氮添加和放牧對(duì)云霧山草原優(yōu)勢(shì)種異速生長(zhǎng)模式的影響
2021-10-19 13:41:12王淑紅晏昕輝
草地學(xué)報(bào) 2021年9期
張 博, 程 杰, 王淑紅, 劉 晶, 楊 雪, 晏昕輝, 李 偉
(1.中國(guó)科學(xué)院水利部水土保持研究所, 陜西 楊凌 712100; 2.中國(guó)科學(xué)院大學(xué), 北京 100049; 3.國(guó)家林業(yè)和草原局西北調(diào)查規(guī)劃設(shè)計(jì)院, 旱區(qū)生態(tài)水文與災(zāi)害防治國(guó)家林業(yè)局重點(diǎn)實(shí)驗(yàn)室, 陜西 西安 710000; 4.寧夏云霧山國(guó)家級(jí)自然保護(hù)區(qū)管理局, 寧夏 固原 756000; 5.西北農(nóng)林科技大學(xué)草業(yè)與草原學(xué)院, 陜西 楊凌 712100; 6.西北農(nóng)林科技大學(xué)水土保持研究所, 陜西 楊凌 712100)
草地是全球陸地綠色植物資源中面積最大的可再生自然資源,既是發(fā)展畜牧業(yè)的農(nóng)業(yè)資源基礎(chǔ),又是陸地生態(tài)系統(tǒng)中最重要、分布最廣的生態(tài)系統(tǒng)類型之一[1]。大氣氮沉降作為全球變化的主要環(huán)境問(wèn)題之一,已引發(fā)人們的廣泛關(guān)注。自工業(yè)革命以來(lái),隨著工業(yè)用氮的增加和農(nóng)業(yè)生產(chǎn)中氮肥的大量使用,大氣中氮含量迅速增加,導(dǎo)致氮沉降在世界范圍內(nèi)的廣泛發(fā)生[2-3]。持續(xù)的氮沉降對(duì)草地生態(tài)系統(tǒng)的組成、結(jié)構(gòu)和功能產(chǎn)生重要影響[4]。氮素是限制草地生態(tài)系統(tǒng)植物生長(zhǎng)發(fā)育和生產(chǎn)力的主要因素之一,同時(shí)也影響草地植物的光合能力和養(yǎng)分利用效率[5]。放牧作為草地主要利用方式之一,能夠直接或間接地影響植物功能性狀、物種多樣性和生產(chǎn)力[6],但過(guò)度放牧?xí)斐刹莸刂脖缓屯寥绹?yán)重退化。因此,明確草地植物在氮沉降和放牧干擾下的響應(yīng)對(duì)策對(duì)退化草地生態(tài)系統(tǒng)的恢復(fù)和重建具有重要意義。
植物功能性狀是植物在長(zhǎng)期進(jìn)化和發(fā)展過(guò)程中形成的與其定植、存活、生長(zhǎng)和死亡密切相關(guān)的一系列核心植物屬性,這些屬性能夠顯著影響生態(tài)系統(tǒng)的功能和過(guò)程,并能客觀反映植被對(duì)環(huán)境變化的響應(yīng)[7]。近年來(lái),植物功能性狀對(duì)環(huán)境變化響應(yīng)已成為植物學(xué)和生態(tài)學(xué)的研究熱點(diǎn)[8-9]。有研究表明,植物在不同營(yíng)養(yǎng)條件下生物量分配差異的主要原因是植物的異速生長(zhǎng)[10]。異速生長(zhǎng)是指生物體某一特征的相對(duì)生長(zhǎng)速率不等于第二種特征的相對(duì)生長(zhǎng)速率的一種特性[11],這種特性是由物種遺傳性質(zhì)所決定的,能夠影響植物的生物量分配、資源利用率以及植物對(duì)環(huán)境變化的適應(yīng)性[12]。目前傳統(tǒng)的分析方法無(wú)法揭示植物性狀的變化哪些是由環(huán)境變化引起的,哪些是由植株個(gè)體大小變化引起的[13-14]。異速生長(zhǎng)分析能夠排除個(gè)體大小變化的影響,分析植物不同性狀之間的關(guān)系,從而揭示環(huán)境影響植物性狀的變化規(guī)律[15-16]。然而,關(guān)于植物的生物量分配差異產(chǎn)生的具體原因還沒(méi)有定論。陸霞梅[17]認(rèn)為植物形態(tài)可能隨植株大小的變化而變化,物種的一些表型差異,可能僅僅是和植株大小不一或者生長(zhǎng)階段不同有關(guān)。Pigliucci[18]認(rèn)為不同物種的可塑性在不同環(huán)境梯度間也有不同。周小玲等[19]的研究結(jié)果表明物候期的改變也能影響植物的生物量分配模式。由于植物性狀是表征植物適應(yīng)性的重要指標(biāo),在異速生長(zhǎng)研究中,關(guān)于植物繁殖分配對(duì)環(huán)境變化的響應(yīng)已被廣泛研究[20-24],但作為植物重要組成部分的營(yíng)養(yǎng)器官的生物量分配模式對(duì)氮添加和放牧干擾下的響應(yīng)機(jī)制方面的研究還相對(duì)較少[25]。基于此,本文以云霧山草地群落中7種優(yōu)勢(shì)植物為主要研究對(duì)象,探究氮添加和放牧干擾下植物各營(yíng)養(yǎng)器官生物量分配變化,植物各營(yíng)養(yǎng)器官的異速生長(zhǎng)模式與植株個(gè)體大小的關(guān)系,以及氮添加和放牧干擾對(duì)植物各營(yíng)養(yǎng)器官生物量與個(gè)體生物量之間的異速生長(zhǎng)模式的影響。
1 材料與方法
1.1 研究區(qū)域概況
試驗(yàn)地位于寧夏云霧山草原生態(tài)系統(tǒng)國(guó)家長(zhǎng)期科研基地(106°21′~106°27′ E,36°10′~36°17′ N),海拔1 800 ~ 2 180 m,該區(qū)域?qū)儆谥袦貛О敫珊祬^(qū),年平均氣溫為5℃,年均日照時(shí)數(shù)為2 300~2 500 h,年均無(wú)霜期137 d,年均降水量400~500 mm,降水主要集中在6月至9月。植被區(qū)劃屬于暖溫帶典型草原區(qū),其建群種和優(yōu)勢(shì)種為甘青針茅(Stipaprzewalskyi)、大針茅(Stipagrandis)、百里香(Thymusmongolicus)等,伴生種為豬毛蒿(Artemisiascoparis)、星毛委陵菜(Comarumacaulis)等。土壤類型以山地灰褐土和黑壚土為主,pH值7.9~8.2[26]。
1.2 試驗(yàn)設(shè)計(jì)
本試驗(yàn)在天然草地進(jìn)行,采用隨機(jī)試驗(yàn)設(shè)計(jì),包括兩個(gè)放牧處理(封育和放牧)和兩個(gè)氮添加(模擬氮沉降)處理(0和10 g·m-2),共4個(gè)組合,分別為:封育(E),封育×氮添加(E+N),放牧(G),放牧×氮添加(G+N)。每個(gè)組合6個(gè)重復(fù),共24個(gè)小區(qū),每個(gè)小區(qū)的面積(長(zhǎng)×寬)為24 m2(6 m×4 m),在小區(qū)之間設(shè)有2 m的緩沖帶。封育草地自1982年開始,至今封育年限達(dá)38年。放牧草地自1996年開始放牧,放牧期為每年夏秋季6—8月,每月放牧一次,連續(xù)放牧10 d,放牧強(qiáng)度為3只羊單位·hm-2,放牧試驗(yàn)由固定的養(yǎng)殖戶執(zhí)行完成。自2013年起,每年在生長(zhǎng)季節(jié)開始時(shí)(4月底)在小區(qū)中施加尿素10 g·m-2[27],選擇陰雨天氣一次性將肥料撒播于小區(qū)內(nèi),一直持續(xù)至2020年。為了保證試驗(yàn)的一致性并減小誤差,封育和放牧草地各小區(qū)布局和氮添加處理保持一致。
1.3 植被調(diào)查與植株采樣
2020年8月下旬對(duì)每個(gè)小區(qū)選取1 m×1 m的植被樣方進(jìn)行調(diào)查,為了避免邊緣效應(yīng),樣方距邊緣的距離大于50 cm,然后測(cè)定樣方內(nèi)物種數(shù),并記錄每個(gè)物種的高度、蓋度和個(gè)體數(shù)。根據(jù)以往在本地的長(zhǎng)期試驗(yàn)資料和2013-2020年本試驗(yàn)點(diǎn)的群落調(diào)查結(jié)果[28],在長(zhǎng)期封育草地選取下列物種:二裂委陵菜(Potentillabifurca)、甘菊(Chrysanthemumlavandulifolium)、甘青針茅(S.przewalskyi)、白蓮蒿(Artemisiasacrorum);在放牧草地選取下列物種:白蓮蒿(A.sacrorum)、大針茅(S.grandis)、賴草(Leymussecalinus)、百里香(T.mongolicus)。這些物種共同決定了群落80%~90%的蓋度和生物量。因氮添加和放牧改變了群落的物種組成和多度,故每個(gè)處理的優(yōu)勢(shì)物種組成不一定完全相同。
在每個(gè)小區(qū)內(nèi)每個(gè)物種選取3~4株成熟且完整的個(gè)體[29],6個(gè)重復(fù),共20株,根據(jù)植物根系生長(zhǎng)的方向,用鐵鍬在每個(gè)植株的基部10~20 cm的范圍內(nèi)挖取20~30 cm深的包含植物全部或絕大多數(shù)根系的土塊,仔細(xì)地去除附著在目標(biāo)植物根系上的土壤及其他雜質(zhì)[30]。對(duì)甘青針茅(S.przewalskyi)等禾本科克隆植物,因營(yíng)養(yǎng)共享或者競(jìng)爭(zhēng)關(guān)系只發(fā)生在緊密連接的幾個(gè)枝條間[31],故以一個(gè)株叢作為一個(gè)單獨(dú)的個(gè)體。對(duì)于放牧地的植株個(gè)體,選擇家畜采食后已經(jīng)恢復(fù)生長(zhǎng)的完整植株,取樣過(guò)程中保證植株地上部分和地下部分相連,并盡量保證所取根系的完整性。將地下部分裝入封口袋放入冷藏箱中帶回實(shí)驗(yàn)室;將地上部分分為葉、莖及繁殖部分,分別裝入信封,并及時(shí)烘干以防腐爛。在實(shí)驗(yàn)室中用冷水(<5℃)將根系樣品清洗干凈后與地上部分一樣,在65℃條件下烘干至恒重,并記錄數(shù)據(jù)。
1.4 數(shù)據(jù)分析
由于研究對(duì)象為多年生植物,此外在采樣過(guò)程中也發(fā)現(xiàn)繁殖部分相對(duì)較少,故將其歸入到植物的莖部分。個(gè)體生物量(Individual biomass,IB)以根、莖(支持部分)和葉干重的總和表示;植物根、莖和葉的生物量分別以RB(Root biomass),SB(Stem biomass)和LB(Leaf biomass)表示。
異速生長(zhǎng)模型的建立是探索植被生長(zhǎng)策略的數(shù)理統(tǒng)計(jì)方法,具有重要的參考意義。在異速關(guān)系中,兩個(gè)相關(guān)的變量y和x在異速方程中關(guān)系為:
y=cxb
(1)
把上面的方程進(jìn)行對(duì)數(shù)轉(zhuǎn)換就得到:
lgy=lgc+blgx
(2)
分別用Y=lgy,X=lgx和a=lgc替換方程(2),就得到一個(gè)線性關(guān)系:
Y=a+bX
(3)
a是異速生長(zhǎng)指數(shù)(截距),b是異速生長(zhǎng)系數(shù)(斜率)。異速生長(zhǎng)指數(shù)(截距)改變,表明總生物量向該器官的分配效率發(fā)生轉(zhuǎn)變;同時(shí),異速生長(zhǎng)系數(shù)(斜率)改變表明該物種的異速生長(zhǎng)曲線發(fā)生變化。
不同群體之間的異速生長(zhǎng)曲線的變化主要有以下4種類型:第1種是異速生長(zhǎng)系數(shù)(斜率)不同,異速生長(zhǎng)指數(shù)(截距)無(wú)明顯差異(Shift A);第2種是異速生長(zhǎng)系數(shù)(斜率)相同,但異速生長(zhǎng)指數(shù)(截距)不同(Shift B);第3種是異速生長(zhǎng)系數(shù)(斜率)相同,且具備相同的線性擬合軸(Shift C);第4種是異速生長(zhǎng)系數(shù)(斜率)不共軸且異速生長(zhǎng)指數(shù)(截距)也不同(Shift D)[32-33]。
利用獨(dú)立樣本t檢驗(yàn)(Independent-sample t-test)分析氮添加對(duì)封育地和放牧地優(yōu)勢(shì)植物個(gè)體生物量及各營(yíng)養(yǎng)器官生物量的影響。異速生長(zhǎng)方程的參數(shù)估計(jì)采用標(biāo)準(zhǔn)化主軸估計(jì)(SMA)方法[34],比較異速生長(zhǎng)系數(shù)(斜率)是否與1有顯著的不同,當(dāng)斜率b=1時(shí)表示個(gè)體生物量與各營(yíng)養(yǎng)器官之間為等速生長(zhǎng)關(guān)系,否則為異速生長(zhǎng)關(guān)系,該部分采用SMATR Version 2.0計(jì)算完成。同時(shí),利用方差分析(ANOVA)檢驗(yàn)氮添加對(duì)封育地和放牧地優(yōu)勢(shì)種植物異速生長(zhǎng)指數(shù)和異速生長(zhǎng)系數(shù)的影響,并進(jìn)行LSD多重比較,分析異速生長(zhǎng)指數(shù)和異速生長(zhǎng)系數(shù)是否在各處理間有顯著的不同。為滿足正態(tài)分布和方差齊性的要求,對(duì)數(shù)據(jù)進(jìn)行對(duì)數(shù)轉(zhuǎn)換,采用SPSS 22.0分析數(shù)據(jù),Origin 2017作圖。
2 結(jié)果與分析
2.1 氮添加對(duì)封育地和放牧地植物個(gè)體大小變化及各營(yíng)養(yǎng)器官的生物量分配的影響
研究結(jié)果表明,氮添加對(duì)植物各營(yíng)養(yǎng)器官的生物量有顯著影響,對(duì)于封育地的優(yōu)勢(shì)種二裂委陵菜、甘菊、甘青針茅而言,這些物種傾向于向地上分配更多生物量(圖1),具體表現(xiàn)為:二裂委陵菜在氮添加處理下個(gè)體生物量(IB)增加了92.2%,其中莖生物量(SB)和葉生物量(LB)分別增加了134.5%和138.5%(表1),且個(gè)體生物量(IB)、莖生物量(SB)和葉生物量(LB)與封育處理相比均存在顯著差異(P<0.05);甘菊在氮添加處理下植株個(gè)體大小(IB)與封育處理的差異不顯著,但根生物量(RB)葉生物量(LB)存在顯著的差異(P<0.05),其中根生物量(RB)降低了58.0%,葉生物量(LB)增加了68.8%;甘青針茅在氮添加處理下,與封育處理相比個(gè)體生物量的差異不顯著,根生物量(RB)下降了37.6%,其中莖生物量(SB)和葉生物量(LB)略有增加,但差異未達(dá)到顯著水平;白蓮蒿氮添加處理與封育處理相比,葉生物量(LB)有所增加,增加了22.6%。

圖1 封育地和放牧地優(yōu)勢(shì)物種各營(yíng)養(yǎng)器官生物量分配比例
對(duì)于放牧地的優(yōu)勢(shì)物種白蓮蒿、大針茅和賴草來(lái)說(shuō),這些物種傾向于將地上更多的資源分配到地下(圖1)。與放牧處理相比,大針茅在放牧×氮添加處理下,植株個(gè)體生物量(IB)下降了22.7%,植株的莖生物量(SB)和葉生物量(LB)顯著下降(P<0.05),分別降低了55.7%和47.2%;與放牧處理相比,百里香在放牧×施肥處理下,個(gè)體大小(IB)顯著降低(P<0.05),下降了46.8%,莖生物量(SB)和葉生物量(LB)也呈現(xiàn)明顯下降的趨勢(shì),分別下降了49.6%和68.0%;對(duì)于白蓮蒿和賴草這兩個(gè)物種在放牧處理和放牧×氮添加處理下各營(yíng)養(yǎng)器官生物量變化不明顯,均未達(dá)到顯著水平(表1)。
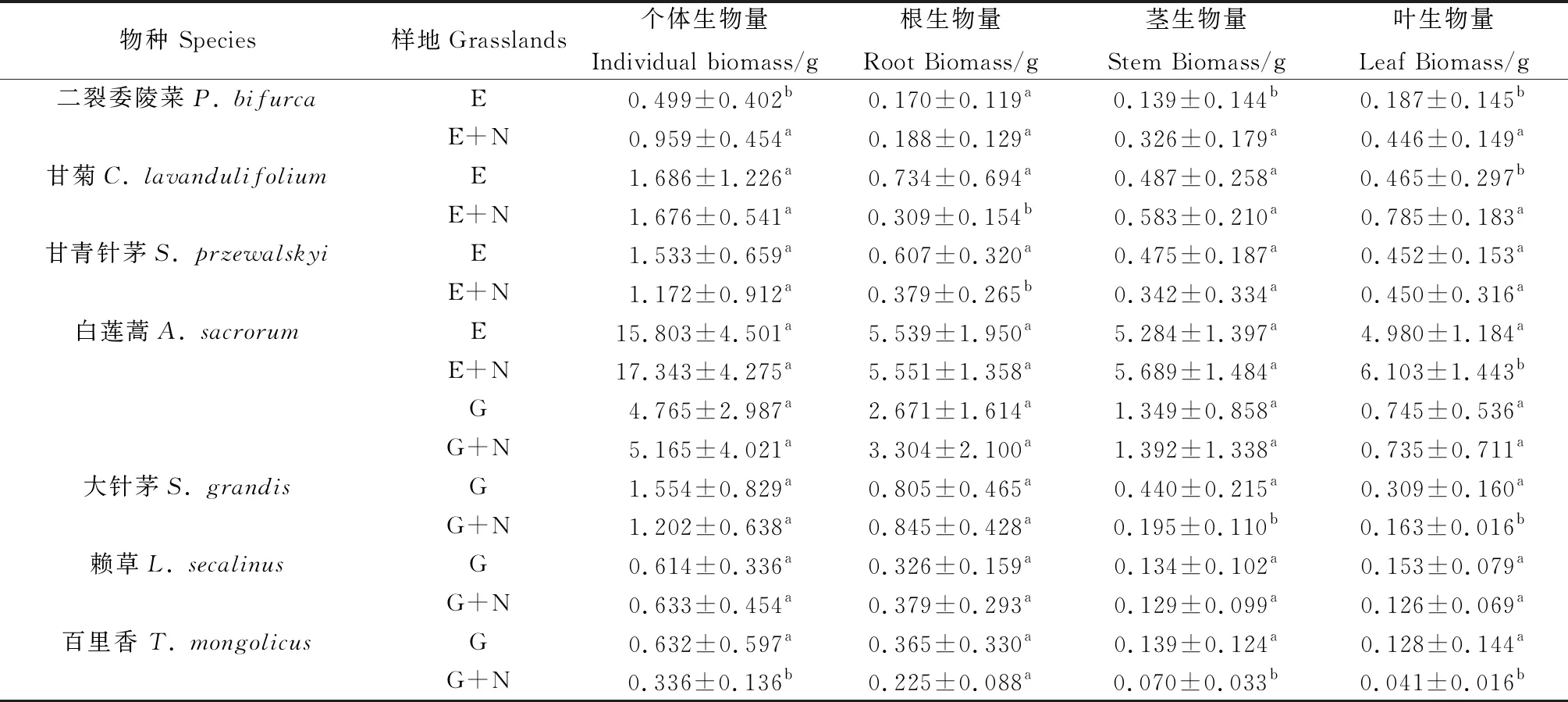
表1 封育地和放牧地優(yōu)勢(shì)種個(gè)體生物量與各營(yíng)養(yǎng)器官生物量
2.2 氮添加對(duì)封育地和放牧地植物各營(yíng)養(yǎng)器官生物量與個(gè)體生物量間異速生長(zhǎng)關(guān)系的影響
對(duì)植物各營(yíng)養(yǎng)器官的生物量與個(gè)體生物量進(jìn)行異速生長(zhǎng)分析表明,對(duì)于IB-RB之間的異速生長(zhǎng)關(guān)系,封育處理(b=0.988,R2=0.990,P>0.05)與放牧處理(b=0.996,R2=0.993,P>0.05)的異速生長(zhǎng)系數(shù)均小于1,但未到達(dá)到顯著水平,故屬于因變量分配速率小于自變量的等速生長(zhǎng)關(guān)系。封育×氮添加處理(b=1.139,R2=0.951,P<0.05)的異速生長(zhǎng)系數(shù)顯著大于1,放牧×氮添加處理下(b=0.977,R2=0.992,P<0.05)的異速生長(zhǎng)系數(shù)顯著小于1,符合異速的相關(guān)生長(zhǎng)關(guān)系;對(duì)于IB-SB之間的異速關(guān)系,4種處理下的異速生長(zhǎng)系數(shù)均顯著大于1(圖2),符合因變量分配速率大于自變量的異速生長(zhǎng)關(guān)系;對(duì)于IB-LB之間的異速生長(zhǎng)關(guān)系,除放牧×氮添加處理下(b=1.025,R2=0.910,P>0.05)的異速生長(zhǎng)系數(shù)大于1,屬于葉生物量(LB)分配速率高于個(gè)體生物量(IB)分配速率的等速生長(zhǎng)關(guān)系外,其余3種處理方式均為異速生長(zhǎng)系數(shù)顯著小于1的異速生長(zhǎng)關(guān)系。

圖2 根(a)、莖(b)和葉(c)生物量與個(gè)體生物量之間的異速生長(zhǎng)關(guān)系
通過(guò)對(duì)不同處理下的植物各營(yíng)養(yǎng)器官生物量與個(gè)體生物量之間的異速生長(zhǎng)指數(shù)(截距)和異速生長(zhǎng)系數(shù)(斜率)進(jìn)行分析,得出了以下試驗(yàn)結(jié)果(圖3):
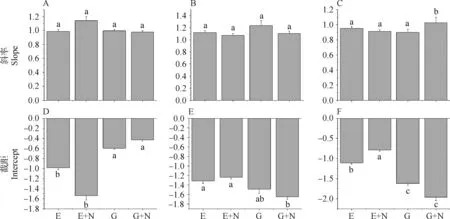
圖3 氮添加對(duì)植物根(A和D)、莖(B和E)和葉(C和F)與個(gè)體生物量之間的異速生長(zhǎng)指數(shù)和異速生長(zhǎng)系數(shù)的影響
1)放牧和放牧×氮添加處理下IB-RB之間的異速生長(zhǎng)指數(shù)(截距)與封育相比均存在顯著性差異,異速生長(zhǎng)系數(shù)(斜率)并未存在顯著性差異(發(fā)生Shift B),而在封育×氮添加處理下,IB-RB之間的異速生長(zhǎng)系數(shù)(斜率)與異速生長(zhǎng)指數(shù)(截距)均未有顯著性差異。
2)對(duì)于IB-SB之間的異速模型,與封育相比,放牧×氮添加處理下的異速生長(zhǎng)曲線的異速生長(zhǎng)指數(shù)(截距)存在顯著性差異,異速生長(zhǎng)系數(shù)(斜率)沒(méi)有明顯變化(發(fā)生Shift B)。封育×氮添加處理和放牧處理下的異速生長(zhǎng)系數(shù)(斜率)和異速生長(zhǎng)指數(shù)(截距)均未有顯著性差異。
3)植物IB-LB之間的異速生長(zhǎng)模型中,與封育相比,封育×氮添加處理下,葉生物量的異速生長(zhǎng)指數(shù)(截距)存在顯著差異,但異速生長(zhǎng)系數(shù)(斜率)均未有顯著變化。放牧處理下的異速生長(zhǎng)指數(shù)(截距)存在顯著性差異,異速生長(zhǎng)系數(shù)(斜率)不存在顯著性差異(發(fā)生Shift B)。而在放牧×氮添加處理下的異速生長(zhǎng)系數(shù)(斜率)和異速生長(zhǎng)指數(shù)(截距)均存在顯著差異(發(fā)生Shift D)。
3 討論
3.1 氮添加和放牧對(duì)植物各營(yíng)養(yǎng)器官生物量分配的影響
根據(jù)最優(yōu)分配理論,植物能在環(huán)境資源限制和干擾條件下將光合作用所固定的能量?jī)?yōu)化分配從而最大化其在多變環(huán)境中的適合度[35]。根據(jù)Müller等[36]的研究,植物在早期生長(zhǎng)階段為了吸收更多的養(yǎng)分會(huì)出現(xiàn)“根化”,而到生長(zhǎng)后期會(huì)出現(xiàn)“莖葉化”。Shipley和Meziane[37]的研究結(jié)果表明,植物在養(yǎng)分含量較低的環(huán)境中,會(huì)分配更多的生物量到植物的根部,而在養(yǎng)分含量較高的環(huán)境下,會(huì)投入更多的資源到植物的莖和葉,以適應(yīng)光競(jìng)爭(zhēng)的增強(qiáng)。就不同處理下植物各營(yíng)養(yǎng)器官的生物量的投入而言,本研究結(jié)果表明,氮添加處理下二裂委陵菜、甘菊和白蓮蒿的葉生物量顯著高于封育處理,而根生物量和莖生物量與封育處理差異并不明顯,這可能是由于植物在養(yǎng)分充足的環(huán)境下,光成為了植物生長(zhǎng)的限制因子,所以植物通過(guò)加大對(duì)葉生物量的分配策略來(lái)提高對(duì)光資源的競(jìng)爭(zhēng)。同時(shí),施氮肥具有明顯的調(diào)水作用,可提高根系的吸水能力,改善葉片的光合能力,有利于提高植物水分利用效率和地上生物量的積累[38]。相對(duì)于封育地,放牧使得氮添加對(duì)植物各營(yíng)養(yǎng)器官生物量分配的影響更為復(fù)雜,Hillebrand等[39]的研究表明,放牧及氮肥的使用對(duì)植物生物量分配的影響主要依賴于群落的均勻度、生態(tài)系統(tǒng)的生產(chǎn)力以及食草動(dòng)物對(duì)植物偏好等眾多因素。而在本研究中,放牧地的優(yōu)勢(shì)物種白蓮蒿、大針茅、賴草和百里香各營(yíng)養(yǎng)器官的生物量分配的趨勢(shì)主要是從地上轉(zhuǎn)移到地下,其中植物莖和葉的生物量的下降最為明顯。這可能與放牧地整體的光環(huán)境有關(guān),放牧后,大個(gè)體的生物被移除,光競(jìng)爭(zhēng)減弱,使得小個(gè)體植物從光競(jìng)爭(zhēng)中釋放出來(lái),投入更多的資源到地下以獲取生長(zhǎng)所需的營(yíng)養(yǎng)。
3.2 氮添加和放牧對(duì)植物異速生長(zhǎng)模式的影響
分配是植物生活史中的一個(gè)重要概念[40],而植物對(duì)于各個(gè)器官或組織的資源分配存在著一個(gè)權(quán)衡,更多的資源分配到某個(gè)器官或組織也就意味著要減少對(duì)其他器官的投入。植物的生物量分配模式?jīng)Q定了植物的優(yōu)先發(fā)展方向,HarPer和Ogden[41]認(rèn)為這種分配策略往往是由物種遺傳決定的或者物種基因型特有的,依賴于植物的個(gè)體大小。Poorte和Nagel[42]認(rèn)為許多植物分配模式的變化是可塑的,環(huán)境變化也可能成為影響植物各器官分配模式的重要因素。植物各器官生物量分配的可塑性通常被認(rèn)為是“表觀的可塑性”或“真實(shí)的可塑性”,這種真實(shí)的可塑性又被定義為植物異速生長(zhǎng)曲線發(fā)生變化,并不只是植物的生長(zhǎng)速率和分配效率發(fā)生了變化[43]。因此,植物各營(yíng)養(yǎng)器官在不同處理下的異速生長(zhǎng)曲線是否發(fā)生改變對(duì)研究植物可塑性對(duì)環(huán)境變化的真實(shí)響應(yīng)具有重要意義。
本研究結(jié)果表明植物各營(yíng)養(yǎng)器官生物量與個(gè)體生物量之間的生長(zhǎng)關(guān)系大部分是異速的(圖2)。Wu等[44]的研究也表明,高山草原植物的生物量分配也符合異速的相關(guān)生長(zhǎng)關(guān)系。Liu等[25]認(rèn)為在區(qū)域尺度上蒿屬植物進(jìn)化出了特有的異速生長(zhǎng)策略。在不同環(huán)境中,各器官之間傾向于保持的特定的異速生長(zhǎng)比例,而不是直接依賴于各器官為響應(yīng)環(huán)境變化而調(diào)整生物量分配。然而Niklas[45]認(rèn)為高寒草原植物生物量分配支持等速分配假說(shuō)而不是異速分配假說(shuō),這也反映了不同的物種獲取資源分配策略在不同的生境中也存在差異。本研究結(jié)果表明,在植物各營(yíng)養(yǎng)器官生物量與個(gè)體生物量之間的異速關(guān)系中,斜率的變化并不明顯,變化主要發(fā)生在截距上。Niu等[46]關(guān)于青藏高寒草原植物繁殖分配與個(gè)體大小之間的異速關(guān)系的研究中,斜率與截距的變化也存在類似的趨勢(shì)。異速生長(zhǎng)曲線截距的變化是常見(jiàn)的,截距的變化說(shuō)明植物的分配效率發(fā)生了改變,并不意味著環(huán)境變化直接影響了植物各營(yíng)養(yǎng)器官的生物量分配策略[47]。本研究結(jié)果表明在植物各營(yíng)養(yǎng)器官生物量和個(gè)體生物量之間的相關(guān)生長(zhǎng)關(guān)系中,只有放牧×氮添加處理下葉生物量和個(gè)體生物量之間的異速生長(zhǎng)斜率發(fā)生了變化,由此也說(shuō)明環(huán)境變化對(duì)植物生物量分配策略的影響相對(duì)較小。在氮添加處理下,封育地植物的競(jìng)爭(zhēng)主要由對(duì)地下土壤養(yǎng)分的競(jìng)爭(zhēng)轉(zhuǎn)變?yōu)閷?duì)地上光資源的競(jìng)爭(zhēng),光競(jìng)爭(zhēng)的增強(qiáng)使得植物分配更多的資源到地上部分;而在放牧地植物地上部分易被動(dòng)物啃食,導(dǎo)致植物出現(xiàn)補(bǔ)償性生長(zhǎng)[48],使得植物地上部分生物量增多。因此,在放牧和氮添加的共同作用下,植物葉分配異速生長(zhǎng)曲線發(fā)生改變可能是因?yàn)橹参镎{(diào)整了原有的分配對(duì)策以更好地適應(yīng)周圍環(huán)境的變化。
4 結(jié)論
總的來(lái)說(shuō),氮添加和放牧對(duì)植物各營(yíng)養(yǎng)器官生物量分配有顯著的影響,氮添加對(duì)封育地植物地上生物量的影響較大,而氮添加對(duì)放牧地植物生物量分配的影響主要集中在植物地下部分。此外,植株各營(yíng)養(yǎng)器官的異速生長(zhǎng)模式依賴于植株個(gè)體大小,且各營(yíng)養(yǎng)器官生物量與個(gè)體大小具有明顯的正相關(guān)關(guān)系。同時(shí),氮添加和放牧對(duì)植物異速生長(zhǎng)模式的影響相對(duì)較小,異速生長(zhǎng)指數(shù)與異速生長(zhǎng)系數(shù)的結(jié)果表明,環(huán)境變化雖然影響了植物各營(yíng)養(yǎng)器官之間的生物量分配效率,但并未直接改變植物各營(yíng)養(yǎng)器官的生物量分配策略。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
鐵道通信信號(hào)(2020年9期)2020-02-06 09:15:22
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
數(shù)學(xué)大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學(xué)科學(xué)(學(xué)生版)(2019年5期)2019-05-21 01:00:18
經(jīng)濟(jì)技術(shù)協(xié)作信息(2018年30期)2018-11-22 06:20:24
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
