運動介導miRNAs調控白色脂肪棕色化的研究進展
2021-11-13 03:22:08劉源
體育科技文獻通報 2021年11期
劉 源
世界衛生組織(WHO)公布的數據顯示,2016年全球18歲及以上的成年人中逾19億人超重,其中超過6.5億人肥胖,肥胖成年人占全球人口的13%。肥胖癥是人們過多地攝入能量,使能量攝入遠大于能量消耗,并且,肥胖癥與很多慢性疾病息息相關,如:II型糖尿病、高血脂、脂肪肝、心血管疾病等。肥胖問題及其產生的一系列慢性并發癥給人們帶來了沉重的經濟負擔,造成生活上的困惑。令人遺憾的是,目前的抗肥胖策略旨在限制能量攝取,且各國沒有形成有效應對肥胖的干預機制[1]。近期,許多基礎及臨床研究已經證實白色脂肪在一定條件下可以向棕色化轉變,且體內棕色脂肪的含量和分布與肥胖及其引起的一些慢性疾病有密切關系,體內棕色脂肪越多,肥胖及代謝異常疾病發病的可能性越少,所以,深入研究白色脂肪棕色化的作用機制對促進目標減肥、減少代謝異常疾病的發病率具有重要意義[2-5]。本文從miRNAs對棕色脂肪組織的生成、白色脂肪棕色化的調控作用,以及運動對miRNAs、白色脂肪棕色化的調節作用及其潛在機制做一綜述,分析探討白色脂肪棕色化的研究趨勢和應用前景,從而為肥胖癥及代謝異常疾病的治療提供新思路。
1 miRNA
miRNA是在真核生物中發現的一類具有調控功能的內源性非蛋白編碼單鏈小分子RNA,它是由20~25個核苷酸組成,通過完全或不完全堿基互補配對原則與特定靶基因的mRNA的3’或5’UTR區結合,能夠抑制mRNA翻譯或直接降解其特定mRNA的作用,對調節基因表達過程具有重要作用[6-9]。研究發現,一個miRNA可以同時調控幾個基因,一個基因也可以同時受幾個miRNAs調控。而且其不僅在細胞的生長、發育、增殖和分化過程中起多種作用,對于機體細胞的棕色化也有非常關鍵的作用。目前,miRNA對白色脂肪棕色化影響的研究已經成為治療肥胖問題的新靶點,進一步探索miRNAs在脂肪細胞和脂肪組織中的分子作用,這將揭示促進脂肪細胞能量消耗的新方法,最終可能成為新型抗肥胖治療劑。
2 棕色脂肪和白色脂肪棕色化
2.1 棕色脂肪
眾所周知,人體中有白色脂肪(WAT)和棕色脂肪(BAT)[10]。白色脂肪是能源儲存的場所,主要作用是通過甘油三酯的形式儲存機體過剩的能量,且能分泌多種脂肪因子調節能量代謝[11-13]。而棕色脂肪呈褐色,其主要作用是加快能量的消耗和釋放,相比與白色脂肪,棕色脂肪細胞體積小,含有更多線粒體,并且解偶聯蛋白1(uncoupling protein 1,UCP-1)在這些線粒體中高度富集[14,15]。UCP-1將底物氧化與ATP產生解偶聯,促進游離脂肪酸代謝,從而產生熱量,促進機體的代謝和能量的消耗[16-20]。在嚙齒類動物中,棕色脂肪主要位于肩胛間區域,是一種類似于蝴蝶的蝶狀組織。在人類中,棕色脂肪主要位于頸部、鎖骨以上和縱隔區域[21]。棕色脂肪組織的活性與體重指數呈負相關,幫助人們認識體內棕色脂肪細胞在控制體重方面的生理相關性[22]。
2.2 白色脂肪棕色化
根據大量的研究顯示,人體中白色脂肪和棕色脂肪組織是混合分布的,而不是完全隔離的,即白色脂肪中存在著棕色樣的脂肪細胞,因為其中棕色樣的脂肪細胞很少,白色脂肪占主導地位,所以顯示出白色脂肪的生理作用。最新發現,哺乳動物中除了這兩種脂肪,還有一種新型的脂肪,被命名為米色脂肪,它是一種靜息細胞,分布于白色脂肪組織中,在冷暴露或腎上腺素受體受刺激的情況下,表現出類似于棕色脂肪細胞的特點,米色脂肪被激活后表現出棕色脂肪細胞特點的這一過程,被稱作“白色脂肪棕色化”[23-25]。促使人體中的白色脂肪向棕色化轉變有利于減少多余的白色脂肪在體內堆積,提高脂肪的利用價值,預防和治療人們的肥胖癥和代謝疾病,促進人體健康[26]。
3 miRNAs對白色脂肪棕色化的影響
miRNAs控制著大部分人類和鼠類基因的表達。多項研究表明,miRNAs對棕色脂肪細胞的分化生成有正反調節兩種方式。采用合理的方法,可以調整miRNAs對脂肪細胞的影響,促進白色脂肪棕色化,從而有效干預肥胖。隨著研究的推進,越來越多的miRNAs對脂肪的分化以及棕色化的調控作用被發現。下面我們將介紹miR-196a、miR-455、miR-30和miR-26對白色脂肪棕色化的正向調節作用以及miR-155、miR-150、miR-133和miR-129-5p對白色脂肪棕色化的負向調節作用。
3.1 正向調控棕色化的miRNAs
在成年人中發現的功能性棕色脂肪細胞開辟了這些細胞在治療肥胖及其相關疾病中的途徑[27]。有研究顯示,miR-196a可以介導棕色脂肪的形成[28]。研究發現miR-196a在白色脂肪祖細胞的棕色脂肪形成過程中抑制了白色脂肪基因Hoxc8在轉錄后的表達。低溫條件下,小鼠皮下白色脂肪棕色化過程中伴隨著miR-196a的表達明顯升高,miR-196a轉基因小鼠與正常對照組的小鼠相比,胰島素的敏感性提高,血糖明顯降低,能夠有效地防止高脂飲食引起的肥胖[28]。分析發現,在寒冷刺激下引起的小鼠白色脂肪棕色化過程中,miR-196a的表達明顯升高,抑制了Hoxc8的表達,Hoxc8作為一種白色脂肪基因,能夠通過抑制C/EBPβ的功能進而誘導棕色脂肪細胞的生成,所以,miR-196a-Hoxc8-C/EBPβ信號通路可能是誘導棕色脂肪形成以對抗肥胖和II型糖尿病治療的新靶點[28]。
miR-455在寒冷刺激或者骨形態發生蛋白7(Bone morphogenetic protein 7,BMP7)的誘導下可以使棕色脂肪細胞特異性表達,促進棕色脂肪的形成[29]。miR-455在棕色脂肪組織細胞以及白色脂肪前體細胞中過表達都可以促進細胞的分化,增加脂質體的累積,促進多種棕色脂肪特異性基因的表達。相反,miR-455的表達被抑制,則棕色脂肪的生成被抑制。有研究發現,通過高脂飲食誘導的miR-455轉基因小鼠的腹內位置出現白色脂肪棕色化,其抵御寒冷和產熱的能力增加,體重明顯降低。而miR-455敲除的小鼠體內棕色脂肪組織明顯減少,同時,miR-455的敲除抑制了過氧化物酶體增殖物激活受體γ(peroxisome proliferator-activated receptorγ,PPARγ)、UCP-1以 及PGC-1α(Peroxisome proliferator activated receptorγcoactivator-1α,)的表達。miRNA-455的3個作用靶點包括Runx1t1、Necdin和低氧誘導因子-1α亞基抑制劑(HIF-1αn)。miRNA-455通過HIF-1αn作用,增強AMPKα1的活性,使PGC-1α的表達增強,從而使線粒體合成加速,使脂肪分解的基因表達增強。因此,miR-455對白色脂肪棕色化的形成具有正向調節的作用。
miRNA-30家族的miR-30b和miR-30c能夠提高機體的產熱能力,對白色脂肪棕色化過程具有促進作用[30]。在寒冷刺激或者腎上腺受體激活狀態下,miR-30b/c通過上調產熱基因UCP-1和Cidea的表達,增強腹股溝區的白色脂肪基質血管成分產熱基因以及棕色脂肪細胞線粒體的呼吸作用,促進棕色脂肪組織的分化。相反地,有實驗結果證明了miR-30b/c表達被抑制會導致UCP-1的表達以及棕色脂肪細胞線粒體的呼吸作用下降。miR-30b/c對UCP-1和Cidea的作用是通過靶向作用受體交叉蛋白140(receptor-interacting protein140,RIP140)實現的,RIP140是產熱基因UCP-1和Cidea的輔助抑制物[31]。因此,miR-30b/c可以正向調控棕色脂肪,促進白色脂肪棕色化。
miRNA-26家族中的miRNA-26a和miRNA-26b是白色脂肪和米色脂肪分化的關鍵調節因子[32]。miRNA-26a在棕色脂肪組織中含量豐富,并且通過寒冷刺激后在白色脂肪細胞分化時表達增強。模擬miRNA-26a/b對白色脂肪和米色脂肪的作用,是通過UCP-1、PGC-1α和aP2這三種棕色脂肪特異性基因的表達,進而增加UCP-1陽性細胞實現的。miRNA-26a/b是通過金屬鈦酶域-17(ADAM17)作用于白色和米色脂肪的形成,它可能是抑制脂肪形成或者抑制棕色化的因子,但目前其作用機制并不明確。所以,明確ADAM17對脂肪合成和棕色化的作用,對ADAM17的上下游進行深入的探究很有必要。
上述可見,miR-196a、miR-455、miR-30和miR-26這四種正向調控棕色化的基因,分別通過Hoxc8、PGC-1α、RIP140以及ADAM17這四種基因的作用途徑來促進白色脂肪的棕色化,并且該過程是在寒冷刺激或是腎上腺受體激活狀態下發生的。這些miRNAs通過特定靶基因的作用通路來正向調控棕色,是預防和治療肥胖的新靶點。
3.2 負向調控棕色化的miRNAs
最初的研究揭示了白色脂肪和棕色脂肪細胞中miRNAs的差異表達和棕色脂肪細胞中肌肉特異性miRNAs的表達[33]。Chen等[34]人鑒定了miR-155是妨礙白色脂肪棕色化的負調節因子。最近的一項研究發現,miR-155參與棕色脂肪形成[35,36]。使用深度測序,研究確定了在棕色脂肪形成過程中差異表達的miRNAs,在這些miRNA中,miR-155具有特別重要的意義,因為它有效地抑制了棕色脂肪的生成。研究顯示miRNA-155富含棕色脂肪組織并在棕色脂肪細胞增殖時高度表達,但誘導分化后下降。更有趣的是,miR-155及其靶標成脂轉錄因子CCAAT/增強子結合蛋白β(C/EBPβ),形成一個雙穩態反饋回路,整合了調節增殖或分化的激素信號,產生雙向負反饋調節作用。相反,miR-155缺陷小鼠表現出棕色脂肪組織的增加和白色脂肪組織的“褐變”。分析得出,抑制miR-155的表達,可以增強棕色脂肪細胞分化,并在白色脂肪細胞中誘導棕色樣脂肪細胞表型褐變。這些數據表明,miRNA-155和CCAAT/增強子結合蛋白β的雙穩態環調節棕色脂肪細胞分化,從而控制棕色和米色脂肪細胞的發育[37]。
miR-150對白色脂肪棕色化具有負向調控的作用。靶向缺失KH-type剪切調控蛋白(KH-type splicing regulatory protein,KSRP)是一種在多個水平上調節基因表達的RNA結合蛋白,能夠降低身體肥胖的發生[38]。棕色脂肪選擇性基因的表達在KSRP缺失小鼠的皮下以及腹股溝的白色脂肪中增加,白色脂肪中miR-150的表達由于缺乏KSRP的初級加工受損而下降。miR-150直接靶向并抑制PRDM16,同時抑制miR-150的表達,由KSRP缺失引起的棕色脂肪基因的表達升高。該研究揭示了KSRP消融通過抑制miR-150表達,從而增強了白色脂肪組織中的棕色脂肪基因程序[38]。
miR-133在心臟和骨骼肌的生理學中具有確定的作用,其對白色脂肪棕色化具有負向調控的作用[39-41]。Trajkovski等[42]人已經描述了miR-133a和-133b參與棕色和米色脂肪形成,冷暴露導致小鼠棕色脂肪和皮下白色脂肪中這兩種miRNA顯著降低。在棕色前脂肪細胞分化過程中,抑制miR-133a/b的表達,使UCP-1及其上游調節劑PRDM16表達增強,導致細胞呼吸增加。相反,miR-133模擬物的轉染使棕色和一般脂肪細胞標志物的表達變差。用從皮下白色脂肪中分離的前脂肪細胞也觀察到miR-133的類似作用。進一步的體外實驗證實,β-腎上腺素能產生刺激,抑制肌細胞增強因子2(Mef2)轉錄因子的表達,由此降低miR-133a/b水平,導致miR-133靶標PRDM16的直接增加,產生白色脂肪組織的褐變。最近的一項研究重申了miR-133a對棕色和米色脂肪細胞分化的調節作用[43]。作者使用了敲除小鼠模型,其中通常產生成熟miR-133a的4個等位基因中的3個被刪除。雖然這種miR-133a的驟減不影響棕色脂肪中的棕色標記基因表達,但觀察到明顯的白色脂肪的“褐變”。此外,與具有匹配的年齡,性別和遺傳背景的野生型小鼠相比,其葡萄糖耐量和胰島素敏感性具有顯著改善。另外,Liu等[44]人的研究提示miR-133a/b的雙重作用,控制脂肪組織以及骨骼肌中的非顫抖產熱。
miR-129-5p對白色脂肪棕色化具有負調節的作用。它作為一個對機體非常重要的腫瘤抑制因子,可以有效地抑制癌細胞的增殖分化。中南大學[45]關于miRNA的微陣列分析研究表明,miR-129-5p在胰島素抵抗的3T3細胞中高度表達,可以推測出,miR-129-5p可能在脂肪細胞中發揮作用。有研究表明,miR-129-5p在小鼠皮下的白色脂肪和棕色脂肪中表達較高,其表達水平在脂肪細胞分化期間呈現升高趨勢。并且過表達miR-129-5p能夠抑制3T3前體脂肪細胞的增殖過程。另外,過表達miR-129-5p對G3BP1基因mRNA及蛋白水平的表達都具有抑制作用,miR-129-5p直接靶定G3BP1基因的3’UTR區域,并且過表達miR-129-5p使p38的蛋白水平及磷酸化水平表達顯著升高。該實驗證明了miR-129-5p可以通過靶定G3BP1并影響p38通路抑制3T3前體脂肪細胞的增殖過程,與此同時miR-129-5p能夠在分子水平上抑制白色脂肪向棕色化的轉變,但是該過程并不會影響脂肪細胞的分化。該研究成果為miRNA對白色脂肪棕色化的調控研究開拓了新方法并提供了理論依據。
從負向調節棕色化的miRNAs中可以看出,miR-155直接作用于C/EBPβ來負向調節棕色脂肪細胞的分化;miR-150和miR-133都是通過PRDM16這個靶基因的作用途徑來調控棕色化;miR-129-5p則是通過G3BP1的作用途徑,負向調控棕色化。可以見得,這些miRNAs都是通過特定靶基因的作用通路來負向調控白色脂肪棕色化。
4 運動對miRNAs以及白色脂肪棕色化的影響
運動訓練可以改變多種組織器官中miRNAs的表達,調節生理系統的功能。有研究顯示,強度為20 min 75%VO2max的運動能夠致使外周血單核細胞中94個miRNA的表達改變[46]。15m/min、90 min運動導致C57BL/6J小鼠骨骼肌中miR-23的表達降低,使miR-181、miR-1、miR-107的表達顯著升高,說明耐力訓練的適應調節過程可能受miRNA調控[47]。Fernandes等[48]人研究發現,運動訓練可以降低肥胖Zucker大鼠miR-208a表達,使Med13表達升高,調節機體的脂類代謝和能量平衡。有研究為了進一步闡明miRNAs在生物學中的作用,測量了28名中年休閑運動員血漿中的miR-133a[49]。研究人員包括28名中年康樂運動員(其中包括11名女性和17名男性,平均年齡為46歲),他們進行了21.1公里的半程馬拉松比賽。在運動前和運動進行后分別測量血漿中miR-133a的濃度,肌酸激酶(CK)和高敏心肌肌鈣蛋白T(hs-cTnT)的血清濃度。結果發現,miR-133a在半程馬拉松賽后立即大幅增加。結合miR-133a研究可以看出,在正常情況下,miR-133a對白色脂肪棕色化具有負調節的作用,那么半程馬拉松運動賽后miR-133a大幅度增加,使其對白色脂肪棕色化的負調節作用更加明顯。我們可以推測,有的運動通過對miRNAs產生影響,可能對白色脂肪棕色化產生潛在的調節機制。
運動能有效減少脂肪組織體積和甘油三酯的儲存,是世界上公認最安全的肥胖干預手段[50-54]。二十世紀末,人們開始研究運動訓練對棕色脂肪組織的影響,但是研究結果并不完全一致[55,56]。Dinas等[57]人的研究報道了體育活動與棕色脂肪活性呈負相關。同時也有研究顯示,運動訓練對棕色脂肪并沒有明顯的影響。Stan-ford等[58]人研究認為,運動可誘導白色脂肪發生巨大改變,能夠增強線粒體的活性和基因的表達,促使白色脂肪棕色化。Spiegelman實驗室[59]發現有氧運動可使白色脂肪向米色脂肪轉變,從而加速脂肪代謝、促進機體的產熱,有氧運動作為一種健康的生活方式相比于藥物是一種更好的治療方式。國內學者有研究[60]發現,有氧運動對SD大鼠腹內的白色脂肪棕色化作用并不明顯,但是有氧運動能提高SD大鼠的肩胛間棕色脂肪的產熱,促進棕色脂肪細胞分化。另外,付鵬宇等[61]人研究發現,以低氧暴露和運動訓練作為干預手段,能夠提高棕色脂肪的標志蛋白UCP-1和白色脂肪棕色化的作用因子PGC-1αmRNA的相對含量,促進白色脂肪的棕色化,提高機體棕色脂肪細胞的活性。也有研究[62]發現,低氧環境下,PGC-1α表達增加,與PPARγ共同作用,促進白色脂肪組織向棕色化轉變,這一研究與常氧環境下,白色脂肪棕色化的作用相一致。Lu等[63]人的研究揭示了,PGC-1α通過低氧刺激調控脂肪細胞產熱、脂肪酸氧化、糖代謝和脂代謝,并且對脂肪細胞的分化具有重要影響[64]。目前有關常氧運動和低氧運動對白色脂肪棕色化的影響研究較少且存在爭議,今后須進一步驗證不同運動方式對白色脂肪棕色化的確切作用機制[65]。
總體來講,有關運動對miRNAs調控的研究處于起步階段,還需進一步的驗證和探討其作用機制及生物學意義。關于運動對白色脂肪棕色化的影響,不同的研究方案,其結果也存在著差異,針對當前的研究現狀,今后的研究需要更全面地闡明不同運動方式誘導白色脂肪棕色化的作用機制和生理學依據。
5 小結和展望
綜上所述,miRNAs與白色脂肪棕色化的過程聯系緊密,miRNAs可進行棕色化基因轉錄后的調節,且miRNAs對白色脂肪棕色化有正向和負向調節之分。但是,當前只有少量的miRNAs對白色脂肪棕色化的作用機制被闡述(如圖1所示),今后須進一步研究其他miRNAs對白色脂肪棕色化的作用機制。值得注意的是,目前有關miRNAs對白色脂肪棕色化的研究僅在動物實驗和人體細胞培養中得到驗證,其在人體中的確切作用機制還有待進一步的驗證。就疾病而言,基于RNA療法已進入臨床試驗階段,進一步探索白色和棕色脂肪組織中miRNAs的功能將會成為幫助人類治療肥胖癥及其后續并發癥的新療法[66]。另外,關于運動對miRNAs的影響研究還在起步階段,有關于運動對白色脂肪棕色化影響的研究較少且存在爭議,今后的研究還需更加全面地闡明不同運動方式誘導miRNAs和白色脂肪棕色化的作用機制及生理學意義,使該領域的研究系統更加完善,盡可能多的發現能夠有效干預肥胖的miRNAs,促進棕色脂肪的生成以及白色脂肪棕色化,預防和治療人類的肥胖及代謝異常疾病。
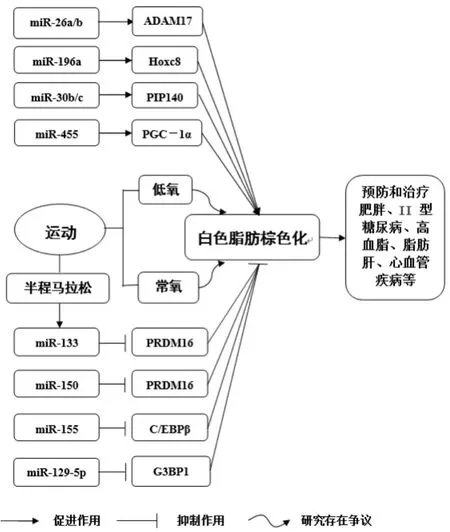
圖1 運動、白色脂肪棕色化、miRNAs三者的作用過程
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
汽車工程學報(2017年2期)2017-07-05 08:13:02
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
