CLCOK基因在母牦牛不同組織中的表達
2021-11-02 08:03:42趙海梅靳燕嬰黃榮
青海畜牧獸醫雜志 2021年5期
關鍵詞:檢測
趙海梅,靳燕嬰,黃榮
(1.青海省海東市樂都區畜牧獸醫站2.青海省剛察縣動物疫病控制中心3.青海省畜牧獸醫科學院)
生物體能夠在生理和行為上適應晝夜節律[1]。哺乳動物的晝夜節律系統由分層多時鐘結構組成,從分子、細胞到機體等不同層系上都表現出明顯的時間周期現象[2]。Clock(circadian locomotor output cycles kaput)基因是生物鐘家族重要的核心基因之一,廣泛表達于視交叉上核以及外周組織。牦牛是青藏高原的重要家畜,對于高原的低溫低氧極端環境有很好的適應性。由于這種極端環境的影響,使得牦牛與其他的牛種生理代謝水平有一定的差異。Clock作為動物生物鐘家族的核心成員,可能參與牦牛適應復雜高原環境的代謝調控。因此,通過實時熒光定量PCR的方法,對牦牛不同組織中CLOCK基因表達進行比較研究,為生物鐘基因在調控牦牛生理代謝功能研究中提供理論基礎。
1 材料與方法
1.1 試驗材料
試驗所需的樣品采集于西寧市本地屠宰場。分別采4頭體型一致成年母牦牛下丘腦、心臟、肝臟、脾臟、肺臟、腎臟、卵巢和肌肉等樣品,將采集的新鮮樣品用生理鹽水沖洗去血漬、切成小塊并裝入凍存管后立刻放入液氮速凍,運回實驗室將樣品轉入超低溫冰箱保存備用。
1.2 組織總RNA提取以及cDNA的合成
取0.5g組織樣品在研缽中加入液氮磨碎,使用TransZol Up Plus RNA Kit試劑盒(北京全式金生物技術有限公司)并根據說明書提取組織的總RNA,適量的無RNA酶水溶解。用核酸蛋白儀Nanodrop 2000檢測RNA濃度,同時取6 μL提取的RNA用2%瓊脂糖凝膠檢測RNA的完整性。分裝RNA并置于-80℃冰箱中保存備用。使用PrimeScript 1st Strand cDNA Synthesis Kit試劑盒,根據說明按步驟對提取的總RNA進行反轉錄得到cDNA,得到的cDNA可以進行常規PCR和RT-PCR。
1.3 引物的設計與合成
根據Genebank數據庫中牦牛CLOCK(登錄號:XM_014479222.1)基因和內參基因GAPDH(登錄號:XM_014482068.1)mRNA序列,利用Oligo7.0軟件設計熒光定量引物,并交由南京金斯瑞生物科技有限公司合成。以下是CLOCK和GAPDH的引物序列:CLOCK(F:TAGTAACCGCTCCTGTAGCCTGTG,R:GCTGCTGCTGCGTGACTGAC,產物135bp),GAPDH(F:ACCATCTTCCAGGAGCGAG,R:ATGATGACCCTCTTGGCGC,產物140bp)。
1.4 引物的特異性檢驗
應用常規PCR檢測引物的特異性,反應體系:2×Taq Mix12.5μL,上游引物2μL,下游引物2μL,cDNA 2μL,ddH2O 6.5μL。反應程序:95℃ 預變性5 min,35個循環(95℃ 30 s,60℃ 30s,72℃ 30s),72℃ 10 min。取5 μL擴增產物經2%瓊脂糖凝膠電泳,電壓100 V,時間30 min。
1.5 RT-PCR
分別檢測CLOCK基因在牦牛各組織的相對表達量。反應體系:SYBR Premix Ex Taq(2×)10μL(寶生物工程有限公司),上游引物0.8μL,下游引物0.8μL,Dye 0.4μL,cDNA 2μL,ddH2O 6μL。反應程序:95℃預變性30 s,(95℃變性5 s,60℃退火30s,72℃延伸30 s)共35個循環,72℃ 10 min。每組實驗4個技術重復,4個生物學重復。用2-△△Ct法計算mRNA的相對表達量。
1.6 數據處理
使用SPSS19.0統計軟件進行數據分析,不同時間點的數據用One-Way ANOVA分析。P<0.05為差異有統計學意義。所有數據以“平均數±標準差”表示。
2 結果
2.1 引物特異性
用卵巢cDNA模板分別擴增CLOCK和GAPDH目的片段,并對擴增產物經進行2%瓊脂糖凝膠電泳檢測,在凝膠成像系統中清晰觀察到目的產物在100bp附近,與所設計引物的目的產物CLOCK的135 bp和GAPDH的140 bp大小相一致,而且它們均為單一條帶(圖 1),說明引物特異性高,可用于后續實驗。
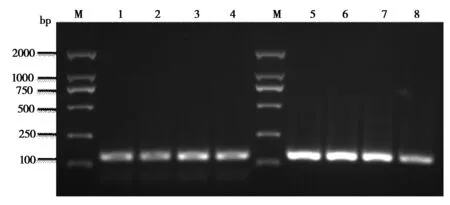
圖1 卵巢CLOCK和GAPDH的擴增結果M:DNA標準;1-4:CLOCK;5-8:GAPDH M:DNA Marker ;1-4:CLOCK;5-8:GAPDH
2.2 RT-PCR檢測目的基因和內參基因的特異性
以制備的卵巢cDNA模板,對CLOCK和GAPDH基因進行RT-PCR,分別得到CLOCK和GAPDH基因的溶解曲線。橫坐標代表溶解溫度(單位:℃);縱坐標為溶解曲線反應的相對熒光讀數(RFU值)即:溫度變化速率/溫度值。顯示溶解曲線峰值單一,PCR產物Tm值相對匯合,表明引物特異性良好,PCR過程中沒有非特異性擴增,排除了實驗過程中出現假陽性的可能。綜上所述,設計的引物可以準確的反應目的基因擴增時的相對表達量。

圖2 牦牛卵巢Clock和GAPDH的溶解曲線
2.3CLOCK基因在牦牛各組織的相對表達量
分別提取母牦牛下丘腦、心臟、肝臟、脾臟、肺臟、腎臟、卵巢和肌肉組織mRNA,利用RT-PCR檢測CLOCK基因表達量,GAPDH為內參基因。結果顯示牦牛CLOCK基因在下丘腦、心臟、肝臟、脾臟、肺臟、腎臟、卵巢以及肌肉中均有表達,且在肺臟和腎臟中高表達,在心臟中的表達量最低(表1,圖3)。CLOCK基因在牦牛不同組織廣泛表達,而且不同組織中含量差異顯著。推測CLOCK基因在牦牛晝夜節律調控中發揮著重要的作用。

表1 牦牛不同組織CLOCK基因mRNA的相對表達量

圖3 牦牛不同組織Clock基因mRNA的相對表達量
3 討論
CLOCK基因在牦牛不同組織中的表達檢測表明,CLOCK基因在下丘腦、心臟、肝臟、脾臟、肺臟、腎臟、卵巢和肌肉8種組織中都有表達,這與其它動物的檢測結果相一致[9,10]。本研究結果表明,在相同實驗條件下,牦牛不同組織內CLOCK基因的表達量存在有很大的區別,其中腎腺中CLOCK的表達量最高,心臟中的表達量最低。
下丘腦是調節晝夜節律的中央時鐘,同時也是調節內分泌的重要樞紐。CLOCK基因在禽類下丘腦—垂體—性腺軸中參與生殖系統發育和繁殖性能的主要調控[11]。通過原位雜交發現,大鼠CLOCK基因主要表達在卵巢顆粒細胞、卵泡膜細胞以及黃體細胞中(Circadian clocks in the ovary)。卵巢的排卵具有自主節律,但是這種節律容易受到黃體激素的影響。我們觀察到牦牛卵巢CLOCK基因表達量相較于除心臟以外的其他組織低,有可能與黃體激素的分泌相關。而有研究發現[12],動物下丘腦中Clock基因的表達會隨著年齡的增加而表達量和節律性出現降低的現象。本研究結果顯示,在牦牛下丘腦中,CLOCK基因的表達量較低,可能是因為實驗樣品采自成年牦牛,其下丘腦的節律性調控作用減弱導致。在Clock基因與代謝功能的研究中,學者們發現敲除了Clock基因的小鼠會導致大量的膽固醇無法發生正常代謝而聚集在肝臟中,說明Clock基因參與肝臟代謝功能的調節[13];心臟的心肌細胞在甘油三酯水平的合成及脂肪分解起著重要作用[14]。在本研究中,牦牛Clock基因在肝臟中的表達量高于心臟,這可能是肝臟和心臟所行使的代謝機能不同所致。作為主要的免疫器官,Clock基因在脾臟存在表達,且表達量較高,這提示了在牦牛中,Clock基因可能參與機體免疫功能的調節。在研究結果中發現,Clock基因在牦牛的肺臟和腎臟都有著較高的表達量,而腎臟中的Clock基因表達量最高,由此可推測Clock基因可能在牦牛的泌尿功能上參與調節。在其它動物上,Clock基因被證實存在于性腺組織中,并參與調控動物的生殖代謝[11]。在本試驗中,我們檢測出牦牛卵巢中存在Clock基因,這為今后研究該基因在牦牛卵巢中激素分泌及卵泡發育功能提供依據。此外,本研究顯示Clock基因在牦牛肌肉的組織中有著較高的表達,揭示Clock基因可能在牦牛調節肌肉節律性活動中具有一定的調節作用。
Clock基因的這種不同組織的相對表達差異也在其它物種中表現:Liu[14]等在鼠的研究中發現,Clock基因在鼠的肝臟的表達量高于脾臟的表達量;而在雞的組織中,胸肌的表達量最高,腎臟、心臟、下丘腦中度表達,肝臟、腸道、卵巢低度表達[9]。實驗的結果也顯示,不同牦牛組織中,Clock基因的表達量存在一定的差異,這可能取決于基因在不同組織中所參與的調控功能的差異有關,具體功能機制還需進一步研究。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
