新疆地區牛分枝桿菌29個毒力基因的突變分析
2021-11-04 07:53:36董玉慧羅力嘉李夢瑩歐喜超劉春法范偉興趙雁林周向梅
中國農業大學學報 2021年9期
關鍵詞:基因突變
董玉慧 羅力嘉 李夢瑩 歐喜超 劉春法 范偉興 趙雁林* 周向梅*
(1.中國農業大學 動物醫學院,北京 100193;2.寧夏大學 農學院,銀川 750021;3.中國疾病預防控制中心,北京 102206;4.中國動物衛生與流行病學中心,山東 青島 266032)
結核病是由毒力和傳播能力不同的結核分枝桿菌復合群(Mycobacteriumtuberculosiscomplex,MTBC)成員感染人類和動物引起的一種人畜共患病[1]。MTBC成員主要包括引起人結核病的結核分枝桿菌、非洲分枝桿菌,以及從其他哺乳動物分離的牛分枝桿菌、田鼠分枝桿菌等[2-3]。牛結核病是一種主要由牛分枝桿菌引起的慢性消耗性疾病。牛分枝桿菌宿主范圍廣泛,可感染牛等多種動物及人類,給畜牧業生產帶來重大的經濟損失,并威脅人類健康,該病被世界動物衛生組織(OIE)列為B類動物疾病[4-5]。但是由于許多國家缺乏對牛結核病的長期監測和流行病學調查,牛分枝桿菌的流行率和對人類的影響被低估[6-7]。因此,評估牛分枝桿菌的遺傳多樣性及其與致病性和疾病嚴重程度的相關性,對養牛業的健康發展和公共衛生安全至關重要[7]。
MTBC可通過突變、重組和自然選擇等途徑實現全基因組多樣化,從而在全球廣泛流行和傳播[8-9]。結核分枝桿菌系統發育譜系具有顯著的地理分布特點,呈現出與人類宿主密切的共同進化趨勢[9]。不同譜系的分離株因多種表型如耐藥性、毒力和致病性而有所不同,這些表型影響疾病的嚴重程度,而多樣性是由影響表達產物結構和功能的基因突變造成的[10]。利用轉座子突變體庫結合不同的體內篩選方法,鑒定出分枝桿菌大量的毒力基因,這些毒力基因大多數編碼脂質代謝相關酶、細胞表面蛋白、調節因子和信號轉導系統蛋白,影響分枝桿菌在宿主巨噬細胞內的存活[11]。目前,中國新疆地區流行牛分枝桿菌毒力變化和基因突變特征仍不明確。比對不同毒力菌株的毒力基因序列,找到關鍵突變位點,是發現細菌致病機理和進化規律的重要方法。Mycobacteriumbovis(M.bovis) C68004分離自上世紀60年代北京結核病牛,M.bovisN由本實驗室近期從全身粟粒性結核病牛分離而來。在BALB/c小鼠模型上研究發現,與M.bovisC68004菌株相比,M.bovisN菌株表現出更強的致病性和更高的致死率。全基因組測序結果表明,M.bovisC68004菌株和M.bovisN菌株共存在750個SNPs (Single nucleotide polymorphisms)和145個INDELs(Insertion/Deletion)差異。與毒力因子數據庫匹配后,發現29個毒力基因發生突變[12]。
本研究通過對分離自新疆地區的66株牛分枝桿菌的29個毒力基因進行測序,將其序列與標準株M.bovisAF2122/97及本實驗室保存的M.bovisC68004和M.bovisN進行序列比對,初步評估新疆地區牛分枝桿菌的毒力情況,并進而探討基因突變對牛分枝桿菌毒力的影響,為后續深入研究牛分枝桿菌的致病機理和進化規律奠定了基礎。
1 材料與方法
1.1 試驗材料
中性改良羅氏基礎培養基(Celnovte,中國);細菌基因組DNA提取試劑盒(Tiangen,中國);Zihel & Neelsen染色試劑盒、PCR Mix、DL 2000 DNA Marker、6 x DNA loading Buffer(Solarbio,中國);PCR擴增儀和凝膠成像分析系統(Bio-Rad,美國);本實驗中所用的試驗菌株包括:從新疆地區分離的66株牛分枝桿菌(由中國疾病預防控制中心結核病預防控制中心保存),M.bovisC68004(購自中國獸醫藥品監察所),M.bovisN(由中國農業大學動物醫學院國家動物傳染性海綿狀腦病實驗室保存)。
1.2 試驗方法
1.2.1牛分枝桿菌的培養
取200 μL凍存菌液,將其均勻涂于改良羅氏基礎培養基上,37 ℃溫箱培養2~3周。
1.2.2細菌基因組DNA的提取
在生物安全三級實驗室(P3)中操作:刮取約20~100 mg(濕重)細菌新鮮培養物于1.5 mL螺旋口離心管中,水浴80 ℃滅活30 min,根據P3的操作規范要求,可將滅活的菌液移出P3,其他步驟參照細菌基因組DNA提取試劑盒說明書進行。
1.2.3引物的設計與合成
參考GenBank上已發表的29個牛分枝桿菌毒力基因序列,使用Primer 5.0軟件設計引物序列,送至金唯智生物科技有限公司合成,引物的序列見表1。
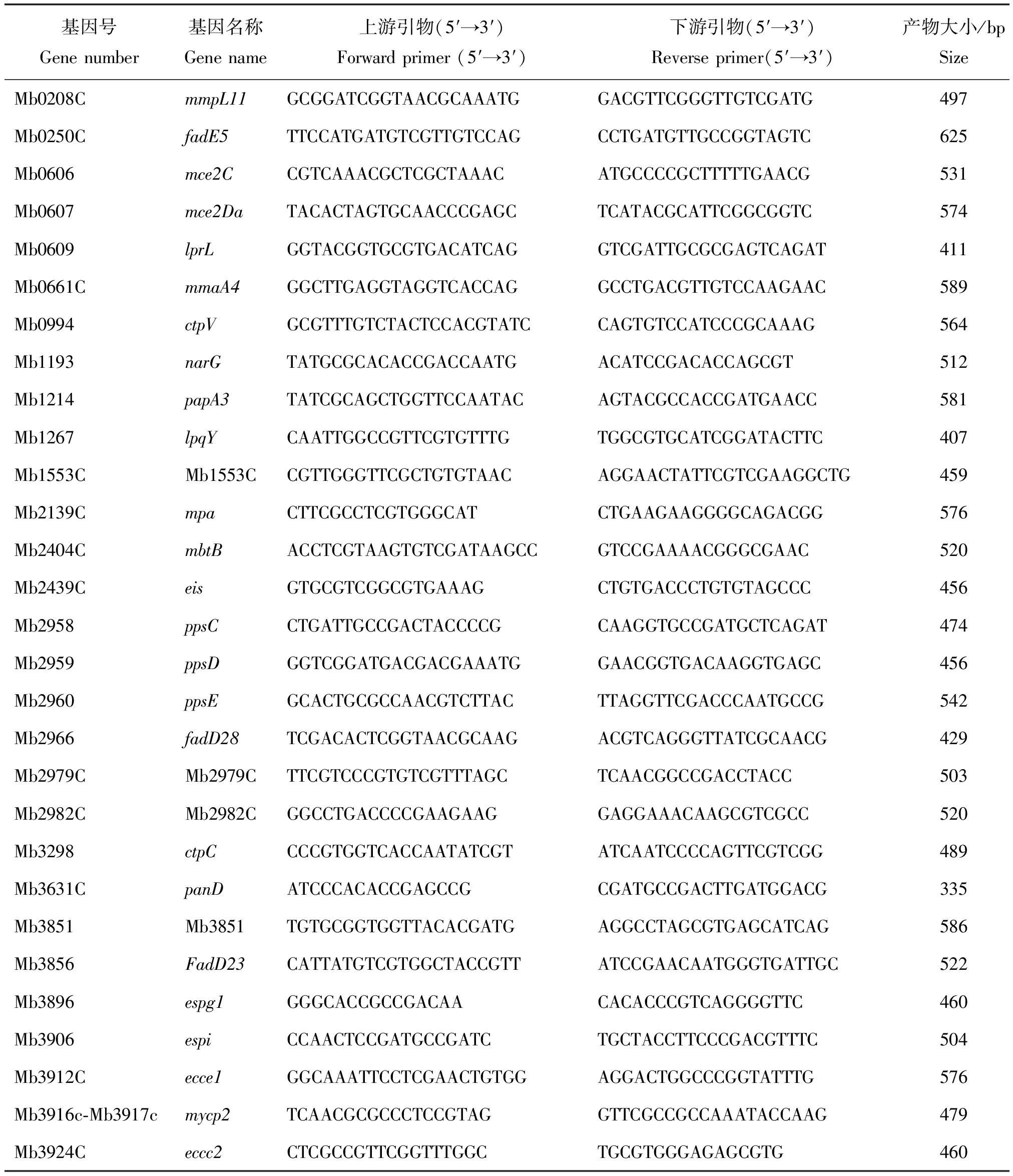
表1 新疆地區牛分枝桿菌29個毒力基因的引物序列和目的擴增產物大小Table 1 Primer sequences and targeted amplification product size of 29 virulence genes from M. bovis in Xinjiang
1.2.4PCR反應體系和程序
反應體系為25 μL,其中模板1 μL,PCR Mix 12.5 μL,ddH2O 10.5 μL,上游引物0.5 μL(10 μmol/L),下游引物0.5 μL(10 μmol/L)。反應程序為:95 ℃預變性20 min,95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸60 s(35個循環),72 ℃延伸10 min,4 ℃保存。
1.2.5PCR產物回收和測序
將各基因的PCR產物送至金唯智生物科技有限公司進行Sanger測序,并在NCBI/GenBank上進行序列比對。
1.3 序列分析
利用PROVEAN和SOPMA在線分析軟件預測基因突變對蛋白質功能的影響,評價牛分枝桿菌毒力基因突變對蛋白質功能或二級結構的影響。
PROVEAN軟件通過執行BLAST搜索以識別同源序列,并生成PROVEAN得分,可預測單個或多個氨基酸替換、插入和刪除對蛋白功能的影響。PROVEAN對單個氨基酸變異的功能預測效果與其他同類工具(如SIFT和PolyPhen-2)具有良好的一致性[13]。SOPMA是一種新的預測蛋白二級結構的方法,通過5種相互獨立的方法進行預測,并將結果匯集整理成一個“一致預測結果”,具有較高預測蛋白二級結構的成功率[14]。
2 結果與分析
2.1 新疆地區牛分枝桿菌29個毒力基因的擴增
使用設計的引物,分別擴增66株牛分枝桿菌的29個毒力基因片段。PCR擴增產物經1.5%瓊脂糖凝膠電泳分析,產物條帶大小符合預期設計目的片段,隨后進行Sanger測序(圖1)。
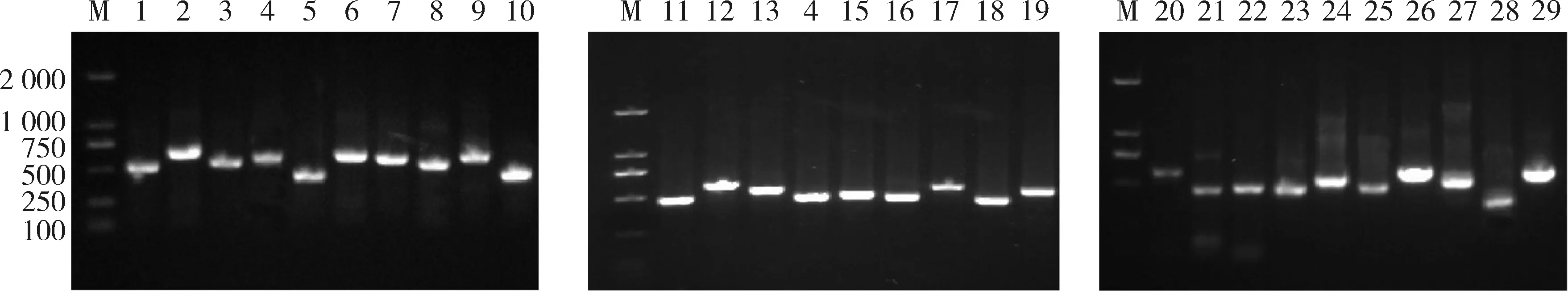
M: DNA分子量標準Trans2K; 1~29: Mb0208C、Mb0250C、Mb0606、Mb0607、Mb0609、Mb0661C、Mb0994、Mb1193、Mb1214、Mb1267、Mb1553C、Mb2139C、Mb2404C、Mb2439C、Mb2958、Mb2959、Mb2960、Mb2966、Mb2979C、Mb3851、Mb3924C、Mb3916C-Mb3917C、Mb3896、Mb2982C、Mb3906、Mb3912C、Mb3298、Mb3631C、Mb3856.M: Trans2K DNA Marker; 1~29: Mb0208C、Mb0250C、Mb0606、Mb0607、Mb0609、Mb0661C、Mb0994、Mb1193、Mb1214、Mb1267、Mb1553C、Mb2139C、Mb2404C、Mb2439C、Mb2958、Mb2959、Mb2960、Mb2966、Mb2979C、Mb3851、Mb3924C、Mb3916C-Mb3917C、Mb3896、Mb2982C、Mb3906、Mb3912C、Mb3298、Mb3631C、Mb3856.圖1 新疆地區牛分枝桿菌29個毒力基因擴增Fig.1 PCR amplification of 29 virulence genes of M. bovis in Xinjiang
2.2 新疆地區牛分枝桿菌臨床分離株基因變異的檢測
為研究牛分枝桿菌臨床分離株毒力基因的變異情況,本研究將66株臨床分離株與標準株M.bovisAF2122/97以及實驗室保存的M.bovisC68004和M.bovisN的基因序列進行比對。結果發現,與標準株M.bovisAF2122/97基因序列相比,其中有17個毒力基因在超過50%的臨床分離株存在突變(圖2)。此外,在29個檢測的毒力基因中,50%以上臨床分離株與M.bovisN突變位點序列一致的共有25個基因,50%以上臨床分離株與M.bovisC68004序列僅在3個基因中表現一致。這些結果表明中國新疆臨床分離的牛分枝桿菌毒力基因突變特征與致病性更強的M.bovisN具有很高的相似性。
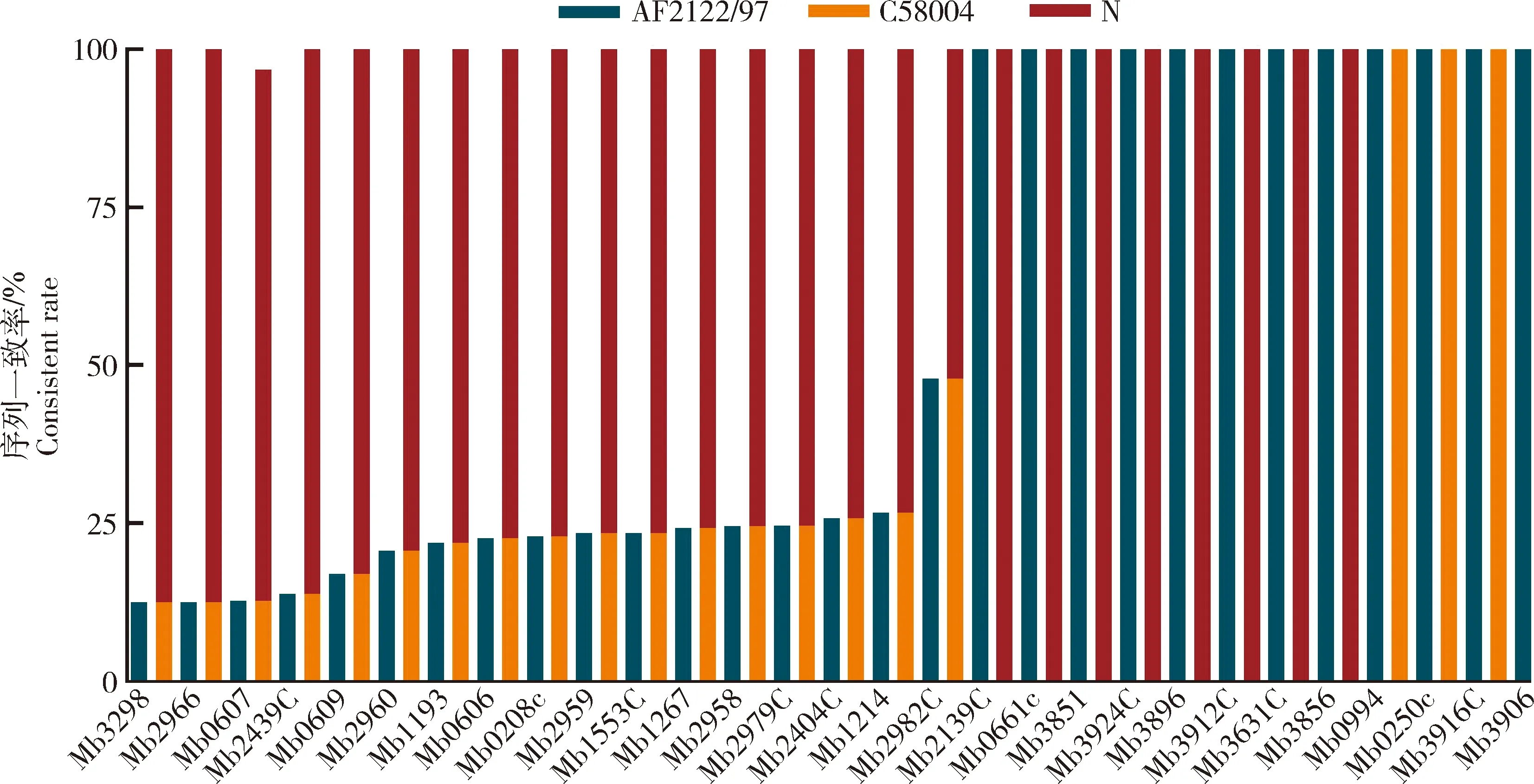
序列一致率:毒力基因突變位點與參比菌株序列一致的臨床分離菌株數/臨床總分離菌株數×100%。Consistent rate: Number of clinical isolates with the same virulence gene mutation site as the reference strain/umber of total cunleal isolates.圖2 牛分枝桿菌臨床分離株毒力基因與AF2122/97、C68004、N菌序列比較Fig.2 Comparison of virulence gene sequences in clinical isolates of M. bovis with AF2122/97,C68004 and N
2.3 基因變異對蛋白功能的影響
2.3.1SNP對蛋白功能的影響
蛋白質編碼區的SNPs可影響氨基酸序列,從而改變蛋白質的結構和功能。在牛分枝桿菌臨床分離株中的12個基因發現了廣泛的堿基替換。為分析SNP對蛋白質功能的影響,本研究使用PROVEAN軟件[13]預測非同義突變對蛋白生物學功能的影響,當PROVEAN評分等于或低于-2.5時,則該點突變對蛋白功能具有“有害”影響;當PROVEAN評分高于-2.5時,則為“中性”突變。結果揭示Mb2958、Mb2982C和Mb1553C基因的非同義突變PROVEAN評分均低于-2.5,表明可能對蛋白質生物學功能有顯著影響。另外,其余基因的PROVEAN評分均高于-2.5,說明基因發生的這些突變對蛋白功能的影響不明顯(圖3)。
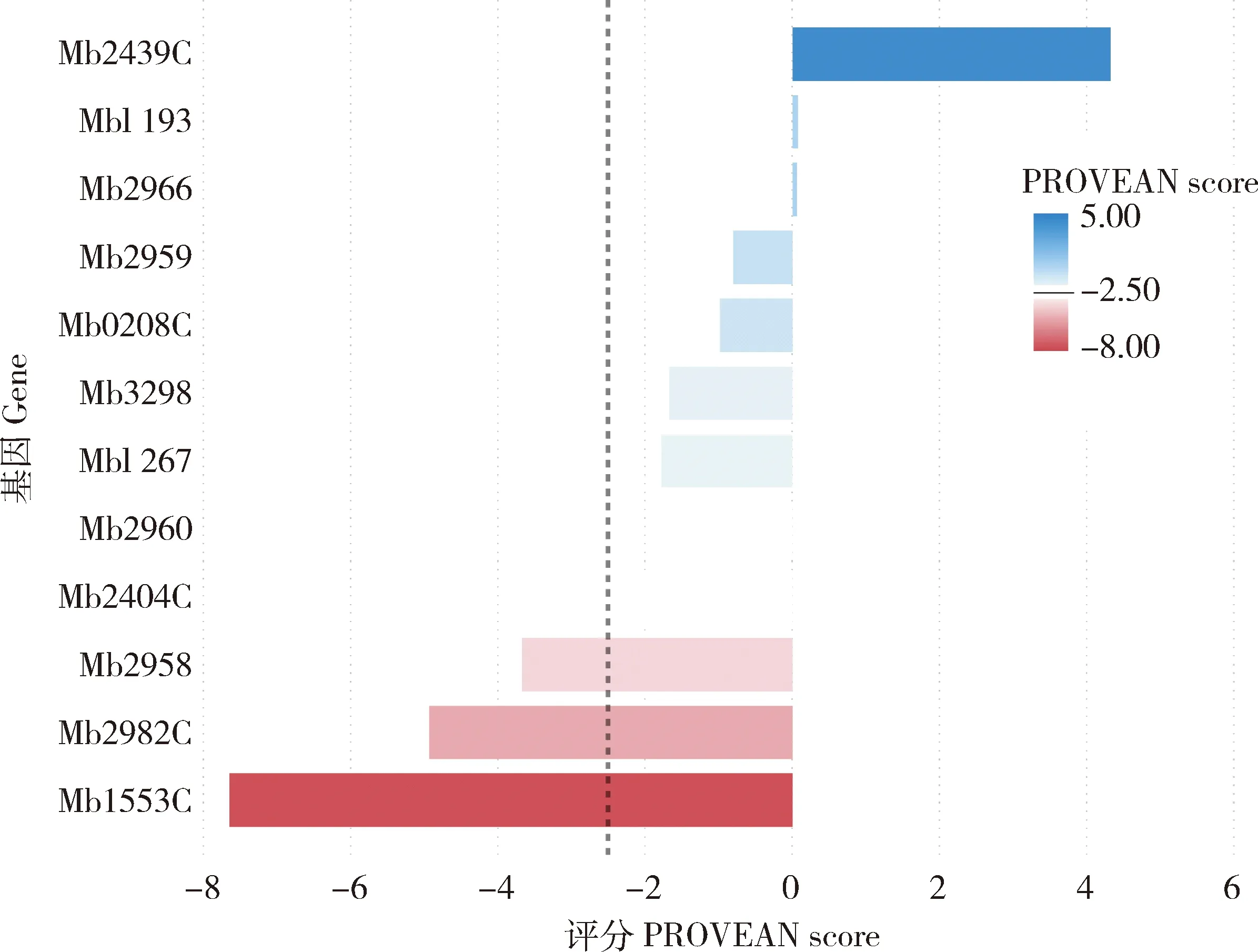
圖3 預測牛分枝桿菌非同義SNP對蛋白質生物學功能的影響Fig.3 Prediction of the effect of nonsynonymous SNP in M.bovis isolates on the biological function of the proteins
2.3.2蛋白及其突變體二級結構的預測
使用SOPMA(Self-optimized prediction method with alignment)服務器[14]預測蛋白質及其突變體的二級結構。結果發現5種廣泛存在于臨床菌株的基因變異導致蛋白質的二級結構發生變化。Mb0607基因的第1 395位核苷酸插入一個堿基(C,胞嘧啶)導致肽鏈的延長(圖4(a))。Mb0609基因編碼的蛋白質終止密碼子突變為Gln,使得翻譯終止延后(圖4(b))。Mb0606基因第195位氨基酸Gln突變為終止密碼子,導致轉錄提前終止(圖4(c))。Mb2979C基因的第232位核苷酸插入了一個堿基(C,胞嘧啶),這種移碼突變造成了肽鏈的縮短(圖4(d))。此外,Mb1214基因的第367位核苷酸插入一個鳥嘌呤,并引起下游的密碼子發生改變,使肽鏈縮短(圖4(e))。結果表明,這5種常見的基因變異可導致相應蛋白二級結構的改變。

W:野生型蛋白二級結構;M:突變體蛋白二級結構W: Secondary structure of wild-type protein; M: Secondary structure of mutant protein圖4 牛分枝桿菌毒力因子基因突變對其蛋白二級結構影響的預測Fig.4 Prediction of effects of virulence gene mutants on the secondary structure of proteins
3 討 論
由于全球多數地區缺乏對牛分枝桿菌基因組多樣性的調查和可比的基因分型數據,目前,對牛分枝桿菌全球分布和進化的研究較少[4]。最近的一項研究表明牛分枝桿菌至少存在4個全球譜系,具有明顯的地理分布特點[15]。但來自亞洲和非洲的牛分枝桿菌基因組數據匱乏,牛分枝桿菌全球分布譜系仍不完整[15-16]。本研究對我國新疆分離的66株牛分枝桿菌的毒力基因突變進行了初步研究,并將其毒力基因序列與兩株牛分枝桿菌臨床分離株(M.bovisC68004和M.bovisN)和牛分枝桿菌標準株M.bovisAF2122/97進行了序列比對。本團隊之前研究發現,與M.bovisC68004株相比,M.bovisN株對小鼠的致病性很強,M.bovisN更強的致病性可能與其毒力基因的突變有關[12]。本研究在檢測的29個毒力基因中,發現50%以上的臨床分離株與M.bovisN突變位點序列一致的基因有25個。這在一定程度上反映了目前新疆地區流行牛分枝桿菌毒力的情況。
近年來,對結核分枝桿菌的致病性、毒力和持久性的分子基礎研究取得了相當大的進展,牛分枝桿菌與結核分枝桿菌的基因組序列相似性超過99.95%[11,17],因此,牛分枝桿菌毒力基因的研究可參考結核分枝桿菌。本研究分析發現臨床菌株廣泛存在的Mb2958、Mb2982C和Mb1553C基因非同義突變對蛋白功能影響較大,對牛分枝桿菌毒力可能有一定影響。Mb2958(ppsC)與結核分枝桿菌Rv2933具有100%相似性,編碼I型聚酮合成酶,催化脂類毒力因子Phthiocerol dimycocerosates(PDIM)和酚糖脂(Phenolic glycolipids,PGL)的生物合成關鍵步驟[18-19]。Mb2982C與結核分枝桿菌Rv2958C具有83.3%的相似性,編碼參與PGL-tb合成的糖基轉移酶。PGL包含有一個由長鏈β二醇結構形成的脂質核心,與多甲基分枝的脂肪酸形成二酯,存在于細胞壁,僅少量結核分枝桿菌可以合成PGL-tb,PGL-tb碳水化合物組分主要形式為2,3,4-三-O-甲基-L-吡喃巖藻糖基-α-(1-3)-L-吡喃鼠李糖基-α-(1-3)-2-O-甲基-L-吡喃鼠李糖[20-21]。而牛分枝桿菌由于基因移碼突變合成一種截短形式的PGL-tb,稱為分枝桿菌糖脂B,碳水化合物區域限制為2-O-甲基鼠李糖[22]。PGL是致病性分枝桿菌的一個重要毒力因子,在抗氧化應激、免疫調節和細胞趨化過程中發揮作用,該化合物有多種結構變異。這些結構改變的影響雖尚未被評估,但可能解釋分枝桿菌引起疾病的不同特征[20]。糖肽脂(Glycopeptidolipids,GPLs)是存在于許多分枝桿菌表面的特異性糖脂類,其特征是糖基化模式的高度變異性,具有多種生物活性,Gtf3是一種催化合成三糖基化GPLs的糖基轉移酶[23]。Mb1553C與結核分枝桿菌Rv1526C基因序列具有100%一致性,為Gtf3的同源物。Mb2958、Mb2982C和Mb1553C基因突變均可能導致其編碼的酶活性發生改變,從而對細胞壁脂類合成產生影響,導致牛分枝桿菌致病性增強。
本研究發現超過半數臨床菌株存在的Mb0607(Mce2Da)、Mb0609(lprL,Mce2E)、Mb0606(Mce2C)、Mb2979C和Mb1214(Papa3)基因突變導致了蛋白二級結構的顯著變化。哺乳動物細胞入侵(Mammalian cell entry,Mce)基因是Mce操縱子的組成部分,其編碼的蛋白在分支桿菌進入哺乳動物細胞和在巨噬細胞存活過程中發揮有重要作用,哺乳動物細胞入侵蛋白在分枝桿菌細胞表面的位置與Mce操縱子在哺乳動物細胞侵襲中的潛在作用一致,因此被視為重要的毒力因子[24-25]。本研究表明基因突變導致Mce2Da、Mce2E和Mce2C蛋白二級結構發生顯著變化,其中,Mce2E發生于終止密碼子區域的基因突變,使其表達產物和M.tuberculosisH37Rv具有100%一致性,因此進一步對這些Mce操縱子進行深入研究將有助于闡明牛分枝桿菌毒力變異的機制。Mb2979C與結核分枝桿菌Rv2955C相似性為99.7%,Rv2955C參與結核分枝桿菌細胞壁PGL-tb的生物合成[26]。Mb1214與結核分枝桿菌Rv1182序列100%一致,編碼Polyacyltrehalose(PAT)生物合成所必需的酰基轉移酶[27]。后2種基因突變導致的編碼蛋白結構的變化可能會對其生物效應產生影響,從而提高牛分枝桿菌的毒力。
基因研究為分枝桿菌屬的遺傳多樣性提供了一個精細化的理解,這些結果為MTBC的宏觀和微觀進化提供了一個新的視角[28-29]。本研究在Mb2958、Mb2982C、Mb1553C、Mb0607、Mb0609、Mb0606、Mb2979C和Mb1214這8個毒力基因中發現了新疆牛分枝桿菌臨床分離株廣泛存在的突變,這些基因突變可能與牛分枝桿菌在牛群中更強的適應性和致病性有關,其具體機制尚需進一步研究。
綜上,本研究通過對牛分枝桿菌毒力基因進行檢測、測序和比對,并分析基因突變對其編碼蛋白功能和結構的可能影響,初步反映了我國新疆地區牛分枝桿菌流行菌株的毒力情況,為后續研究牛分枝桿菌分子流行病學和闡明牛分枝桿菌致病機理及進化規律奠定了基礎。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫學影像學雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學生導刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國現代醫學雜志(2015年26期)2015-12-23 11:04:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
