硝磺草酮抗性菌株的篩選及抗性hppd基因的克隆表達
2021-11-04 07:53:34張磊李林
中國農業大學學報 2021年9期
關鍵詞:除草劑
張 磊 李 林
(1.西華師范大學 生命科學學院,四川 南充 637000;(2.南京振旭生物科技有限公司,南京 210000)
草甘膦和抗草甘膦作物的組合在農作物轉基因領域超過20年的大面積持續應用,對全球范圍農田雜草的控制起到了非常大的作用[1]。然而,長期高劑量使用草甘膦造成了嚴重的環境危害和巨大的選擇壓力,已經導致了40多種雜草對草甘膦產生了抗藥性[2-3]。草甘膦及其抗性基因替代組合的發掘和應用,是植物轉基因領域重要且緊迫的研究任務之一。
4-羥基苯丙酮酸雙加氧酶(4-hydroxyphenylpyruvate dioxygenase,HPPD,EC1.13.11.27)抑制劑類除草劑是近年來新開發的一類新型白化型除草劑,其除草機理是通過競爭性抑制植物酪氨酸代謝途徑中HPPD的活性,從而使質體醌和生育酚的合成受阻,間接影響了類胡蘿卜素的合成和功能,使植物光合作用不能正常進行,導致雜草葉片白化,組織壞死,最終死亡[3-5]。HPPD抑制劑類除草劑廣泛應用于防治玉米田中的闊葉雜草和禾本科雜草,具有環境安全性高、作物安全性好和雜草抗性產生慢等優點,被認為是替代草甘膦的轉基因作物靶標除草劑[4-5]。HPPD抑制劑類除草劑主要品種有硝磺草酮、苯吡唑草酮和異噁唑草酮,其中硝磺草酮是用量最大的一類。目前,巴斯夫和先正達利用來自于燕麥的HPPD突變體AvHPPD-03開發出抗硝磺草酮的轉基因大豆SYHT0H2并已進行了商業化生產[6-9],而國內僅克隆出了3個HPPD基因[10-12],尚無高抗硝磺草酮的HPPD編碼基因的報道。
自然界微生物庫中存在廣泛的各類基因資源,本研究從除草劑污染的土壤中分離篩選到一株硝磺草酮高耐受菌株MST-5,從中克隆到硝磺草酮抗性基因Sthppd,利用E.coliBL21(DE3)對StHPPD進行了異源表達和純化,研究了StHPPD對硝磺草酮的抗性,旨在為新型除草劑抗性基因的發掘和應用提供基礎材料。
1 材料與方法
1.1 材料和培養基
L-酪氨酸購自百靈威公司(中國,上海),分子生物學試劑購自大連寶生物公司和南京諾維贊生物科技有限公司,引物及測序服務委托上海生工生物公司。其他化學試劑(分析純)購于國藥集團化學試劑有限公司。硝磺草酮、苯吡唑草酮和二酮腈(異噁唑草酮代謝物)購買自美國Sigma-Aldrich公司,分別用DMSO配置成300 mmol/L的母液,過濾除菌后保存于4 ℃冰箱。
LB培養基:NaCl 5.0 g/L,蛋白胨10.0 g/L,酵母膏5.0 g/L。基礎鹽培養基(MSM):NaCl 1.0 g/L,NH4Cl 1.0 g/L,KH2PO40.5 g/L,K2HPO41.5 g/L,MgSO4·7H2O 0.2 g/L,加超純水至1 L,并調節pH至7.0。酪氨酸無機鹽培養基(TMSM)為MSM培養基添加1 g/L的酪氨酸。
1.2 抗性菌株的篩選分離
土壤樣品采自于南京某農藥廠和安徽某農藥廠表層土壤和下水道污泥。稱取2.0 g樣品加入到添加有2 000 μmol/L硝磺草酮的50 mL MSM培養基中,在30 ℃,180 r/m富集培養5 d。然后,將5 mL 培養物轉移到50 mL新鮮的MSM培養基中,繼續培養5 d。經過連續富集馴化5輪后,吸取1 mL上清培養液并稀釋涂布于TMSM平板上,挑取在平板上生長的單菌落反復劃線,獲得純培養物,并經革蘭氏染色法篩選獲得其中的陰性菌。純化得到的革蘭氏陰性菌株轉接至含有500 μmol/L硝磺草酮的TMSM平板上,挑取能在平板上生長的菌株,然后轉接到新的篩選平板上,轉接過程中逐漸增加篩選平板上硝磺草酮的濃度以增加選擇壓力,最終篩選出硝磺草酮高度耐受菌株。
1.3 抗性菌株對硝磺草酮的抗性測定
為比較抗性菌株對硝磺草酮的抗性情況,將篩選得到的抗性菌株轉接至TMSM液體培養基中,在30 ℃,180 r/m條件下培養至對數生長期,5 000 r/m離心收集菌體,用TMSM液體培養基重懸菌體并調節OD600為10.0,以1%的接種量分別接種到含500、1 000、2 000、3 000、4 000和5 000 μmol/L硝磺草酮的20 mL TMSM液體培養基中,設置不接抗性菌株作為對照。于37 ℃、180 r/m培養60 h后檢測OD600值。
1.4 抗性菌株的16S rRNA基因序列分析及系統進化樹構建
采用SDS-高鹽法提取抗性菌株基因組DNA,根據16S rRNA基因通用引物27F/1492R(表1),PCR擴增抗性菌株的16S rRNA基因序列。PCR產物TA克隆至pMD18-T載體,轉化至EscherichiacoliDH5α。提取陽性克隆子質粒并委托上海生工進行測序。得到抗性菌株的16S rRNA基因序列提交到在線網站EzBioCloud(https:∥www.ezbiocloud.net/)中進行序列比對。下載同源性高的模式菌株的16S rRNA基因序列,在MEGA 6.0軟件中,利用鄰接法構建進化樹。
1.5 硝磺草酮抗性基因hppd的克隆
根據同屬的StenotrophomonasindicatrixWS40T的hppd基因(Locus tag: CR918_RS17620)序列,設計引物對Sthppd-F和Sthppd-R(表1),以菌株MST-5基因組DNA為模板,PCR擴增Sthppd基因,并克隆到pMD18-T載體上,然后轉化至E.coliDH5α中。提取陽性克隆子的質粒并對Sthppd基因進行測序。通過NCBI數據庫中BLASTN和BLASTP對獲得的Sthppd基因序列進行核酸和蛋白序列的比對分析。
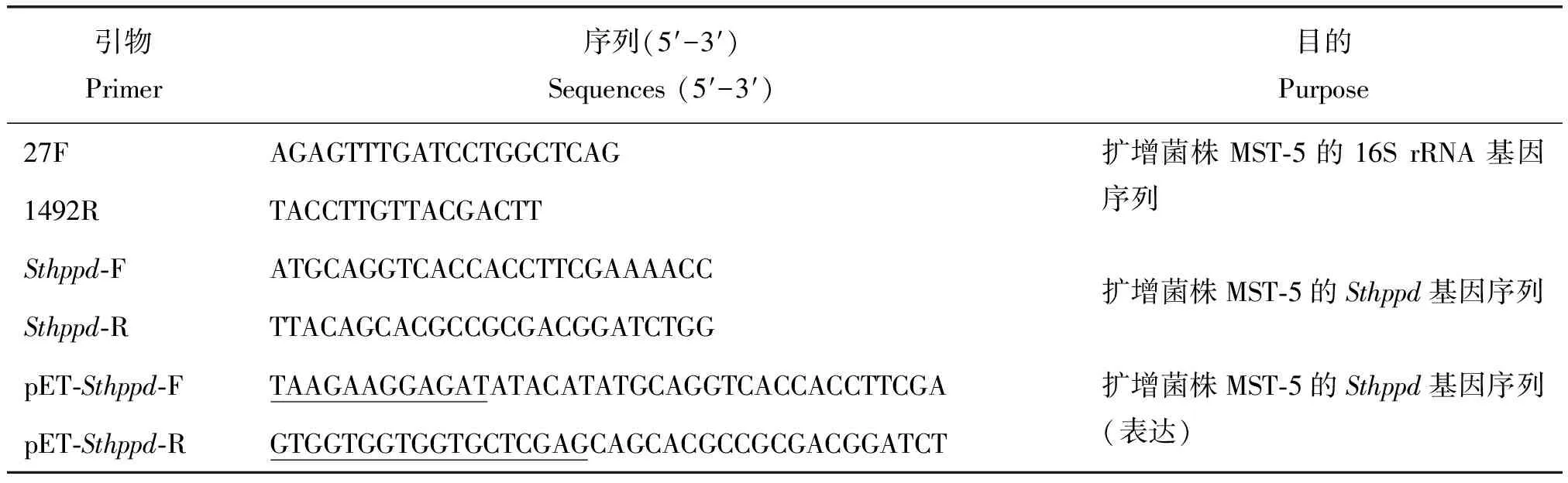
表1 引物序列Table 1 Primers used in this study
1.6 HPPD蛋白表達和純化
以MST-5菌株基因組DNA為模板,根據引物對pET-Sthppd-F和pET-Sthppd-R,PCR擴增其hppd基因。對PCR產物進行回收純化,然后利用同源重組的方式克隆至線性化的pET-29a(+)載體。構建好的pET-Sthppd表達載體轉化至E.coliBL21(DE3)感受態細胞中。StHPPD重組蛋白在16 ℃、160 r/m搖床中過夜誘導表達,利用Co2+柱進行蛋白純化。具體方法參考彭乾等[13]。
1.7 HPPD酶活檢測
在50 mmol/L PBS(pH 7.4)緩沖溶液中添加2 mmol/L 抗壞血酸,1 mmol/L FeSO4,0.2 mmol/L 4-HPP和25 μgStHPPD,構建總體積為1 mL的酶促反應體系,以不添加酶作為對照。將酶促反應體系置于30 ℃反應15 min后,沸水浴5 min終止反應。樣品經離心過濾后,利用HPLC檢測尿黑酸的生成量,HPLC檢測方法見1.10。
1.8 HPPD抗性測定
在1.7構建的標準酶反應體系中添加0~50 μmol/L 的硝磺草酮、苯吡唑草酮和二酮腈,于30 ℃反應15 min后沸水浴5 min終止反應,利用HPLC檢測尿黑酸的生成量。以不添加HPPD抑制劑時的酶活為100%,計算相對酶活。在GraphPad Prism 6軟件中利用非線性擬合計算不同抑制劑的半抑制濃度(IC50)值。
1.9 HPPD酶學性質分析
將1.7構建的酶促反應體系置于10~50 ℃條件下測定StHPPD的最適反應溫度,以30 ℃時的酶活為100%,計算相對酶活。配置3種50 mmol/L緩沖溶液,在pH 4.0至9.0范圍內分析StHPPD的最適pH:檸檬酸緩沖液(pH 3.0~6.0)、PBS緩沖液(pH 6.0~8.0)和Tris-HCl緩沖液(pH 7.0~9.0)。以在50 mmol/L PBS緩沖液中pH 7.0時的酶活為100%,計算相對酶活。在酶促反應體系中分別加入1 mmol/L不同的金屬離子(Fe2+、Fe3+、Mg2+、Ca2+、Al3+、Co2+、Mn2+、Hg2+、Cu2+、Ni2+、Zn2+和Ba2+),5 mmol/L Urea,5 mmol/L 金屬螯合劑EDTA,5 mmol/L SDS和0.1%的DEPC,測定它們對酶活的影響,以添加Fe2+時的酶活為100%。
1.10 HPLC檢測方法
4-HPP、HGA和硝磺草酮的HPLC檢測方法參照彭乾等[13]。其中4-HPP和HGA的流動相為V(乙腈)∶V(水)=3∶7,水中添加體積分數為0.1%的三氟乙酸;硝磺草酮的流動相為V(甲醇)∶V(水)=55∶45,水中添加0.1%的磷酸;流速1.0 mL/min,柱溫40 ℃;4-HPP、HGA和硝磺草酮的檢測波長分別為282、292和245 nm。
2 結果與分析
2.1 硝磺草酮抗性菌株的篩選
經分離純化,從采集的土壤樣品中共獲得了96株能以酪氨酸為唯一碳源生長的細菌,分別編號為MST-1~MST-96。為從中篩選硝磺草酮抗性菌株,將96株菌分別逐漸轉接至含有500~5 000 μmol/L硝磺草酮的TMSM平板上。在低濃度硝磺草酮(500 μmol/L)作為選擇壓力時,大部分細菌的生長不受抑制,從中篩選獲得了72株菌。然而,隨著硝磺草酮濃度的不斷提高,細菌的生長受到抑制,獲得的菌株也越來越少。在含有5 000 μmol/L硝磺草酮的TMSM平板上篩選到3株菌株,分別為MST-5、MST-32和MST-66。
為進一步比較3株菌株對硝磺草酮的抗性,將它們分別接種于含有500~5 000 μmol/L硝磺草酮的TMSM液體培養基中,37 ℃培養60 h后測定培養液的OD600,結果如表2所示。在含有500 μmol/L硝磺草酮的TMSM液體培養基中,菌株MST-5和MST-32的生長基本不受影響;然而,隨著硝磺草酮濃度的增加,菌株MST-5和MST-32的生長逐漸被抑制;相比較而言,菌株MST-5對硝磺草酮的抗性要稍好于MST-32。對于菌株MST-66,在含有500~5 000 μmol/L硝磺草酮條件下,其生長不受抑制。因此,3株菌株對硝磺草酮的抗性強度為:MST-66>MST-5>MST-32。
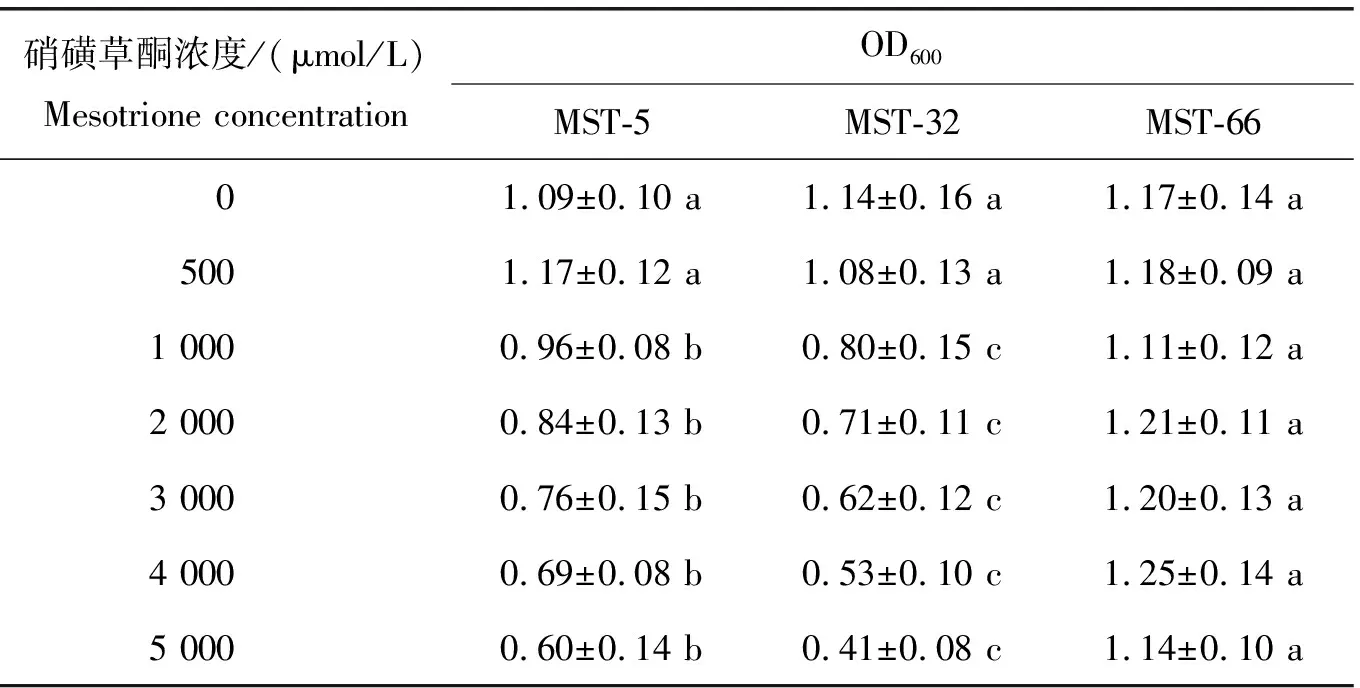
表2 菌株MST-5、MST-32和MST-66對不同濃度硝磺草酮的抗性Table 2 The resistance of strains MST-5, MST-32 and MST-66 to mesotrione
據文獻報道,細菌對HPPD抑制劑的抗性具有多種機制,而最常見的是體內靶標酶HPPD對除草劑不敏感以及降解酶將除草劑降解而獲得的抗性[18]。為研究菌株MST-5、MST-32和MST-66對硝磺草酮的抗性是來自靶標酶抗性還是降解獲得的抗性,選取上述添加了500 μmol/L硝磺草酮的TMSM液體培養基,利用HPLC對培養基中的硝磺草酮含量進行檢測。如圖1所示,硝磺草酮的出峰時間在5.9 min左右,處理(接種菌株MST-5、MST-32和MST-66)和對照(未接菌)中的硝磺草酮特征峰相互疊加,且峰面積基本一致,也沒有新的特征吸收峰的生成,表明硝磺草酮沒有被降解。因此菌株MST-5、MST-32和MST-66對硝磺草酮的抗性可能是來自靶標酶抗性而非降解獲得的抗性。
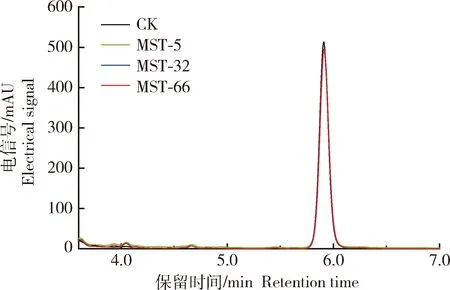
圖1 菌株MST-5、MST-32和MST-66對硝磺草酮的降解能力Fig.1 HPLC analysis of the degradation of mesotrione by strains MST-5, MST-32 and MST-66
2.2 菌株MST-5的系統進化分析
經16S rRNA基因序列分析,菌株MST-5、MST-32和MST-66分別為嗜麥芽窄食單胞菌屬(Stenotrophomonassp.)、伯克氏菌屬(Burkholderiasp.)和節桿菌屬(Arthrobactersp.)。鑒于節桿菌中沒有HPPD基因,其通過3,4-二羥基苯乙酸途徑(Homoprotocatechuate pathway)代謝酪氨酸[19],因此節桿菌對HPPD抑制劑不敏感。而菌株MST-5對硝磺草酮的抗性要高于菌株MST-32,因此選擇菌株MST-5進行后續研究。
菌株MST-5為革蘭氏陰性、好氧細菌,其在LB平板上生長2 d后,菌落為黃色圓形,邊緣光滑(圖2(a))。用透射電子顯微鏡觀察,MST-5菌體形態為桿狀,大小為0.9~1.0 μm× 2.1~2.2 μm(圖2(b))。PCR擴增得到菌株MST-5的16S rRNA基因序列長為1 441 bp。將其提交至EzBioCloud進行序列比對,結果表明菌株MST-5與StenotrophomonasindicatrixWS40T相似性最高,為99.73%,其次為StenotrophomonaslactitubiM15T(99.66%)和StenotrophomonaschelatiphagaDSM 21508T(99.05%)。Neighbor-joining法構建菌株MST-5的系統進化樹,表明菌株MST-5屬于Stenotrophomonas屬,與StenotrophomonasindicatrixWS40T形成一個分枝(圖3)。因此將菌株MST-5初步鑒定為嗜麥芽窄食單胞菌屬。

(a)菌落形態;(b)透射電鏡(放大倍數為10 000倍)照片。(a) Colony morphology; (b) Transmission electron micrograph (10 000×).圖2 菌株MST-5的形態特征Fig.2 Morphological characteristics of strain MST-5
2.3 hppd基因的克隆
Blast分析表明,同一個屬的HPPD基因非常保守,其核酸序列和氨基酸序列相似性高達90%以上。雖然抗性菌株MST-5的基因組尚未測定,但和其親緣關系最近的菌株StenotrophomonasindicatrixWS40T的基因組序列已經公布(登錄號:PEJS00000000.1)。因此,在StenotrophomonasindicatrixWS40T氨基酸序列文庫中,通過同源序列比對找到其HPPD(WP_032951734.1)以及對應的hppd基因(Locus tag: CR918_RS17620),并以此設計引物對Sthppd-F和Sthppd-R(表1),以菌株MST-5基因組DNA為模板,利用PCR成功擴增出了hppd基因,命名為Sthppd。經測序,Sthppd長為1 071 bp,GC含量為62%,編碼356個氨基酸。
同源序列分析表明,StHPPD和已報道的抗性HPPD序列一致性較低(圖4)。如:StHPPD與來自Burkholderiasp. BW-1的BkHPPD相似性為55%,與來自PseudomonasfluorescensP.J.874的PfHPPD相似性為51%,與來自Ensifersp. Ace-21 的EnHPPD相似性為51%,與來自Pseudomonassp. AM-H4的PsHPPD的相似性為47%,與來自Sphingobiumsp. TPM-19的SpHPPD的相似性為49%。
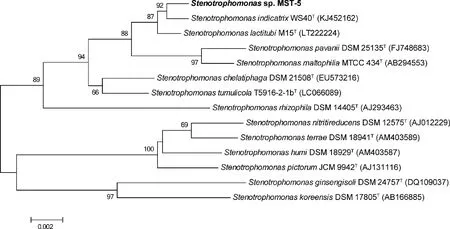
括號中的序號代表GenBank登錄號;節點上的數字表示Bootstrap值;刻度0.002表示序列偏差值。Accession numbers of sequences are given in parentheses. Bar, 0.01 substitutions per nucleotide position. Numbers at the nodes indicate the bootstrap values. Bar, 0.002 substitutions per nucleotide position.圖3 菌株MST-5基于16S rRNA基因序列的系統發育樹Fig.3 Phylogenetic tree reconstructed by the neighbour-joining method based on 16S rRNA gene sequences of strain MST-5
2.4 StHPPD的表達與純化
為測定StHPPD的酶活,將Sthppd克隆至pET29a(+)表達載體上,并轉化至E.coliBL21(DE3)進行異源表達,表達產物通過Co2+親和層析純化。SDS-PAGE分析表明純化的蛋白的分子量大約為42 000 u,與理論值40 100 u相符(圖5)。純化的StHPPD蛋白質量濃度為436 μg/mL。
2.5 StHPPD的酶活及對不同HPPD抑制劑的抗性分析
利用HPLC檢測StHPPD的酶活,如圖6所示,不添加酶的對照只有4-HPP特征峰,出峰時間為9.7 min;而添加StHPPD的處理在3.9 min有一個新的特征峰生成,與HGA標準品的出峰時間一致。HPLC檢測結果表明StHPPD具有4-羥基苯丙酮酸雙加氧酶的活性,可以在體外將4-HPP轉化成HGA。
為進一步研究StHPPD對3種HPPD抑制劑類除草劑的抗性,利用HPLC檢測了StHPPD在添加有不同濃度的HPPD抑制劑時殘留的酶活,結果顯示,硝磺草酮、苯吡唑草酮和二酮腈對StHPPD的IC50分別為15.5、10.8和28.2 μmol/L。
2.6 溫度、pH、金屬離子及化學添加劑對StHPPD酶活影響
如圖7(a)所示,StHPPD的最適反應溫度為30 ℃;在20~40 ℃范圍內,StHPPD的酶活能夠保持在50%以上;當溫度高于40 ℃時,StHPPD酶活快速下降;在50 ℃時,StHPPD基本喪失酶活。如圖7(b)所示,StHPPD的最適pH為7.0;pH為6.0~7.5,StHPPD酶活能保持在90%以上;當pH為8.0時StHPPD酶活能保持在50%左右;當pH<5.0或pH=9.0時,StHPPD基本檢測不到酶活。金屬離子和一些化學添加劑對StHPPD酶活的影響如表3所示。大部分金屬離子,如Al3+、Co2+、Hg2+、Cu2+、Ni2+、Zn2+和Ba2+,嚴重抑制StHPPD酶活;只有Fe3+和Mg2+不影響StHPPD的酶活;加入5 mmol/L脲對StHPPD酶活基本沒影響,但加入EDTA、SDS和DEPC之后,StHPPD酶活完全被抑制。

表3 金屬離子和化學添加劑對StHPPD酶活的影響Table 3 Effects of metal ions and chemicals on StHPPD activity
3 討論與結論
HPPD抑制劑類除草劑被認為是理想的抗除草劑轉基因靶標除草劑,近年來,HPPD抑制劑類除草劑抗性菌株的篩選及抗性基因的克隆成為當前的研究熱點。目前已有近10種HPPD抑制劑類除草劑抗性菌株被分離出來[6-11]。本研究分離到一株硝磺草酮抗性菌株MST-5,初步鑒定為嗜麥芽窄食單胞菌屬(Stenotrophomonassp.)。該屬細菌的HPPD抑制劑抗性是首次報道。MST-5能夠耐受5 000 μmol/L的硝磺草酮,與能耐受250 μmol/L硝磺草酮的Sphingobiumsp. TPM-19相比[12],Stenotrophomonassp. MST-5對硝磺草酮具有更高的抗性。
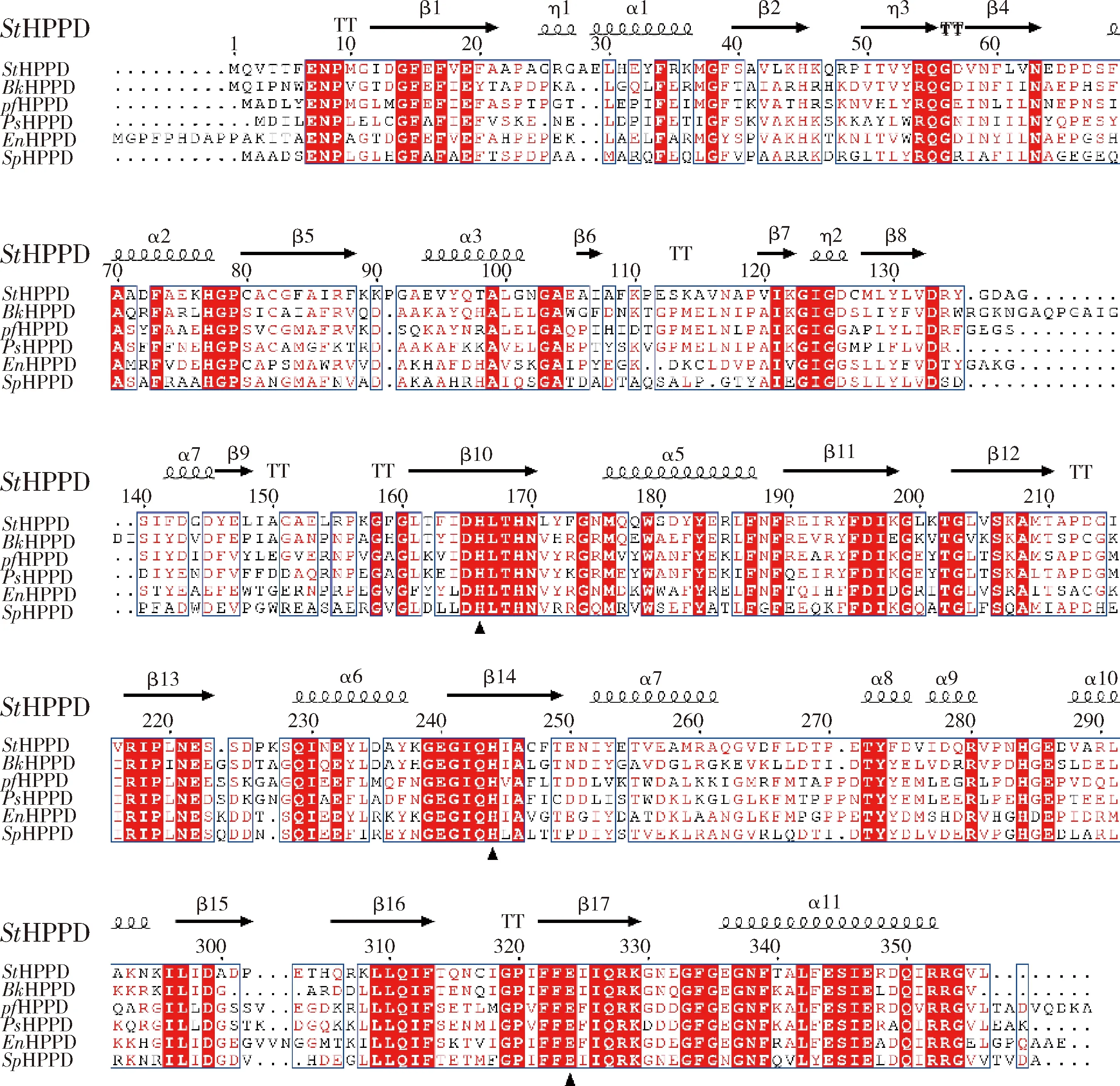
序列上方帶箭頭部分為StHPPD二級結構,包括α螺旋和β折疊,相同的序列背景用紅色背景標出,保守序列位點用方框標出。黑色填充三角標記的嚴格保守的殘基(His170,His248 和Glu332)是Fe(II)結合位點。Numbers above the amino acid sequences indicate the residue positions of StHPPD. The predicted StHPPD secondary structure is shown above the alignment with α-helices, β-strands, and turns. The conserved amino acid residues are emphasized in box and identical residues are shown with a red background. Strictly conserved residues (His170, His248 and Glu332) labeled with black filled triangles are Fe(II) binding site.圖4 HPPD氨基酸序列比對Fig.4 Multiple alignment of HPPD enzymes protein sequence
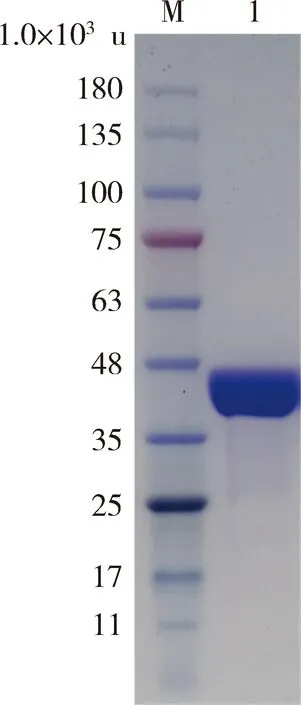
M:蛋白Marker;1:StHPPD純酶。Lane M: Standard protein molecular marker;Lane 1: Purified StHPPD.圖5 StHPPD的SDS-PAG檢測圖Fig.5 SDS-PAGE analysis of StHPPD
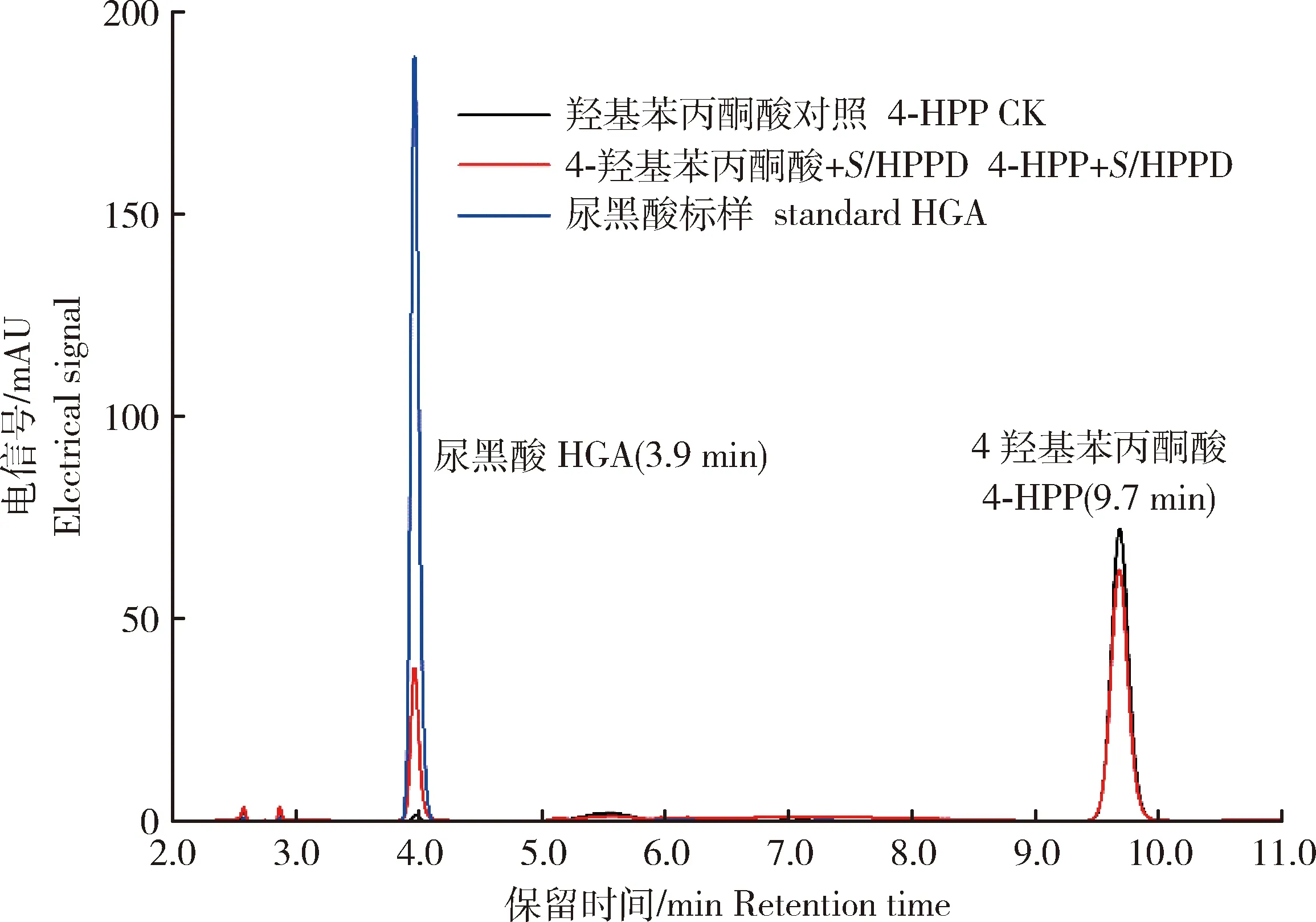
圖6 StHPPD催化4-HPP生成HGA的液相色譜圖Fig.6 HPLC analysis of 4-HPP transformation by StHPPD
國內外克隆獲得了多個HPPD抑制劑除草劑抗性HPPD基因[6~11],其中有2個HPPD抗性基因突變體,分別是從Pseudomonasfluorescens的HPPD中篩選得到的突變體GW336[8]和從Avenasativa的HPPD中篩選得到的突變體339[9],已經用于轉基因大豆生產中,并使轉基因大豆產生了對異噁唑草酮和硝磺草酮的較強抗性。本研究從Stenotrophomonassp. MST-5中克隆HPPD基因Sthppd,并外源表達StHPPD。最近,Liu等[12]和彭乾等[13]分離到2個抗性HPPD,分別命名為SpHPPD和EnHPPD。SpHPPD和EnHPPD表現出廣譜的HPPD抑制劑抗性,硝磺草酮、苯吡唑草酮和二酮腈對SpHPPD的IC50為4.2~8.7 μmol/L,對EnHPPD的IC50為12.7~35.1 μmol/L。在本研究中3種HPPD抑制劑對StHPPD的IC50為10.8~28.2 μmol/L,與SpHPPD和EnHPPD處在同一個數量級水平上。這說明StHPPD對多種HPPD抑制劑類除草劑具有較高的抗性,是優良的廣譜HPPD抑制劑類除草劑抗性基因資源,具有潛在應用價值。

數值以“平均值±標準偏差”表示;n=3。Data are shown as mean±SD. n=3.圖7 溫度(a)和pH(b)對StHPPD酶活的影響Fig.7 Effects of temperature (a) and pH (b) on the activity of StHPPD
同源序列分析結果顯示,StHPPD和已報道的抗性HPPD序列相似性僅為47%~51%,其C端序列相對保守,而N端序列則與已報道HPPDs差異較大。前人研究表明,HPPD一級結構中的C末端氨基酸序列高度保守,是酶的活性中心,而N末端同源性較低,沒有催化功能,在維持蛋白質穩定性方面發揮作用[12,20]。Liu等[12]通過對已報道的4-羥基苯丙酮酸雙加氧酶進行序列比對發現,雖然基因序列相似性不高,但各HPPD蛋白中與Fe2+結合的2個His和1個Glu高度保守。我們預測的StHPPD一級結構中也有三個催化活性中心Fe2+結合位點His166、His244和Glu332。這些結果說明StHPPD可能具有與已報道的HPPD蛋白類似的催化機制。
本研究分離到一株硝磺草酮高耐受菌株Stenotrophomonassp. MST-5,并從中克隆到硝磺草酮抗性基因Sthppd。抗性分析表明StHPPD對HPPD抑制劑類除草劑表現出較高的抗性,但其抗性機制和在轉基因植物中的抗性性能有待進一步研究。本研究為抗HPPD抑制劑類除草劑轉基因作物的培育提供優良的基因資源。
猜你喜歡
世界農藥(2019年3期)2019-09-10 07:04:10
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
長江蔬菜(2016年10期)2016-12-01 03:05:27
獸醫導刊(2016年12期)2016-05-17 03:51:29
現代農業(2016年5期)2016-02-28 18:42:36
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:14
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年22期)2015-02-28 22:05:11
