MDA含量在培養7 d后急劇升高,至15 d時較對照組分別上升了127.1%和312.7%(圖5-D)。
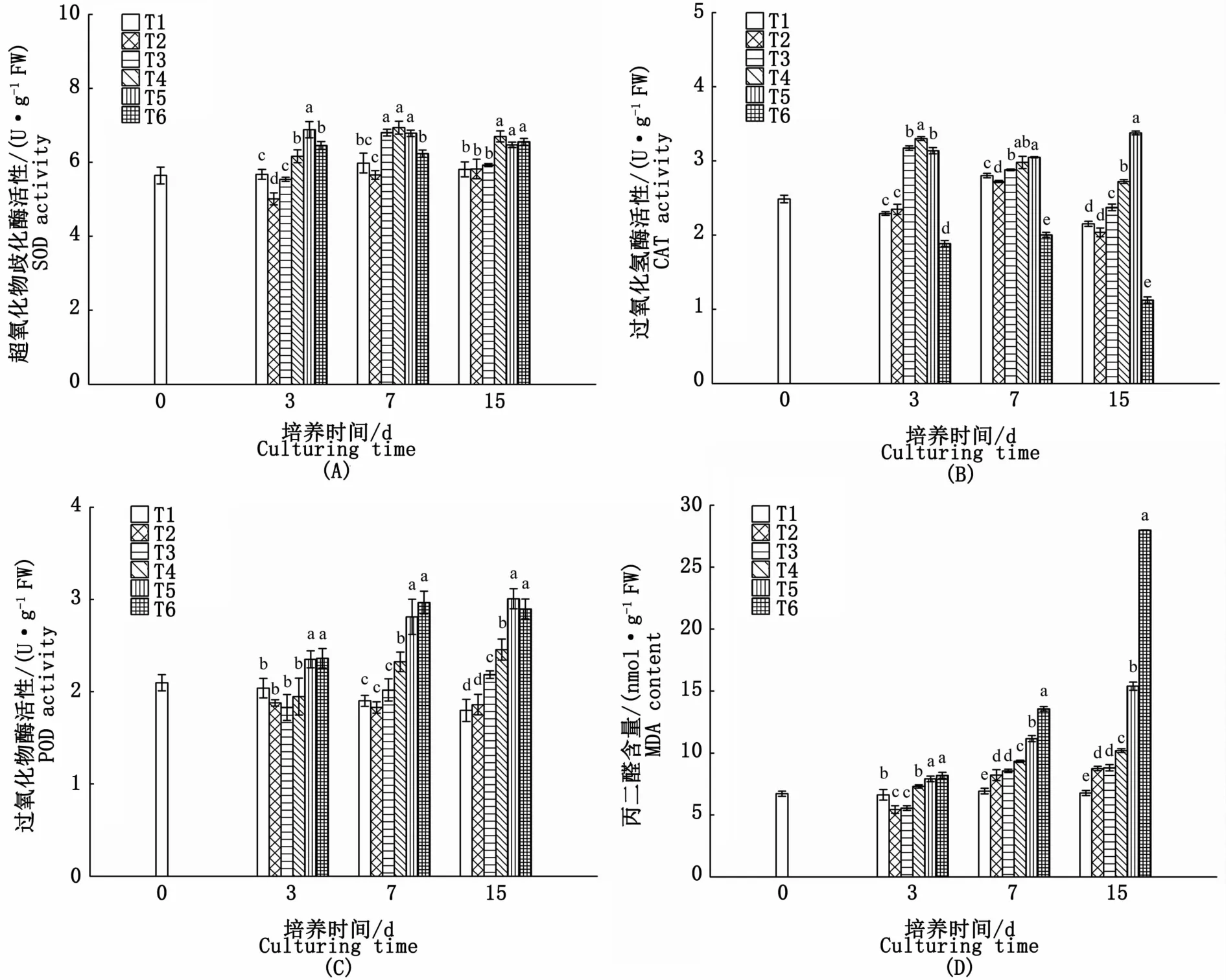
圖5 不同氮、磷濃度對海黍子抗氧化系統的影響Fig.5 Effects of different N,P concentrations on antioxidant system of S.muticum
3 討論
3.1 氮、磷濃度對海黍子生長的影響
海藻的生長受海水中營養鹽的影響,其中氮、磷是主要的限制性營養元素。本研究結果表明,氮、磷濃度和培養時間顯著影響海黍子的生長。氮、磷濃度在不高于400、25μmol·L-1下,培養3 d時,藻體的生長速率隨著氮、磷濃度的升高而增加,這與YU和YANG[6]報道的龍須菜(Gracilarialemaneiformis)在氮、磷濃度較高的情況下生長速率增大的結果一致,其原因是海水中營養鹽濃度的上升可使藻體利用營養生長的一系列生理過程的底物濃度升高,從而使海藻的同化作用增加,促進藻體生長。而當海水中氮、磷含量超過一定濃度后,其對藻類的生長則產生抑制作用,本研究也進一步證實了海藻生長存在臨界營養鹽濃度[11],超過該臨界值反而不利于海藻的生長。當繼續培養至7 d時,氮、磷濃度超過100、6.25μmol·L-1,隨著氮、磷濃度的升高藻體生長速率有下降的趨勢,可見該臨界營養鹽濃度與海藻的營養狀態有關,當海藻處于營養虧損狀態下,營養鹽加富會導致海藻出現超補償生長[12],當海藻處于營養過剩條件下,營養的持續加富會由促進作用轉為抑制作用。因此海黍子在人工增養殖過程中養殖海區的選擇尤為重要,氮、磷濃度在不超過400、25μmol·L-1時海黍子生長良好,當氮、磷濃度上升到600、37.5μmol·L-1時,海黍子的生長在比較短的時間內就會受到抑制。此外,氮磷比也是影響藻類生長的重要影響因素,這些都有待進一步的研究。
3.2 氮、磷濃度對海黍子吸收速率的影響
在培養3 d時,海黍子對氮、磷的吸收速率隨著外界營養鹽濃度的升高而增大,而同期藻體的生長速率卻呈先升高后下降的趨勢,原因可能是當氮、磷濃度達到海藻生長所需的臨界營養鹽濃度時,藻類生長達到飽和狀態,而營養鹽濃度過高則導致營養鹽在藻體內積累[6]。研究表明,海藻從環境中吸收的氮主要是形式,其中首先要被還原為才能為藻體所同化,所以當環境中和等氮源共存時,絕大多數藻類總是優先選擇吸收本研究發現,當和等量存在時,海黍子對兩種氮源的吸收無顯著差異,這可能與海黍子的生理狀態和實驗條件有關,也可能是不同形態氮之間的相互影響所致[14],具體的作用機制還有待進一步研究。
3.3 氮、磷濃度對海黍子生化特性的影響
海水中氮、磷濃度的高低在影響養殖海藻生長的同時,也對藻體內的各種生化組分(如色素、蛋白質等)的含量進行著調節。葉綠素作為藻類進行光合作用的主要色素,其含量的多少直接影響光合速率和光合能力,此外葉綠素也是藻體內一種重要的氮貯藏庫。研究表明,在外界氮源豐富的條件下,藻體細胞可以將外界的氮源合成葉綠素積累于葉綠體中,而在缺氮情況下,葉綠素就會降解釋放出氮以供應細胞的生長,進而起到調節氮源的作用[15],因此養殖海藻葉綠素含量的變化對研究海藻的生長特征具有重要意義。本研究中氮、磷加富培養3 d時,可促進海黍子藻體內葉綠素a的生成,但繼續培養至15 d,氮、磷濃度高于100、6.25μmol·L-1時反而阻止了葉綠素a的生成,在其他一些大型海藻中也得到類似的結果[6,16],推測其原因可能是高濃度的氮、磷抑制了葉綠素相關合成酶的活性,從而阻礙了葉綠素的合成,導致藻體內葉綠素含量的顯著下降。類胡蘿卜素是光合作用的輔助色素,也是一種光氧化保護劑,本研究顯示其含量在氮、磷加富3 d時顯著升高,至15 d,氮、磷濃度高于200、12.5μmol·L-1時則顯著降低,進一步說明過高濃度的氮、磷在長時間作用下會抑制海藻的生長。
藻體內可溶性蛋白是衡量藻類代謝水平的重要指標之一,其含量的升高有助于維持藻細胞的正常代謝,提高其抗逆性。NH+4或NO-3是藻體內蛋白質合成的底物,因此海水中氮濃度的變化對蛋白質的合成具有顯著影響。培養7 d后,各試驗組可溶性蛋白含量均顯著升高,說明海黍子可能通過提高可溶性蛋白的含量來增加細胞滲透濃度和功能蛋白的數量,而啟動自身保護機制以維持藻細胞的正常代謝,其中當氮、磷濃度為400、25μmol·L-1培養15 d時,藻體可溶性蛋白含量達到峰值,提示海水中氮、磷濃度超過該值后,進入藻體的氮、磷不能參與蛋白的合成和生長。
3.4 氮、磷濃度對海黍子抗氧化能力的影響
藻類在生長過程中,活性氧的產生與清除處于動態平衡狀態。當藻類處于逆境脅迫下,動態平衡被破壞,造成活性氧自由基的大量累積,由此啟動抗氧化酶類防御系統。SOD、CAT和POD是藻類細胞內清除活性氧的主要抗氧化酶,其活性被認為是藻類抗逆性的重要指標[17-18]。本研究表明,氮、磷濃度和培養時間可顯著影響海黍子藻體的抗氧化酶系統。氮、磷濃度為50、3.13 μmol·L-1加富培養3 d時,CAT和POD活性沒有顯著變化,而SOD活性略有下降,表明該處理沒有造成藻細胞活性氧的積累。當氮、磷濃度升高至200、12.5μmol·L-1以上時,SOD活性顯著升高,而CAT的活性則在氮、磷濃度為600、37.5 μmol·L-1的試驗組中顯著降低,推測發生此種變化的原因可能是藻體受脅迫后產生活性氧(ROS)的量上升,從而激活更多的SOD來清除活性氧,而產生的超氧化物進一步歧化為H2O2,H2O2的大量積累導致CAT代謝失衡,活性下降。培養7 d后,SOD、CAT和POD 3種抗氧化酶活性變化各異,表明海黍子對海水中不同氮、磷營養條件的響應是一個復雜過程。對于作為衡量細胞膜脂損傷程度重要指標的MDA含量,氮、磷濃度在低于200、12.5μmol·L-1的試驗組中培養3 d時出現了短暫的下降,進一步說明藻體內活性氧可能沒有大量積累,膜脂過氧化程度較低,而持續培養7 d后,各氮磷加富試驗組藻體MDA含量較之對照組均明顯升高,表明藻體內活性氧增多,造成膜脂過氧化程度增強。這一現象說明,長期富營養化環境會使海黍子的抗逆性受到抑制,不但不能起到生物修復作用,反而受其脅迫,海黍子生長受到抑制。
4 小結
本研究探討了不同氮、磷營養條件下海黍子的生長、生理生化特性以及抗氧化能力的變化情況,表明氮、磷濃度在一定濃度范圍和培養時間內對海黍子生長起到明顯的促進作用,而當氮、磷濃度超過一定值或培養時間時,海黍子的生長及藻體的各項生理生化指標開始受到抑制,并逐漸發生過氧化損傷,抗逆性受到抑制,該結果可為進一步完善海黍子的健康養殖技術和對富營養化污染水域的修復機制研究提供一定的理論參考。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
