基于宏基因組測序的嘎日迪-5亞急性毒性機理初探
2021-11-03 02:00:32烏日漢麗麗畢力格陳玉花肖田梅孟香花寶樂爾朝魯蒙格日勒白梅榮
中醫藥信息 2021年10期
烏日漢,麗麗,畢力格,陳玉花,肖田梅,孟香花,寶樂爾,朝魯蒙格日勒,白梅榮*
(1.內蒙古民族大學蒙醫藥研發工程教育部重點實驗室,內蒙古 通遼 028000;2.內蒙古民族大學化學與材料學院,內蒙古 通遼 028000)
蒙藥嘎日迪-5,又稱五鳳丸,由訶子60 g、制草烏30 g、石菖蒲45 g、麝香7.5 g和木香15 g組成[1]。具有殺黏、止痛、燥協日烏素(黃水)、消腫的功效,主治黏刺痛、黏疫、白喉、丹毒、炭疽、赫如虎、吾亞曼、陶賴、協日烏素癥、腫脹等證候。是沿用至今的消黏、燥黃水病的含有毒藥味草烏的地域性知名復方制劑[2]。臨床上由于個體差異或不合理應用含草烏藥物可出現神志不清、脈緩、呼吸困難等一系列中毒現象[3]。故嘎日迪-5毒性是使廣大醫患關注的熱點問題之一。目前有關于嘎日迪-5抗炎鎮痛的研究報道[4],但尚未見其嘎日迪-5 相關現代毒理學研究。
宏基因組學規避了不可分離培養的微生物,應用基因組學技術更真實的反應樣本中微生物組成、互作情況;直接分析微生物中所含全部基因組信息并在分子水平挖掘其代謝通路、基因功能的一種現代藥理和毒理學研究手段[5-6]。從腸道宏基因組學角度切入,對揭示藥物多因素、多途徑、多基因參與的中毒機制和評價藥物安全性均具有非常重要的實際意義。本研究通過宏基因組測序技術分析嘎日迪-5 對腸道菌群的影響,以期探討嘎日迪-5 亞急性毒性作用和機制,并為其臨床安全合理應用提供工作基礎。
1 實驗材料
1.1 動物
SD雄性大鼠,體質量(200±20)g,由遼寧長生生物技術股份有限公司提供[SCXK(遼)2015-0001]。均飼養在SPF級動物室內,并24 h維持室溫為(23±1)℃。
1.2 藥品與試劑
嘎日迪-5 由內蒙古民族大學附屬醫院蒙藥制劑室提供;谷丙轉氨酶(ALT)、谷草轉氨酶(AST)、肌酸激酶(CK)、肌酸激酶同工酶(CKMB)、尿素氮(UREA)、肌酐(CREP)、堿性磷酸酶(ALP)試劑盒均購自羅氏診斷產品(上海)有限公司。
1.3 儀器
羅氏全自動生化分析儀(cobas c 311,上海羅氏診斷產品有限公司);核酸電泳儀(DYCP-32C 型瓊脂糖水平電泳儀,北京市六一儀器廠);Covaris超聲波破碎儀(Covaris M220,美國Massachusetts);Qubit Fluorometric Quantification(Qubit 2.0,美國Life Technologies);安捷倫Agilent 2100 生物分析儀(Agilent 2100,美國Agilent Technologies Co.Ltd);PCR儀(T100PCR,美國Bio-Rad公司);測序儀(Novaseq 6000,美國Illumina公司)。
2 實驗方法
2.1 動物分組及取材
SD大鼠32只,按體質量隨機分為正常組(Z)、嘎日迪-5高劑量組(GG)0.342 9 g/kg(臨床等效量4倍)、中劑量組(GZ)0.085 7 g/kg(臨床等效量)、低劑量組(GD)0.021 4 g/kg(臨床等效量1/4倍)等,每組8只。從實驗第1天開始,各給藥組灌胃相應的藥物溶液,每日灌胃1次,連續灌胃28 d,正常組灌胃等容量生理鹽水。末次給藥后,將大鼠肛周消毒后,通過懸尾刺激法無菌采集糞便樣品置-80 ℃保存,待測糞便DNA提取及菌群檢測。麻醉動物,取血,分離血清,待測心肝腎相關生化指標;采集心、肝、腎肝組織固定于10%甲醛,備用于HE染色。
2.2 CTAB法提取糞便樣品DNA
取1 000 μL CTAB 裂解液,加入溶菌酶,將適量的樣品加入裂解液中,充分裂解。離心取上清,加酚∶氯仿∶異戊醇(25∶24∶1)混勻,12 000 rpm 離心10 min 取上清,加氯仿∶異戊醇(24∶1),離心取上清加入異丙醇,-20 ℃沉淀。離心倒出液體,用1 mL 75%乙醇洗滌2 次,晾干,加ddH2O 溶解DNA 樣品,加RNase A 1 μL消化RNA,37 ℃放15 min即可。
2.3 宏基因組建庫測序
2.3.1 樣品檢測
用瓊脂糖凝膠電泳對DNA 樣品的純度和完整性進行檢測。
2.3.2 文庫構建及庫檢
將檢測合格DNA樣品進行打斷,再經末端修復、加A尾、加測序接頭、純化、PCR擴增等步驟完成整個文庫構建。使用Qubit 2.0 進行初步定量,隨后用Agilent 2100 對文庫的插入片段進行檢測,插入片段符合預期后,通過Q-PCR方法對文庫的有效濃度進行定量。
2.3.3 上機測序
文庫檢測合格后,將不同文庫pooling至flowcell,cBOT成簇后使用Illumina PE150高通量測序平臺進行測序。
2.4 數據處理
采用Knead Data 對原始數據進行質控和去宿主。通過Kraken2 和微生物數據庫來鑒別樣本所含物種,再用Bracken 對樣本中物種的實際相對豐度進行注釋。運用MEGAHIT 軟件,將去宿主基因后的clean reads 進行組裝,得到contigs 預測其基因序列,用Cdhit 對得到的基因進行去冗余和定量,使用DIAMOND軟件,注釋去冗余基因相對豐度。
基于物種豐度表和功能豐度表,進行PCoA(主坐標分析)分析來評估物種組成差異。用LEfSe(多級物種差異判別分析)和LDA(線性判別分析)鑒定組間差異物種和功能。使用spearman correlation 分析物種與血清因子、病理指標的相關性。
應用SPSS 22.0 進行統計分析。采用t檢驗、秩和檢驗、單因素方差分析分別進行組間比較。以P<0.05為差異有統計學意義。
3 結果
3.1 嘎日迪-5血液生化指標檢測結果
與正常組比較,嘎日迪-5 高劑量組ALT、UREA、CK、CKMB 含量顯著升高(P<0.05),嘎日迪-5 中、低劑量組UREA、CREP、CKMB、ALP 含量顯著下降(P<0.05),低劑量組AST含量顯著下降(P<0.05)。提示,嘎日迪-5對心、肝、腎臟具有一定的毒性。結果見表1。
表1 嘎日迪-5對大鼠生化指標的影響(±s,n=8)

表1 嘎日迪-5對大鼠生化指標的影響(±s,n=8)
注:與正常組(Z組)比較,**P <0.01,*P <0.05。
ALP(U/L)118.00±26.24 113.14±15.20 77.71±28.61*86.42±32.25*組別Z組GG組GZ組GD組ALT(U/L)34.15±5.04 48.71±14.72*36.34±11.35 31.38±7.13 AST(U/L)63.08±12.85 69.97±22.97 56.92±18.69 51.78±16.84*UREA(mmoL/L)4.22±0.46 5.35±1.23*3.36±0.53**3.18±0.71**CREP(μmol/L)79.83±15.11 83.71±12.86 62.85±8.51*57.33±13.70*CK(U/L)133.40±19.42 226.83±60.36*127.00±64.26 127.83±81.58 CKMB(U/L)263.24±15.19 335.83±63.99*180.93±46.79*194.01±64.24*
3.2 病理檢測結果
3.2.1 肝組織病理檢測
正常組肝小葉結構清晰,肝細胞索和肝血竇大致呈放射狀排列,肝細胞圓潤、飽滿,肝板排列規則、整齊。嘎日迪-5高劑量組肝細胞廣泛水腫;中劑量組個別肝細胞水腫、胞質疏松淡染;低劑量組未見明顯異常。見圖1。

圖1 肝組織病理切片圖(HE染色,×400)
3.2.2 心臟組織病理檢測
正常組心肌纖維排列整齊規則,胞核呈藍色,胞質呈淡粉色,細胞結構清晰。嘎日迪-5高、中劑量組心肌纖維排列紊亂,胞質疏松淡染,部分細胞水腫。低劑量組心肌纖維排列略紊亂,胞質疏松淡染。見圖2。
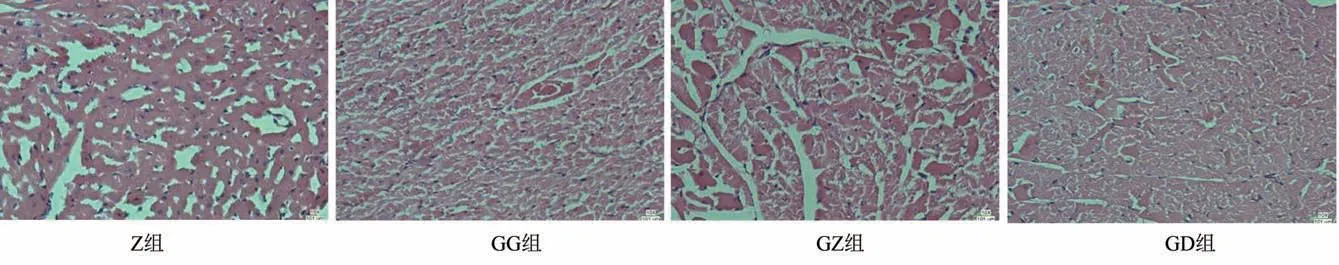
圖2 心組織病理切片圖(HE染色,×400)
3.2.3 腎組織病理檢測
正常組腎小球毛細血管較清晰,腎小管排列緊密,上皮細胞形態正常。嘎日迪-5 高劑量組腎小管上皮細胞有明顯水腫、部分呈空泡化現象;中、低劑量組腎小管上皮細胞略有水腫。見圖3。
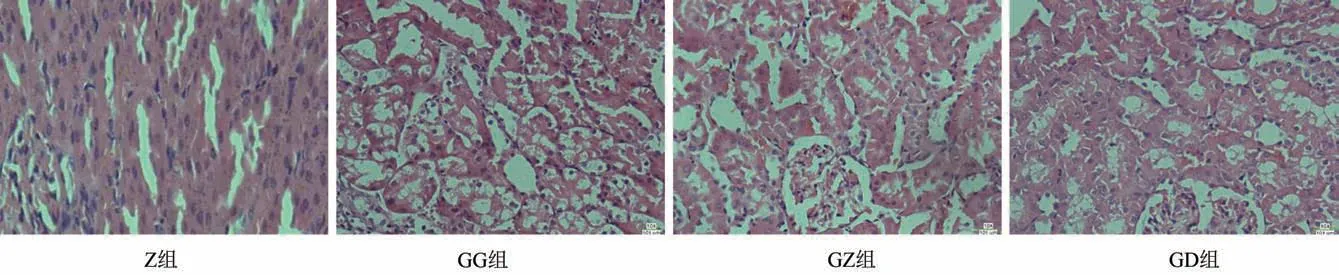
圖3 腎組織病理切片圖(HE染色,×400)
3.3 腸道菌群測序結果
3.3.1 菌群豐度分析
在門、種水平菌群組成注釋結果顯示(圖4、圖5):在門水平,GG 組厚壁菌門(Firmicutes)擬桿菌門(Bacteroidetes)和放線菌門(Actinobacteria)豐度顯著增加,疣微菌門(Verrucomicrobia)、變形菌門(Proteobacteria)豐度下降。
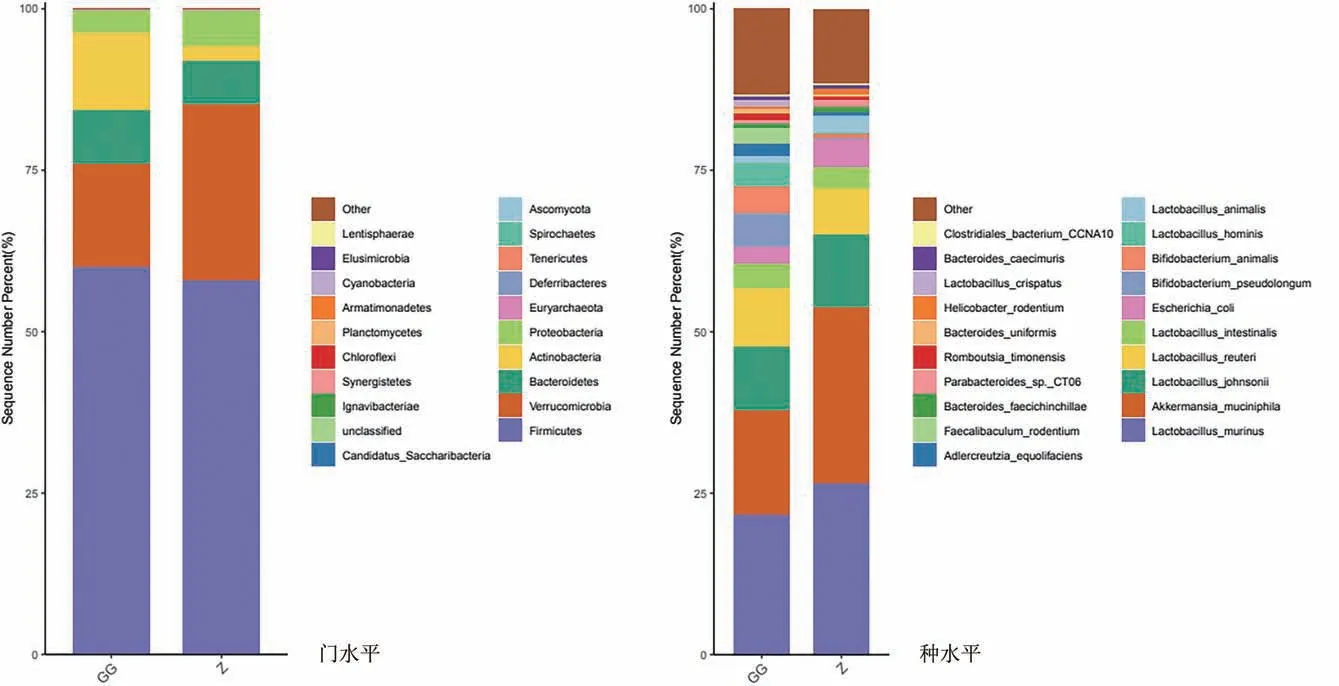
圖4 在門、屬水平菌群柱形圖

圖5 門、屬水平菌群熱圖
在種水平,GG組鼠乳桿菌(Lactobacillus murinus)、嗜黏蛋白艾克曼菌(Akkermansia muciniphila)、約氏乳桿菌(Lactobacillus johnsonii)、大腸桿菌(Escherichia coli)豐度下降,羅伊氏乳桿菌(Lactobacillus reuteri)豐度增加。說明GG明顯改變大鼠腸道菌群結構。
3.3.2 組間差異物種分析
為鑒定兩組間顯著差異物種,使用LEfSe 及LDA(LDA score >2 閾值)分析結果如圖6所示:每橫向柱形代表一個物種,柱形的顏色對應該物種的分組,柱形長度對應LDA 值。Z 組和GG 組間菌群豐度存在明顯的差異。

圖6 LEfSe分析LDA柱形圖
與Z 組相比,假長雙歧桿菌(Bifidobacterium pseudolongum)、動物雙歧桿菌(Bifidobacterium animalis)、Alistipes timonensis、Alistipes finegoldii、北擬桿菌(Bacteroides nordii)、多枝梭菌(Erysipelatoclostridium ramosum) 、Desulfovibrio fairfieldensis、隱 藏 梭 菌(Clostridium celatum)、大腸桿菌病毒Rtp(Escherichia virus Rtp)、達卡雷梭菌(Clostridium dakarense)、Olsenella uli、脫硫弧菌G11(Desulfovibrio sp G11)、尸毒梭菌(Clostridium cadaveris)等35個菌種明顯在GG組中富集。
為兩組間物種組成的相似性及差異情況,進一步經PCoA 分析結果所示(見圖7):GG組與Z組間樣本未重疊,表示兩組間菌群結構存在一定差異。

圖7 基于Bray Curtis距離矩陣的PCoA 圖
3.4 菌群相關性分析
使用Spearman 相關分析,探討了草烏亞急性毒性與差異菌種的關聯。結果見圖8,其中顏色深淺代表相關系數,橫坐標為血清因子,縱坐標為物種。

圖8 相關性熱圖
血清UREA 與相對豐度升高的生孢梭菌(Clostridium sporogenes)、Alistipes indistinctus、無害梭菌、隱藏梭菌、Alistipes ihumii、Alistipes finegoldii、Alistipes obesi 顯著正相關,與相對豐度降低的嚙齒類幽門螺桿菌(Helicobacter rodentium)負相關。
血清CK、CKMB、ALT、AST與相對豐度升高的豚雙歧桿菌(Bifidobacterium choerinum)顯著正相關,與相對豐度降低的停滯棒桿菌(Corynebacterium stationis)、馬里斯棒狀桿菌(Corynebacterium maris)、解脲寡源桿菌(Oligella ureolytica)、Butyricimonas synergistica顯著負相關。
血清CK、CKMB、ALT 也與相對豐度升高的Desulfovibrio fairfieldensis、假長雙歧桿菌、Faecalibaculum rodentium、糞異桿菌顯著正相關,與相對豐度降低的大腸桿菌負相關。
血清CREP 與相對豐度降低的[Clostridium]glycyrrhizinilyticum 顯著負相關。
血清ALP 與相對豐度降低的生孢梭菌、Alistipes indistinctus 顯著負相關。說明,這些菌種主要參與心肝腎血清因子的調控。
3.5 KEGG功能注釋
基于KEGG數據庫、LEfSe及LDA(LDA score >2)方法進行代謝通路分析,結果見圖9所示:每橫向柱形代表一個通路,柱形的顏色對應相應分組的特征代謝通路ID,柱形的長度對應LDA值。兩組間共鑒定出12個豐度相對較高的代謝通路。GG組顯著富集在蛋白質輸出(map03060)、肽聚糖生物合成(map00550)、牛磺酸和低牛磺酸代謝(map00430)、金黃色葡萄球菌感染(map05150)、群體感應(map02024)、ABC轉運蛋白(map02010)等通路。

圖9 KEGG代謝通路LEfSe分析LDA柱形圖
通過KO(KEGG Orthologous groups)LEfSe 組間差異比較結果顯示,草烏組在蛋白質輸出通路中顯著富集,說明注釋到的功能基因在這通路中占據優勢。如圖10 和圖11所示:蛋白質輸出通路中GG 組yidC、SRP54 特征基因大部分來自約氏乳桿菌、羅伊氏乳桿菌、脆乳桿菌、動物雙歧桿菌、產液阿德勒克羅伊茨菌、其他、未分類菌。其中,yidC 也來源于大腸桿菌、massiliensis 擬桿菌(Bacteroides massiliensis);SRP54也來源于假長雙歧桿菌、陰道乳桿菌(Lactobacillus vaginalis)。

圖10 蛋白質輸出通路
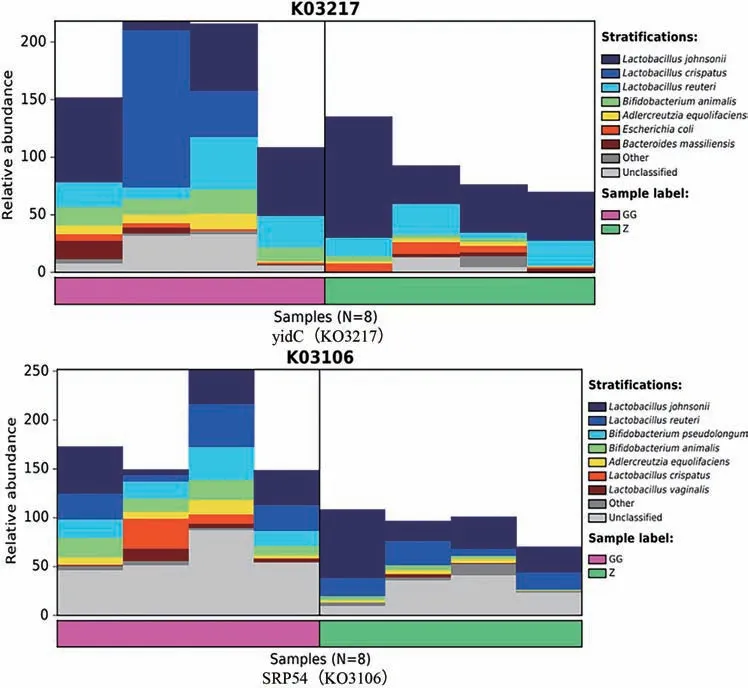
圖11 特征基因分層柱狀圖
4 討論
4.1 血清生化和病理組織學層面對嘎日迪-5 毒性的評價
血清ALT、AST 能靈敏的評估肝細胞的功能狀況。本試驗中,GG 組ALT 與正常組比明顯升高,伴隨肝細胞廣泛水腫;GZ、GD 組ALP 顯著下降(P<0.05);GD組AST 顯著下降(P<0.05)。說明嘎日迪-5 導致大鼠肝細胞損傷和功能障礙。血清UREA、CREP 是反映腎功能變化的常用指標。本試驗中,GG 組UREA 含量增高,腎小管上皮細胞明顯水腫;GZ、GD 組UREA、CREP 顯著下降(P<0.05)。提示嘎日迪-5引起對大鼠腎小球濾過率功能產生損害。血清CK、CKMB 是心肌受損程度的主要指標。本研究發現,GG 組CK、CKMB 顯著升高,心肌纖維排列紊亂、部分細胞水腫、胞質淡染疏松;GZ、GD 組CKMB 顯著下降(P<0.05)。表明嘎日迪-5 對大鼠心肌細胞產生損害。即嘎日迪-5對心、肝、腎臟具有不同程度的毒性。
4.2 差異菌種層面對嘎日迪-5毒性的評價
阿利斯佩斯菌屬包括Alistipes finegoldii、Alistipes ihumii、Alistipes timonensis 等13 種亞型,與腸道炎癥、肝炎、肝性腦病等息息相關[7-9]。多枝梭菌過量增加有助于代謝綜合征和肥胖的發展。大腸桿菌病毒Rtp、達卡雷梭菌、Olsenella uli 與炎癥密切相關[10-11]。尸毒梭菌的顯著富集可增加對腸道、肝臟、腎臟的嗜性,具有嚴重的致病性,也會引起部分感染、膿腫、菌血癥等[12]。Desulfovibrio fairfieldensis 是脫硫弧菌屬一種常見致病性菌,脫硫弧菌在腸道中顯著富集可促進非酒精性肝病、肝纖維化的發生和發展[13-14]。本研究經LEfSe 組間分析結果顯示,GG 組中Alistipes timonensis、Alistipes finegoldii、多枝梭菌、Desulfovibrio fairfieldensis、Alistipes ihumii、隱藏梭菌、大腸桿菌病毒Rtp、達卡雷梭菌、Olsenella uli、脫硫弧菌G11、尸毒梭菌等顯著富集。提示,阿利斯佩斯菌屬、尸毒梭菌、Desulfovibrio fairfieldensis 的顯著富集可能與嘎日迪-5引發肝腎毒性有關。腸道菌群紊亂能加劇炎癥反應,并減弱腸道屏障,促進毒素從腸道位移入血液循環到達肝臟,從而擾亂肝臟代謝和分泌功能[15]。可見,嘎日迪-5 中大腸桿菌病毒Rtp、達卡雷梭菌、Olsenella uli菌的增加可能有助于肝功能的損害。
相關性分析顯示,血清UREA與Alistipes indistinctus、Alistipes ihumii、Alistipes finegoldii、Alistipes obesi顯著正相關。推測嘎日迪-5 腎毒性表達可能與Alistipes 菌屬的增加有關。血清CK、CKMB、ALT與相對豐度升高的Desulfovibrio fairfieldensis 顯著正相關。由此推斷,Desulfovibrio fairfieldensis 的富集可能是嘎日迪-5 產生心臟毒性和肝功能紊亂的主要機制之一。
4.3 蛋白質輸出通路層面對嘎日迪-5毒性的評價
yidC屬于Oxal/Alb3/YidC蛋白家族,細菌中的yidC能輔助其他膜蛋白催化插入到細胞膜,并負責參與大部分SecYEG轉運蛋白在膜上正確地插入及組裝。因此在能量代謝中至關重要。當細菌暴露于細胞表面后,yidC蛋白水平升高,而相應的mRNA水平顯著降低。在藥物誘導下,會導致yidC蛋白的顯著誘導[16-17]。yidC、secD、secA和yajC基因的表達能敏感的通過不同環境因素來影響細菌的黏附[18],從而可能促進毒力因子的產生,致使細胞膜發生病理性變化。本實驗結果表明,蛋白質輸出通路中yidC、secD、secA 和yajC 在GG 組中有表達,其中yidC 是GG組特征基因。SRP54是信號識別蛋白(SRP)的重要組分,是核糖核蛋白復合物的保守成分,能介導蛋白質的共翻譯、靶向和易位至內質網,對各組織的發育非常重要。研究報道,SRP54過表達能增加病毒的復制[19]。SRP54的突變負面影響整個SRP介導的蛋白質分泌途徑,從而導致嚴重的中性粒細胞減少癥,伴胰腺功能不全及骨骼疾病等[20]。在腫瘤相關通路中SRP54 基因的表達具有活性,有望作為膀胱尿路上皮癌(BLCA)的生物標志物[21]。本實驗結果表明,蛋白質輸出通路中SRP54是GG組特征基因。可見,蛋白質輸出通路中高豐度的yidC、secD、secA和SRP54的表達與嘎日迪-5亞急性毒性有關。
綜上,嘎日迪-5以臨床等效量4倍量連續灌胃28 d有一定的心肝腎毒性,即具有一定亞急性毒性;初步認為腸道中阿利斯佩斯菌屬、Desulfovibrio fairfieldensis的顯著富集是嘎日迪-5產生心臟毒性和肝腎功能紊亂的主要機制之一;其亞急性毒性與蛋白質輸出通路相關。該研究為進一步采用敲除yidC、secD、secA和SRP54基因法驗證其嘎日迪-5肝腎心毒性研究奠定了工作基礎。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
