Y-box結(jié)合蛋白1通過Gli2調(diào)控肝祖細(xì)胞增殖
2021-10-24 05:38:38李飛李兵航郭悅承沈波顧天翊曲穎張啟迪蔡曉波陸倫根
肝臟 2021年5期
李飛 李兵航 郭悅承 沈波 顧天翊 曲穎 張啟迪 蔡曉波 陸倫根
Y-box結(jié)合蛋白(YB-1)是一種廣泛存在于細(xì)菌和人類的高度保守的轉(zhuǎn)錄因子,能特異性結(jié)合目的基因啟動子和增強子內(nèi)部Y-box序列(CTGATTGGCCAA),發(fā)揮多種重要的生物學(xué)功能,包括胚胎發(fā)育、細(xì)胞增殖、腫瘤的發(fā)生和轉(zhuǎn)移等[1-4]。Didier等[5]發(fā)現(xiàn),YB-1可以與人MHC II基因啟動子區(qū)內(nèi)含有CCAAT序列的順式作用原件Y-box特異性結(jié)合,并抑制MHC II的轉(zhuǎn)錄,發(fā)揮負(fù)調(diào)控轉(zhuǎn)錄因子的作用。現(xiàn)已發(fā)現(xiàn),YB-1可以與多種基因啟動子或增強子結(jié)合,調(diào)控相應(yīng)靶基因的轉(zhuǎn)錄,進而調(diào)控各種生物學(xué)功能[1]。Okamota等[6]發(fā)現(xiàn),YB-1能與p53直接結(jié)合,并且在人表皮癌細(xì)胞中降低YB-1的表達促進p53的轉(zhuǎn)錄。因此,YB-1能通過調(diào)控多種基因啟動子的轉(zhuǎn)錄活性抑制或促進相應(yīng)蛋白的表達,進而調(diào)控細(xì)胞增殖。
研究發(fā)現(xiàn),慢加急性暴發(fā)型肝炎患者的YB-1主要在肝臟膽管細(xì)胞、肝癌細(xì)胞(HPC)細(xì)胞和肝細(xì)胞中高表達。在誘導(dǎo)HPC增殖的小鼠模型中也發(fā)現(xiàn)YB-1主要在肝臟HPC中表達。因此,YB-1可能參與調(diào)控HPC增殖和肝再生。本研究擬在體外細(xì)胞實驗中探索YB-1對HPC增殖的影響及其作用機制。
資料與方法
一、實驗動物與材料
(一)動物 雄性C57BL/6J 小鼠,4周齡,體質(zhì)量20~25 g,購自上海必凱實驗動物中心,飼養(yǎng)于上海市第一人民醫(yī)院SPF級實驗動物中心。
(二)試劑 William′s E培養(yǎng)基、胎牛血清、L-谷氨酰胺、胰蛋白酶購自美國Gibco公司;胰島素樣生長因子-II、表皮生長因子、胰島素購自美國Peprotech公司;Trizol購自日本Takara公司;抗YB-1、抗Gli1、抗Gli2、抗Shh、抗C-myc、抗GAPDH抗體均購自美國Abcam公司;抗Cyclin D1抗體購自美國CST公司;GANT61購自美國Selleck公司。
二、實驗方法
(一)肝祖細(xì)胞分離與培養(yǎng) 4周齡雄性野生型C57BL/6J小鼠予50%膽堿缺乏飼料+0.1%乙硫氨酸喂養(yǎng)3周,制備肝損傷模型,采用膠原酶兩步灌注法分離HPC,分離方法見文獻[7]。采用William′s E完全培養(yǎng)基(含10%胎牛血清、20 ng/mL表皮生長因子、10 μg/mL胰島素和30 ng/mL胰島素樣生長因子-II),在37℃、體積分?jǐn)?shù)為0.05的CO2環(huán)境中培養(yǎng)。
(二)慢病毒載體系統(tǒng)構(gòu)建和細(xì)胞轉(zhuǎn)染 攜帶有特異性干擾YB-1表達的慢病毒載體系統(tǒng)由上海吉瑪生物技術(shù)有限公司構(gòu)建制備。對照組和實驗組干擾序列如下:對照組:5′-TTCTCCGAACGTGTCACGT-3′、shYB-1#1:5′-GAGAGCAAGGTAGACCAGTG-A-3′、shYB-1#2:5′- GTCAAATGGTTCAATGTA-AGG -3′。轉(zhuǎn)染前,用完全培養(yǎng)基重懸肝祖細(xì)胞,調(diào)整密度至2×104cell/mL,取0.5 mL接種于24孔板,共3孔,培養(yǎng)過夜;待細(xì)胞匯合至50%,更換培養(yǎng)基,分別加入20 μL病毒液,繼續(xù)培養(yǎng)24 h,1∶5傳代培養(yǎng)。通常在轉(zhuǎn)染后第9天觀察到Y(jié)B-1干擾組細(xì)胞數(shù)顯著減少,然后收集細(xì)胞提取蛋白和RNA進行后續(xù)實驗。
(三)細(xì)胞增殖檢測 CCK-8實驗:采用細(xì)胞計數(shù)試劑盒(cell counting kit-8,CCK-8)檢測細(xì)胞增殖情況,在96孔板加入100 μL HPC細(xì)胞懸液(2×104/mL),每組6個復(fù)孔,37℃、體積分?jǐn)?shù)為0.05的CO2細(xì)胞培養(yǎng)箱孵育72 h;每孔加入10 μL CCK-8試劑,孵育2 h后應(yīng)用酶標(biāo)儀測定450 nm處的吸光度值(A)。
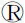
(四)流式細(xì)胞術(shù) 按照美國BD公司細(xì)胞周期檢測試劑盒提供的說明書進行操作,制備單細(xì)胞懸液,PBS洗滌一次,預(yù)冷95%酒精冰上固定過夜,PBS洗滌后加入PI/RNase Staining Buffer 0.5 mL室溫孵育30 min,PBS重懸細(xì)胞后上機檢測。
(五)實時PCR 用Trizol提取總RNA,采用Nanodrop測定RNA濃度。按照Takara公司第一鏈cDNA反轉(zhuǎn)錄試劑盒(RR036A)說明書提供的操作步驟將總RNA反轉(zhuǎn)錄成cDNA。按照日本Takara公司SYBR Green PCR熒光定量PCR試劑盒(RR820)說明操作。PCR條件為:95 ℃變性30 s,55 ℃ 退火30 s,72 ℃延伸30 s,循環(huán)35次 。各靶基因引物序列見表1。

表1 實時PCR 引物序列
(六)高通量測序 采用帶有Oligo(dT) 的磁珠(購自澳大利亞Bioclone公司)捕獲帶有poly A尾的mRNA,將mRNA片段化為100~200 bp大小的核苷酸片段。cDNA純化后,進行PCR擴增、制備文庫。采用HiSeq2500 平臺(美國Illumina公司)進行高通量測序。所得數(shù)據(jù)與小鼠基因組進行比對,對差異表達基因進行生物本體學(xué)、信號通路分析。
(七)染色質(zhì)免疫共沉淀結(jié)合高通量測序 采用Upstate公司提供的試劑盒(目錄號:17-375)進行染色質(zhì)免疫共沉淀,實驗步驟參見試劑說明書。所得染色質(zhì)經(jīng)片段化、末端修復(fù)、3′末端加A、連接、片段選擇及純化、PCR擴增和純化、以及文庫質(zhì)檢等操作后,采用Illumina HiSeq2000平臺(美國Illumina公司)測序,數(shù)據(jù)經(jīng)去污染、去接頭等處理后使用SOAP軟件與Hgl9基因組序列進行比對,并對相關(guān)基因進行GO功能聚類分析KEGG生物通路富集分析。
(八)免疫印跡 采用RIPA裂解液提取胞質(zhì)蛋白,BCA蛋白濃度測定試劑盒確定蛋白濃度。30 μg 細(xì)胞總蛋白與上樣緩沖液按比例混勻,煮沸變性5 min,經(jīng) 10%SDS-PAGE 電泳后,轉(zhuǎn)移至 PVDF 膜上。PVDF 膜以 5%脫脂牛奶室溫封閉 2 h,一抗 4°C孵育過夜。以 TBST 洗滌 3 次,每次5 min,加入HRP 標(biāo)記的二抗(1∶500稀釋),室溫孵育 2 h,用 TBST 洗滌 3 次,每次5 min。用 ECL 化學(xué)發(fā)光。
(九)熒光素酶報告基因試驗 由上海齊合生物公司構(gòu)建含有小鼠Gli2基因啟動子雙熒光素酶報告基因重組質(zhì)粒,將其分別轉(zhuǎn)染至YB-1干擾組和對照組HPC,24 h 后采用雙熒光素酶報告基因檢測系統(tǒng)觀察熒光素酶的表達活性。
結(jié) 果
一、YB-1調(diào)控HPC增殖
首先,通過慢病毒載體系統(tǒng)轉(zhuǎn)染HPC并成功下調(diào)YB-1的表達。CCK-8實驗顯示,沉默YB-1顯著抑制HPC增殖。siYB-1#1和siYB-1#2對HPC中YB-1 mRNA的抑制率分別達到67%和75%;與此相應(yīng),HPC增殖率分別下降了58%和70%。流式細(xì)胞術(shù)檢測細(xì)胞周期顯示,YB-1下調(diào)后G0/G1期細(xì)胞比例增加,表明沉默YB-1能阻滯細(xì)胞周期,進而抑制細(xì)胞增殖,見圖1A-C。
二、YB-1調(diào)控HPC基因表達
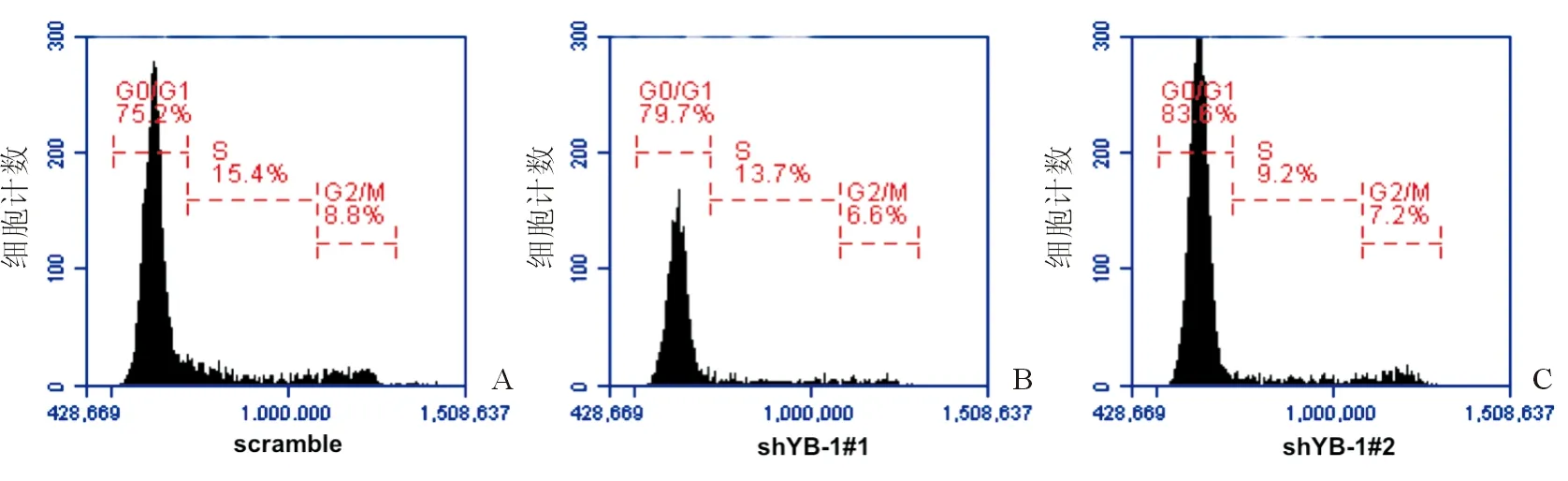
為了進一步研究YB-1調(diào)控HPC增殖的分子機制,RNA-sequence被用來分析YB-1對HPC表達譜的影響。與對照組相比,YB-1沉默組HPC有395個基因表達上調(diào),296個基因表達下調(diào),如圖2A。基因本體學(xué)分析(GO Analysis)表明差異基因參與多種生物學(xué)功能,包括細(xì)胞形態(tài)、細(xì)胞增殖和細(xì)胞粘附等,如圖2B。KEGG分析表明差異基因參與多條重要的信號通路,包括TGF-β信號通路、ECM-receptor interaction、Hedgehog信號通路、Wnt信號通路以及MAPK信號通路,如圖2C。
注:A- C.與scramble組相比,沉默YB-1能將更多肝祖細(xì)胞阻滯在G0/G1期圖1 沉默YB-1抑制肝祖細(xì)胞增殖
注:A.YB-1調(diào)控基因表達;B.基因本體學(xué)(GO)分析;C 信號通路(KEGG)分析圖2 沉默YB-1改變肝祖細(xì)胞基因表達譜
四、YB-1在HPC內(nèi)靶基因分析
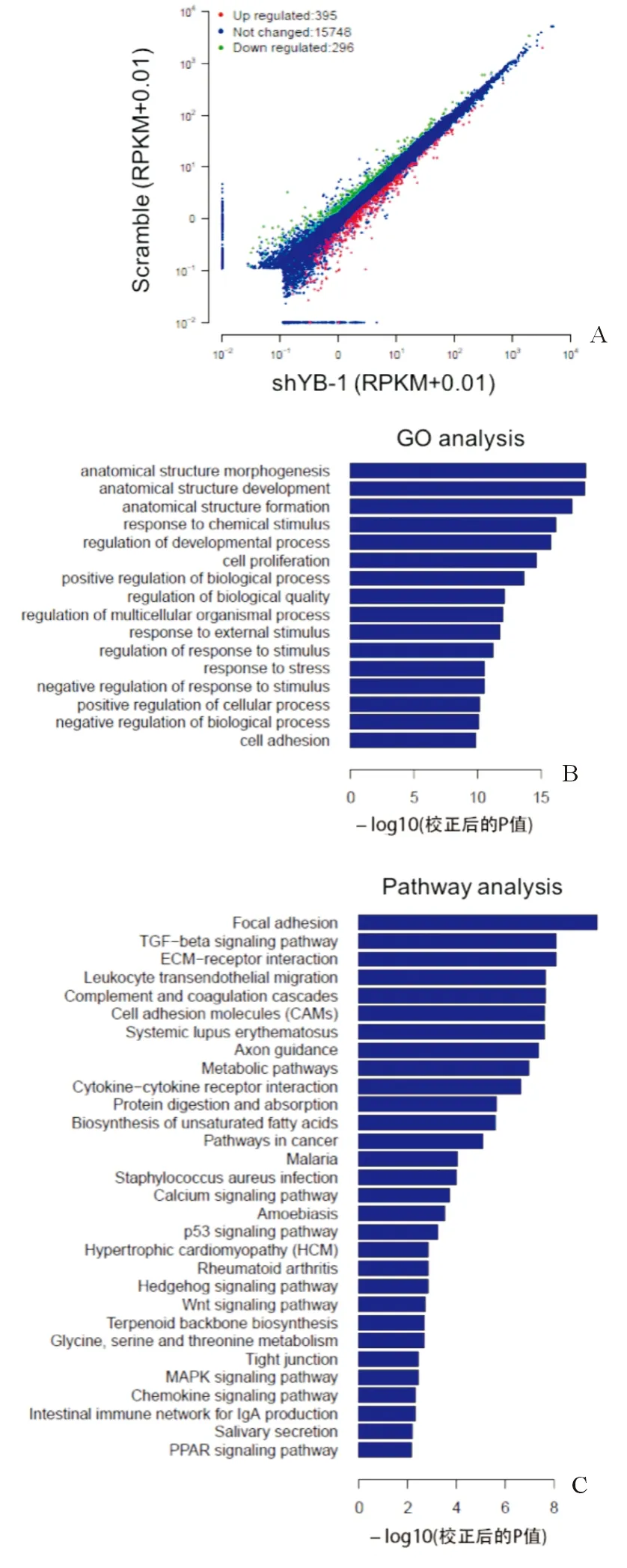
YB-1在HPC全基因組上有8524個結(jié)合位點,其中3609(62%)和1821(31.5%)個結(jié)合位點分別位于間區(qū)和內(nèi)含子區(qū),另外有202(3.4%)和192(3.1%)個分別位于外顯子區(qū)和啟動子區(qū),如圖3A。基因本體學(xué)分析(GO Analysis)表明這些靶基因具有廣泛的生物學(xué)功能,如細(xì)胞形態(tài)、細(xì)胞粘附、信號通路調(diào)控等,如圖3B所示。KEGG分析表明YB-1在HPC中的靶基因參與細(xì)胞粘附、Wnt信號通路、Hedgehog信號通路等,如圖3C。Motif序列研究表明YB-1在HPC中的結(jié)合序列具有多樣性,如圖3D。
注:A.ChIP-sequence表明,YB-1在肝祖細(xì)胞全基因組上共有8524個結(jié)合位點;B.靶基因基因本體學(xué)分析;C.靶基因信號通路分析;D.YB-1在HPC中的結(jié)合序列具有多樣性圖3 YB-1在肝祖細(xì)胞全基因組上的靶基因
五、YB-1調(diào)控Hedgehog信號通路
RNA-sequence和ChIP-sequence均表明YB-1能調(diào)控Hedgehog信號通路,且ChIP-sequence 顯示YB-1在 Gli2啟動子區(qū)有結(jié)合位點。實時PCR表明沉默YB-1能下調(diào)Hedgehog信號通路的配體Shh和Ihh,受體Patched,中間信號分子Gli1和Gli2及其靶基因cyclin D1、cyclin D2、C-myc和N-myc。蛋白質(zhì)印跡法也獲得了相同的結(jié)果,如圖4A。ChIP-PCR表明YB-1能直接結(jié)合Gli2的啟動子區(qū),如圖4B;熒光素酶報告基因?qū)嶒炦M一步表明沉默YB-1能下調(diào)Gli2啟動子的轉(zhuǎn)錄活性,siYB-1#1和siYB-1#2對Gli2基因啟動子雙熒光素酶報告基因重組質(zhì)粒的轉(zhuǎn)錄抑制效率分別達到42%和57%。
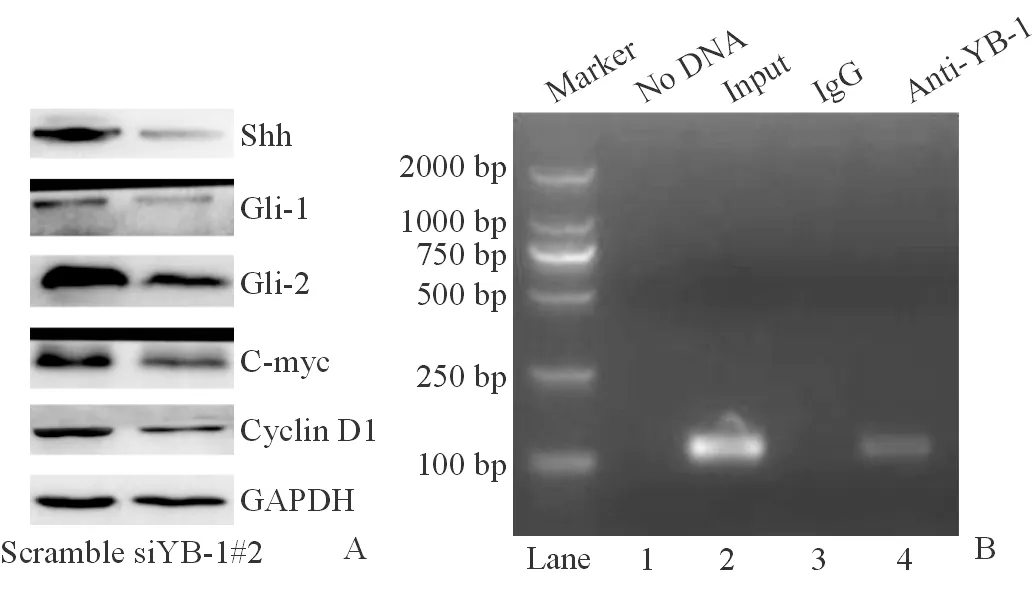
注:A.沉默YB-1抑制Hedgehog信號通路;B.ChIP-PCR表明YB-1能與Gli2啟動子區(qū)結(jié)合圖4 YB-1調(diào)控Hedgehog信號通路
六、Gli1/Gli2阻斷劑抑制肝祖細(xì)胞增殖
為進一步驗證Hedgehog信號通路能調(diào)控HPC增殖,Gli/Gli2抑制劑GANT61被用于阻斷Hedgehog信號通路。CCK-8細(xì)胞增殖試驗表明,GANT61能有效抑制HPC增殖并呈量-效關(guān)系,5 μm GANT61和10 μm GANT61對HPC的抑制效率分別為60%和87%。EdU細(xì)胞增殖試劑盒檢測發(fā)現(xiàn),對照組超過70% HPC 處于有絲分裂期,5 μm GANT61和10 μm GANT61處理組細(xì)胞處于有絲分裂期細(xì)胞數(shù)顯著減少,其比例分別為32%和17%,如圖5A-C。
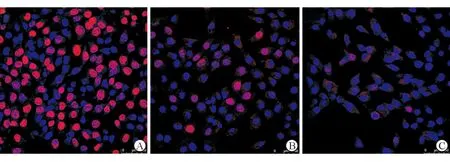
注:A-C.EdU細(xì)胞增殖實驗表明,Gli1/Gli2的抑制劑GANT61處理肝臟細(xì)胞48 h,肝祖細(xì)胞增殖受到明顯抑制,進入有絲分裂期的細(xì)胞顯著減少圖5 Gli1/Gli2阻斷劑抑制肝祖細(xì)胞增殖
討 論
HPC活化和增殖過程較為復(fù)雜,現(xiàn)已發(fā)現(xiàn)多種因素能調(diào)控HPC的生物學(xué)行為。研究表明,腫瘤細(xì)胞因子樣凋亡微弱誘導(dǎo)劑(TWEAK)[8]、肝細(xì)胞生長因子(HGF)[9]、表皮細(xì)胞生長因子(EGF)、成纖維細(xì)胞生長因子-7(FGF-7)、IL-6、TNF-α等細(xì)胞因子或炎癥因子能調(diào)節(jié)HPC的增殖和分化[10]。本研究發(fā)現(xiàn),YB-1能通過Hedgehog信號通路調(diào)控HPC的增殖。既往研究表明,YB-1能通過多種方式調(diào)控細(xì)胞增殖。Homer等[11]也發(fā)現(xiàn)YB-1能抑制p53的轉(zhuǎn)錄,導(dǎo)致細(xì)胞中p53的減少,抑制細(xì)胞凋亡的作用減弱,而腫瘤細(xì)胞增殖加速。Stratford等[12]發(fā)現(xiàn),YB-1能通過促進EGFR的表達而促進乳腺癌的進展。此外YB-1還能促進cyclin D1的轉(zhuǎn)錄調(diào)控非小細(xì)胞肺癌的增殖[13]。本研究通過RNA-sequence和ChIP-sequence首次發(fā)現(xiàn)YB-1能夠調(diào)控Hedgehog信號通路,而該通路被證實廣泛參與各種干/祖細(xì)胞的增殖以及胚胎發(fā)育,并且多項研究表明該通路亦參與肝損傷后肝再生過程[14-16]。同時,ChIP-sequence確定該通路中的關(guān)鍵信號分子Gli2是YB-1的靶基因,并且結(jié)合位點位于啟動子區(qū),因此推測YB-1能通過Hedgehog信號通路調(diào)控HPC增殖。進一步實驗證實,YB-1能與Gli2啟動子區(qū)結(jié)合并正向調(diào)控其轉(zhuǎn)錄,Gli1/Gli2的抑制劑GANT61能抑制HPC增殖,這些結(jié)果均表明YB-1能通過調(diào)控Gli2的表達影響Hedgehog信號通路的強度,進而調(diào)節(jié)HPC的增殖。該研究結(jié)果有利于進一步認(rèn)識肝細(xì)胞和肝再生的調(diào)控機制,能為急性和亞急性肝衰竭開發(fā)新的治療方案提供理論基礎(chǔ)。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年3期)2019-02-01 06:12:26
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(2015年3期)2015-11-11 17:20:00
