海洋生物固氮研究進展
2021-11-13 03:39:02李志紅李勁尤劉甲星
生態科學 2021年5期
李志紅, 李勁尤, 劉甲星
海洋生物固氮研究進展
李志紅1, 李勁尤2, 劉甲星3, 4, *
1. 珠海萬山海洋開發試驗區海洋漁業科技發展促進中心, 珠海 519005 2. 香港城市大學, 數據科學學院, 香港 3. 中國科學院南海海洋研究所, 海洋生物資源可持續利用重點實驗室, 廣州 510301 4. 南方海洋科學與工程廣東省實驗室(廣州), 廣州 511458
海洋生物固氮因可以支持初級生產所需的氮而在全球碳氮循環中具有重要作用。從二十世紀九十年代分子生物學和15N2同位素示蹤法應用于固氮研究領域以來, 逐漸發現了單細胞固氮藍藻和異養固氮細菌的重要性, 是近年來海洋固氮研究領域的最大進展之一, 表明以前基于束毛藻為主要固氮生物估算的固氮量可能低估了生物固氮在全球海洋生物地球化學循環中的地位。另一方面, 傳統的海洋生物固氮研究僅局限于熱帶亞熱帶的寡營養鹽區域, 對高營養鹽區域如上升流、河口等高營養鹽區域較少關注, 因此有必要對這些區域的生物固氮進行重新評估和再認識。綜述了國際固氮研究的最近進展, 主要包括固氮生物多樣性及分布特征、生物固氮的限制性因素、研究方法以及存在的問題。同時綜述了南海生物固氮方面的最新進展和問題。
海洋生物固氮; 單細胞固氮藍藻; 異養固氮細菌; 分子生物學技術; 高營養區域; 南海
0 前言
海洋固氮生物可以利用氮氣轉化為生物可利用氮, 這一過程叫作生物固氮, 每年固定的新氮(大約100—150 Tg)占全球氮輸出總量一半以上[1]。在貧營養環境中, 生物固氮固定的氮是上層海洋新氮的重要來源, 可以有效緩解上層海洋的氮限制[2]。同時, 對于二氧化碳的凈吸收也起著重要作用[3]。因此, 海洋生物固氮在海洋碳氮循環中發揮著重要作用, 是國際研究的熱點。
自1961年在馬尾藻海發現束毛藻可以固氮以來[4], 由于研究方法的限制, 人們一直認為束毛藻是最重要的海洋固氮生物。直到20世紀90年代分子生物學技術和15N2同位素標記法應用于海洋生物固氮研究, 在對固氮生物種類和分布的認識上取得了一系列突破性的進展, 每年發表的研究論文逐漸增加(圖1a, 表1)。如Zehr等(2001)發現了單細胞固氮藍藻在海洋生物固氮中也起著同樣重要的作用, 隨后也發現異養固氮細菌在某些海域固氮生物群落中占據優勢[5–6], 拓展了人們對于固氮生物的認識[7]。盡管國際上海洋生物固氮相關的研究已經相當多, 但是依然有很多問題尚待探究。相對而言, 中國邊緣海的海洋生物固氮研究薄弱, 直到2000年以后才逐漸有一些研究報道(圖1b), 研究的深度和廣度明顯落后于其他國家。本文在綜述國際上生物固氮領域的最新進展的同時, 以南海為例, 介紹了我國固氮研究的現狀和存在的問題, 期望為中國邊緣海固氮研究做參考。
1 海洋固氮生物多樣性
海洋固氮生物主要包括固氮藍藻和固氮異養細菌, 目前認為固氮藍藻是海洋中主要的固氮生物[1, 24], 包括絲狀固氮藍藻、共生固氮藍藻和單細胞固氮藍藻[25]。其中, 束毛藻(Trichodesmium spp.)是最常見且最重要的一類絲狀固氮藍藻[4, 26], 主要生活在水溫高于20℃的寡營養海域上層[27], 在24℃—30℃度之間最適合固氮, 易在海洋表層形成赤潮[26]。束毛藻固氮具有一定的節律性, 固氮發生在白天。目前發現的共生固氮藍藻主要有: 與根管藻(Rhizosolenia spp.)和半管藻(Hemiaulus spp.)共生的Richelia(分別為Het-1和Het-2), 以及與角毛藻(Chaetocerous)共生的Calothrix (Het-3)28–29], 這些固氮藍藻喜歡生活在營養豐富的近岸區域[7, 29-36]。單細胞固氮藍藻個體一般小于10μm, 在寡營養和富營養區域都有廣泛分布, 且在深水層仍有較高豐度, 其固氮量可達海洋固氮總量的30%—70%, 在某些區域超過束毛藻[37], 主要分為三個類群(UCYN-A, UCYN-B和UCYN-C)[15, 38–40]。UCYN-A尚未得到室內培養, 直徑小于1μm, 固氮發生在白天, 生長溫度范圍為15–30°C, 不能獨立進行光合作用[41], 與進行共生[42–43]; UCYN-B以可在室內培養的為代表, 細胞呈球形, 直徑范圍為3—10μm, 可以獨立進行光合作用, 固氮一般在晚上進行, 最適生長溫度為22—36°C[15, 38, 44–45]; UCYN-C以可培養的為代表, 可以進行光合作用, 固氮也在晚上發生[46]。此外, 也發現一些異養細菌含有固氮基因(), 如α、β和γ變形菌等[6, 7, 47–50], 盡管在南太平洋、印度洋和南海北部區域報道了異養細菌是主要的固氮生物[5–6, 51–53],但是其固氮活性重要性尚無法判斷[7]。
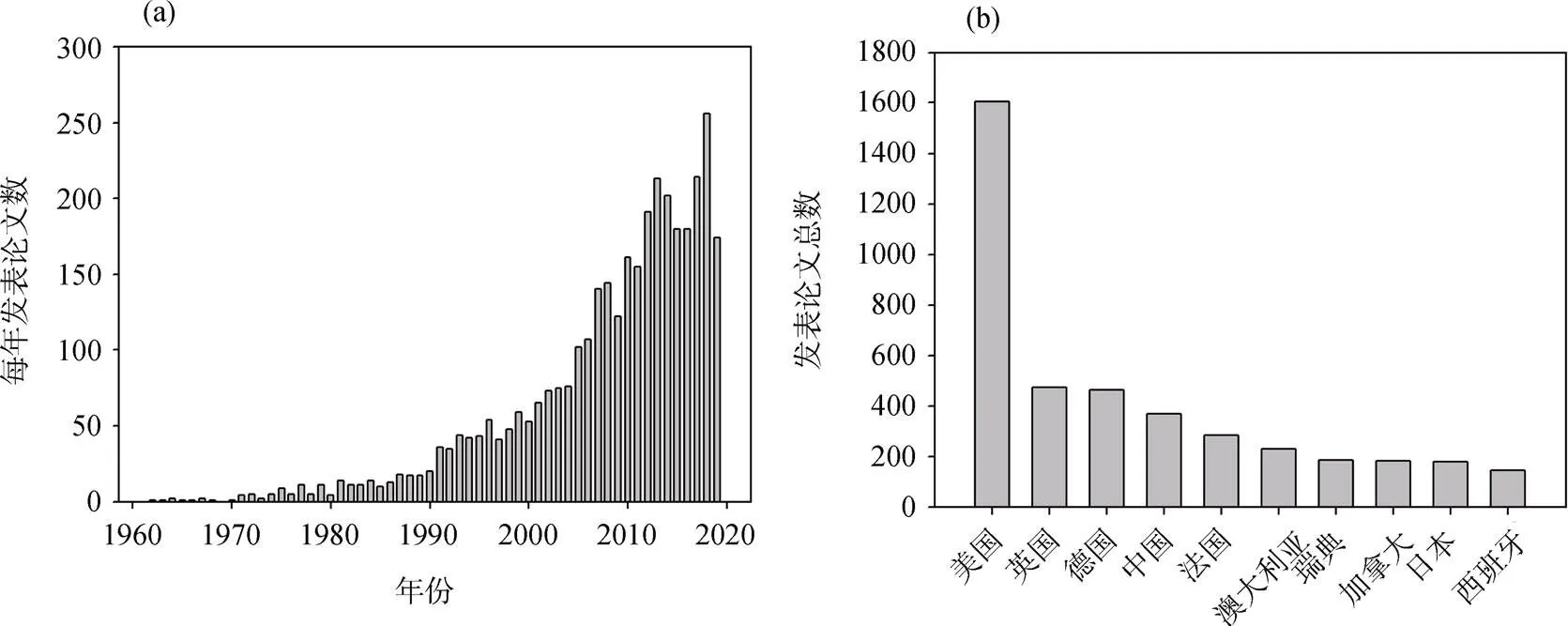
圖1 Web of science檢索到的海洋生物固氮年發表文章數(a)與國家發表論文數(b)
Figure 1 Number of published articles on marine nitrogen fixation every year (a) and national publications (b) retrieved by web of science
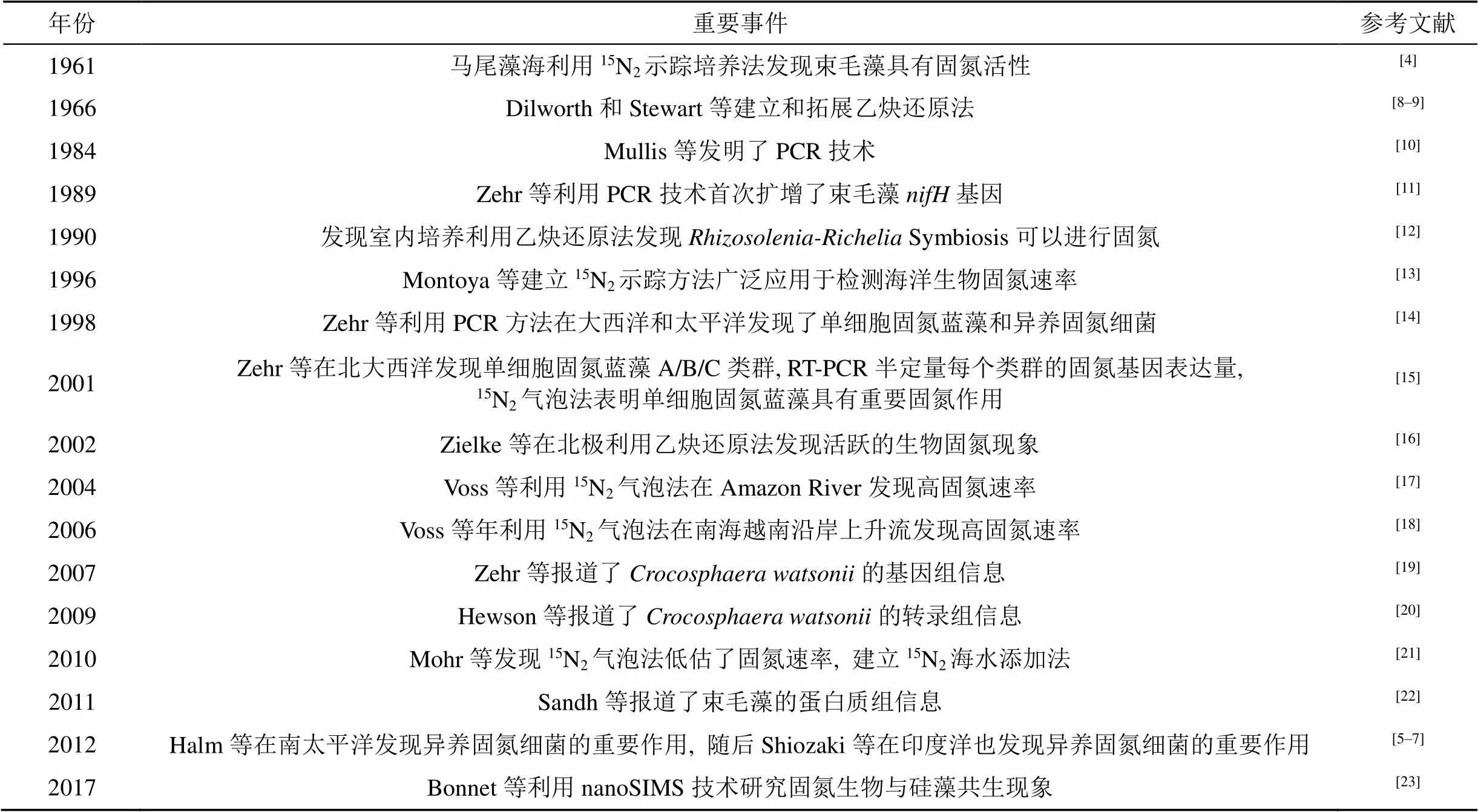
表1 海洋生物固氮研究歷史中的重要事件
2 海洋生物固氮的影響因素
2.1 溫度、鹽度、光和氧氣
溫度可以影響固氮生物的生長和固氮, 因此影響著固氮生物的分布[54–55]。野外調查的結果表明, 束毛藻可以生活在20—30℃之間的海域, 最適溫度為25—30℃[26–27, 56–57]; 室內培養實驗也表明, 20—34℃為束毛藻生長和固氮的溫度范圍, 最適范圍為24—30℃, 在27℃時最活躍[58]。單細胞固氮類群似乎更能耐受更低的溫度, 如可以生活在18—30℃[59], 在溫度較低區域也很常見[39–40]。另外, 鹽度也可以影響固氮生物生長和固氮, 束毛藻有較廣泛鹽度耐受性(22—43), 33—37是最適固氮鹽度區間[60]; 與共生固氮藍藻可以在較低的鹽度生存, 所以經常在河口等近岸[29, 61]。
光可以影響固氮酶的活性, 同時也影響細胞的光合作用, 因此也能影響固氮生物的生長和固氮[62]。束毛藻一般在表層豐度較高, 因為其對光的需求較高, 光強增加可以促進固氮速率增加[63], 西太平洋的現場培養實驗表明束毛藻的光飽和強度在400 μmolquanta·m-2·s-1左右[64]; 室內實驗也表明, 束毛藻在670 μmol quanta·m-2·s-1光照強度下還可以快速生長(生長速率可以達到0.49 d-1)[65]。光強對單細胞固氮藍藻同樣也具有一定的影響, 如室內實驗表明Crocosphaera在180 μmol quanta·m-2·s-1光強下生長速率和固氮速率隨著光照強度增加而增加, 在300 μmol quanta·m-2·s-1光照條件下生長速率和固氮速率沒有受到明顯抑制[66]; 當溫度在14-30°C范圍內時,在60 μmol quanta·m-2·s-1可以達到光飽和狀態[59]。另外, 光照周期也會對固氮藍藻造成影響, 12h光照/12h黑暗的光照周期條件下Cyanothece固氮速率最大, 延長或者縮短光照時間都明顯抑制固氮速率[67]。除了光強和光周期, 不同光譜也會影響固氮生物的生長, 室內培養實驗表明紫外線可以抑制束毛藻生長和固氮, 但束毛藻在受到紫外線激發時產生類菌胞素氨基酸MAAs以減少紫外線的抑制作用[68]。
此外, 氧氣也能抑制固氮酶活性, 因此固氮生物采取多種機制來避免光合作用產生的氧氣對生物固氮的抑制[1, 69–70]。具有異形胞的絲狀固氮藍藻固氮作用發生在異形胞內部, 異形胞是分化的細胞, 不能進行光合作用, 但是可以進行固氮作用, 這一特定分化的細胞使整個絲狀固氮藍藻的光合作用和固氮作用形成區域分割, 有效的避免了氧氣對固氮酶的抑制[26]。沒有異形胞的絲狀藍藻束毛藻, 也可以在白天進行固氮作用, 可能與其獨特的細胞結構或者固氮酶特征有關[26]。對于單細胞固氮藍藻, UCYN-A細胞營共生生活, 由于細胞缺乏光系統而不能進行光合作用, 因此在白天進行固氮[41]; 而對于UCYN-B和UCYN-C而言, 固氮光合作用在晚上發生, 以有效避免氧氣對固氮酶的抑制[15, 38, 44–46]。
2.2 營養元素
鐵參與固氮酶復合體的形成, 因此, 固氮生物對鐵的需求量要高于其他浮游植物[71]。在寡營養的大洋區域, 鐵主要來源于大氣沉降和水體的垂直或水平輸送, 大氣干濕沉降的鐵對上層水體的固氮生物生長和固氮起著至關重要的作用[72]。已經在一些區域得到證實, 如南大西洋固氮速率與溶解性鐵成正相關而與磷負相關[73]; 在邊緣海也有鐵限制固氮生物的報道, 如在南海海盆區域, 盡管大氣沉降帶來大量鐵, 但依然存在缺乏配體而導致生物可利用鐵缺乏[74]; 基于原位分子生物學的結果也表明大西洋和太平洋一些區域鐵是束毛藻生長和固氮的限制性元素[75]。原位培養實驗也表明, 鐵是西北大西洋固氮生物,和固氮的限制性因素[76]。室內培養實驗也發現鐵缺乏可抑制束毛藻固氮[77–79], 缺鐵也可以對生長和固氮產生不利影響[80–81]。
磷在參與細胞構建以及核酸、蛋白質等大分子物質合成、能量傳遞等方面起著重要作用, 同時也對固氮生物的生長和固氮等生理過程具有重要影響。很多原位觀測結果證實了這一結論, 如基于無機磷濃度的結果表明西北大西洋上層固氮生物生長可能受到磷限制[82]; 基于堿性磷酸酶和固氮速率的直接證據表明北大西洋海盆區域束毛藻生長受到磷限制[83]; 基于堿性磷酸酶、鐵結合蛋白基因和固氮速率的觀測結果表明西南大西洋生物固氮的限制性因子可能為磷[84]; 基于固氮速率和束毛藻豐度的結果表明南海瓊東上升流影響區域磷可能是生物固氮的限制性因子[85]。同時, 原位培養實驗的結果也證明了許多區域存在磷限制, 如波羅的海添加磷促進水體總固氮速率[86], 西南大西洋的實驗表明添加磷也有效促進了束毛藻的生長和固氮[84], 東北大西洋區域的現場加富實驗表明該區域磷是限制固氮生物,和固氮的主要因素[76]。為了適應磷匱乏的環境, 固氮生物進化了許多生存策略, 如束毛藻可以通過調節浮力在上層水柱進行垂直遷移獲得磷[87], 同時可以在環境磷濃度低時高表達高親和磷蛋白等相關基因以高效獲取磷, 此外, 束毛藻具有分解有機磷的相關基因如和家族基因, 通過高表達堿性磷酸酶等水解有機磷來滿足其生存[88]; UCYN-B也具有高親臨蛋白基因和分解有機磷相應的基因來應對磷缺乏[89–90]; 盡管UCYN-A基因組缺乏分解有機磷相關的基因和代謝途徑[43], 但是其能與單細胞藻類進行共生[42–43], 這種關系可以使UCYN-A在低磷環境中生存[76]。
此外, 某些區域也存在磷和鐵協同限制固氮生物生長的情況, 如北大西洋的現場加富實驗表明, 固氮作用受到磷和鐵的協同限制, 撒哈拉沙塵所攜帶的大量磷和鐵共同刺激了該海域的固氮作用[91], 北大西洋赤道附近區域的現場加富實驗也表明該區域磷和鐵可能是協同限制固氮生物,和固氮[76]。
盡管鉬也是固氮酶復合體的一個重要組成元素[92], 但在原位環境中報道鉬限制固氮生物生長的研究較少, 可能是因為自然海水中具有較高的鉬。此外, 溶解無機氮也會影響固氮生物固氮[1], 如束毛藻和也可以利用無機氮源, 但是固氮會被抑制[93–94]。另外, 也有研究表明環境中氮磷的比例也會對固氮造成一定影響[95–96]。
2.3 物理過程
2.3.1 中尺度渦
中尺度渦是一種重要的海洋動力學現象, 常見的中尺度渦旋分為三類, 氣旋冷渦(cyclonic eddy), 反氣旋暖渦(anticyclonic eddy)和模式水渦(Mode- water eddy), 其中冷渦一般導致中心水團向上涌升, 暖渦導致中心水團向下沉降, 模式水渦在一般導致中心水團上層涌升而下層沉降[97]。大量研究表明中尺度渦可以通過垂直輸運水團改變水體的理化特征和營養鹽等來影響浮游植物群落組成, 因而在全球碳循環中發揮著重要作用[98–99]。一方面, 中尺度渦可以通過物理混合作用影響固氮生物, 如水團混合導致束毛藻固氮酶活降低[100], 水體層化系數與束毛藻豐度負相關[101]。另一方面, 中尺度渦可以通過影響溫度、鹽度、營養鹽等環境因子來調控固氮生物的分布及其固氮(表2)。已有不少研究表明反氣旋渦可以促進固氮, 如北大西洋反氣旋渦內束毛藻豐度較高[102–103], 北太平洋反氣旋渦內固氮藍藻(spp.,和)的豐度及總固氮速率都高于臨近海域[104–105], 澳大利亞西海岸反氣旋渦內固氮速率是氣旋渦內的2.5倍[106], 地中海區域反氣旋渦內固氮速率是臨近海域的10倍[107], 南太平洋的研究也表明反氣旋渦影響區域最大固氮速率出現在真光層以下, 且反氣旋渦刺激的固氮作用在向近岸區域輸送氮方面起著重要作用[108], 南海東北部暖渦區域固氮速率顯著高于臨近參考區域和冷渦區域, 同時UCYN-A和UCYN-B豐度較高[109]。相對而言, 冷渦區域的固氮報道的較少, 有限的結果表明冷渦也可以影響固氮, 如南太平洋的研究也表明, 氣旋渦影響區域固氮僅發生在表層[108], 南海東北部冷渦區域固氮速率和固氮藍藻豐度(UCYN-B 和Trichodesmium)低于臨近區域[109], 南海海盆區氣旋渦區域發現高豐度的固氮異養細菌[110]。這些研究表明中尺度渦可能在影響生物固氮方面起著重要作用, 同時可以明顯調控固氮對初級生產力所需氮的貢獻, 如南海東北部反氣旋渦可以顯著刺激生物固氮速率, 反氣旋渦內固氮對初級生產力所需氮的貢獻率可以達到9%左右[109]。
2.3.2 上升流等高營養鹽區域生物固氮
沿岸上升流作為一種重要的中尺度物理現象, 其可以通過改變營養鹽和浮游植物群落結構而對海洋生態系統具有重要影響[113-114]。一般認為, 沿岸上升流區域不存在或具有較弱的生物固氮作用。首先, 沿岸上升流區域有較高的氨氮和硝氮, 固氮生物不會耗費大量能量固氮(固氮過程需要消耗大量能量)[115]; 其次, 固氮生物在與其他浮游植物生長競爭中也常處于劣勢地位[116]; 另外, 上升流區較低的溫度似乎也不適合喜高溫環境的固氮生物生長[1]。因此, 沿岸上升流區域的生物固氮研究一直沒有得到學者們的重視。然而, 隨后在如亞馬遜河口和湄公河口等高營養區域也發現較強的固氮作用[31, 61]。一般認為固氮生物喜歡在高溫高鹽寡營養鹽的低緯度地區, 然而在上升流、河口及沖淡羽等富營養鹽海區也發現生物固氮相當活躍[18, 29, 31, 34–35, 40, 106, 116–121]。
盡管已有少量研究報道上升流對固氮生物及其固氮的影響, 然而, 對上升流區域的生物固氮分布模式及其形成機制的認識仍然十分有限。首先, 上升流區域是否存在生物固氮作用依然存在爭議, 如在南太平洋的本格拉(Benguela)上升流區域檢測到較高的固氮速率[25]; 但另外的研究認為, 該區域在上升流季節并不存在生物固氮現象, 原位培養實驗也證實了這一結論[120]。其次, 上升流對固氮生物生長及其固氮影響的機制存在爭議, 且固氮生物群結構落數據相對缺乏。生物固氮一般受到磷或鐵的限制, 上升流帶來的磷或鐵可以刺激固氮生物生長及其固氮, 如在越南沿岸上升流和湄公河帶來的大量磷和金屬使該區域出現高固氮速率, 但沒有直接檢測固氮生物群落結構[18]; 大西洋赤道上升流帶來的磷和鐵可以促進該區域的生物固氮, 推測單細胞固氮藍藻對該區域固氮具有明顯貢獻, 但也沒有直接檢測固氮生物類群[116]; 在南海的瓊東上升流區域, 相對較高的固氮速率可能與上升流帶來的磷和鐵有關[85]。另一些研究認為上升流區域出現高固氮速率可能與有機物質和低氧有關。如在南太平洋上升流區域, 400米處依然檢測到生物固氮作用, 且缺氧的次表層固氮速率是真光層的5倍之多, 在上層水體中沒有發現固氮藍藻, 卻存在大量異養固氮細菌, 低氧可能是該區域高固氮的重要因素[122]; 在東南太平洋的智利上升流區域, 低氧和有機物質可能是上升流區域出現固氮的主要因素, 同時浮游植物群落結構及其碳氮比也會影響固氮速率, 且異養固氮細菌對該區固氮起著重要作用[123]。這些結果表明上升流對生物固氮影響的機制尚不清楚, 值得繼續探究。

表2 中尺度渦對固氮生物群落結構和生物固氮速率影響的報道
3 海洋生物固氮的研究方法
早在1961年, Dugdale等就利用15N2示蹤培養法發現束毛藻具有固氮活性[4]。通過添加15N標記的氮氣, 利用元素同位素質譜儀檢測培養后的顆粒有機氮, 就能示蹤結合到顆粒有機氮的15N, 進而可以計算生物固氮速率。該方法靈敏度高, 是目前檢測海洋固氮速率的常用方法[13]。該方法最早通過向培養體系中直接添加15N同位素標記的氮氣來實現, 被稱為氣泡法[13], 后來有研究表明氣泡法的結果可能低估了實際固氮速率[21, 124], 因此又發展出了海水添加法[125]。
Dilworth等在1966年發現固氮酶可以將乙炔還原為乙烯[9], 隨后Stewart等在1967年將乙炔還原法擴大應用于湖泊等野外環境中[8], 此后該方法一直沿用至今。該方法的原理為, 在同時存在乙炔和氮氣的情況下, 固氮酶優先把乙炔還原為乙烯[9], 一般認為被還原的乙烯與被固定的氮氣存在固定的比例即3:1, 以此便可以估算出固氮速率[8]。但是該方法的也有一定的局限性[126], 因為不同環境下乙烯和氮氣之間的比值不盡相同[127]。
盡管同位素法和乙炔還原法可以有效的檢測環境中的總固氮速率, 但是隨著越來越多的固氮生物被發現, 檢測環境中每個類群的固氮活性已經成為迫切。自1989年Zehr等利用PCR技術首次擴增了束毛藻基因以來, 分子生物學方法逐漸應用到海洋生物固氮的研究中。固氮酶一般由含鐵還原酶(N2reducatse)和含鉬鐵固氮酶(N2dinitrogenase)組成, 分別由和基因編碼, 此外還有一些含釩離子的固氮酶[128], 其中基因序列高度保守, 功能信息比的更豐富, 且其表達受到時間、空間、光照和營養鹽影響, 因此序列是鑒定固氮生物物種及分析固氮活性的重要依據[129]。大部分固氮生物的拷貝數目相對穩定, 可以基于的拷貝數來估算某一類群固氮生物的豐度和揭示固氮生物群落結構信息[40–41]。同時, 利用基因表達的水平可以推測某一類固氮生物類群的固氮活性。因此分子生物學方法是研究固氮生物群落結構及固氮活性的好方法[130], 尤其隨著定量PCR技術、二代測序技術、基因組、轉錄組和蛋白質組技術的發展, 基于海量數據的大數據分析可以得到更加全面豐富的信息, 以便于從基因水平上揭示固氮生物對環境的適應性機制, 該方法是目前研究固氮生物的重要方法。
分子生物學方法盡管可以檢測原位環境中固氮基因或者固氮基因表達量, 然而固氮基因表達量在某些情況下并不代表生物固氮的活躍程度, 因此仍然不能直接證明原位環境中的固氮量問題。熒光原位雜交-納米二次離子質譜技術(FISH-NanoSIMS), 即把熒光原位雜交技術與二次離子質譜技術聯用能夠實現在原位上同時觀察自然環境樣品中微生物的形態結構、種類分布及功能代謝信息[131]。近年來已經普遍用于定量單細胞固氮速率和共生固氮固定氮轉移問題的研究中, 展現了研究原位環境固氮的強大優勢[33, 42]。
4 南海生物固氮的研究現狀
南海大部分區域具有強層化、寡營養、溫暖和強大氣沉降的特征, 具有利于固氮生物生長的良好環境[25, 74]。氮是限制南海外海浮游植物生長的關鍵因子[132], 因此固氮對南海的碳氮生物地球化學循環具有重要作用。盡管南海生物固氮的研究起步較晚, 但近年來取得了一系列新成果。
在對固氮生物多樣性的認識上, 束毛藻是南海常見的固氮類群, 普遍分布于珠江口影響區域[133]、三亞灣[134]、大亞灣[135]、瓊東上升流區[85]、粵東上升流區[136]、湄公河河口和越南沿岸等近岸區域[18, 61, 137–138], 也在呂宋海峽及鄰近南海海盆[109, 139–141]和南沙島礁等離岸區域廣泛存在[142], 同時在某些區域如大亞灣[135, 143]和粵東沿岸區域[85]附近經常發生束毛藻赤潮; 單細胞固氮藍藻UCYN-A和UCYN-B在南海東北部區域普遍存在[96, 109, 133, 141, 144], 在呂宋海峽區域附近也發現了較高豐度且對總固氮量的貢獻可達65%以上[145]; 在珠江口(Het-1)[133]、粵東上升流(Het-1)[96]和湄公河河口(Het-1和het-2)[18, 61, 138]等近海域存在與硅藻共生的固氮生物; 另外, 異養固氮細菌也廣泛存在于大亞灣、南海北部和海盆區域[109, 133, 146–149]。
相對于固氮生物的研究而言, 直接檢測固氮速率的研究相對較少[32]。已有發表的結果表明, 臺灣海峽為11—40 μmol N·m-2·d-1[150], 秋季南海東北部近岸區域表層固氮速率范圍為4—213 nmol N·m–3·h–1[143], 大亞灣四個季節固氮速率為0—4.51 nmol N·L-1·h-1[149], 夏季瓊東上升流區域為7.5—163 μmol N·m-2·d-1[85], 夏季粵東上升流區域為0—7.51 nmol N·L-1·d-1[136], 南海東北部海盆區四個季節固氮速率為2.4—168.1 μmol N·m-2·d-1, 夏季出現最高固氮速率, 其次為春季和秋季, 冬季固氮速率最低[101, 109]。湄公河河口固氮速率為5.05—22.77 nmol N·L-1·h-1[61], 越南沿岸區域固氮速率為1.9—190.6 μmol N·m-2·d-1[18]。
在固氮生物分布的區域上, 除了傳統上認為適合固氮生物生活的高溫高鹽寡營養鹽的低緯度區域發現固氮生物以外, 也在河口、上升流和海灣等高營養也存在較高豐度的固氮生物, 如在湄公河河口束毛藻和硅藻共生固氮藻(Het-1和Het-2)出現高豐度和高固氮量[61, 137–138]; 在珠江口影響區域, 束毛藻在靠近外海站位相對較多[133], 同時珠江口影響的局部區域發生束毛藻赤潮和高固氮速率[143]。在越南沿岸上升流區域, 且在季風期間固氮速率是季風轉換期間固氮速率的10倍左右[18]; 在瓊東上升流區域也發現活躍的生物固氮作用, 上升流帶來的鐵可能刺激該區域固氮, 且局部出現束毛藻赤潮[85]; 在粵東上升流, 也發現上升流影響區域存在活躍固氮能力, 同時發現大量固氮藍藻(束毛藻、UCYN-A和Het-1)[96]。同時, 在營養豐富的大亞灣等水域也經常會發生固氮生物束毛藻赤潮現象, 如1987年8月大亞灣海域的發生大面積束毛藻赤潮; 2004年6月大亞灣灣口發生束毛藻赤潮, 呈黃褐色“彩帶”, 寬3海里、長8海里, 面積約為100平方公里[135]; 2007年深圳大鵬灣梅沙赤潮, 主要為漢氏束毛藻[151]; 2016年大亞灣灣外也檢測到肉眼可見的束毛藻赤潮, 同時實測固氮速率異常高[143]; 大亞灣開展的研究也同時表明大亞灣區域高營養區域并沒有限制固氮速率, 在某些區域反而出現較高固氮速率[149]。以上結果表明, 在高營養的近岸區域也會發生明顯生物固氮現象, 與以往固氮生物尤其固氮藍藻喜歡生活在貧乏營養狀況下的結論不一致, 值得深入探究。
在對固氮生物生長限制性因子的認識上, 相關研究較少且存在爭議。在南海海盆區盡管大氣沉降可以輸送充足的鐵, 但是由于鐵配體不足的致使生物可利用鐵濃度較低(溶解性鐵濃度僅0.2–0.3 nM), 推測固氮生物生長的限制性因子可能為鐵[74], 但是并沒有在該區域直接實驗證明, 盡管北部灣冬季灣口的現場加富培養實驗表明生物固氮作用可能受到鐵限制[152], 但是在南海北部區域基于痕量金屬潔凈技術采樣發現南海北部區域鐵較為充足(0.50 nM), 可能不會限制浮游生物的生長[153], 但是固氮生物生長對鐵的需求其實要高于其他浮游植物[71], 并不足以說明鐵不是限制因子。粵西瓊東上升流區域的研究則認為, 上升流帶來大量鐵, 而磷卻被浮游植物快速利用而缺乏, 最終該區域表現為磷限制[85]。
南海不同海域固氮作用對初級生產力所需氮的貢獻有很大差別, 如越南沿岸區域固氮可以支持初級生產力氮需求的0.1—8.2%[18, 154], 而湄公河河口區域貢獻率可達47%[61], 三亞灣束毛藻固氮量對初級生產氮貢獻率為0.03—1.63%[134], 而粵西瓊東上升流海區貢獻率僅為0.01—2.52%[85], 南海北部海盆區域暖渦影響區域該貢獻率最高可以達到9%[109]。
5 結論與展望
海洋生物固氮研究的發展與研究技術方法密切相關, 技術的突破是研究突破的基礎, 如從1961年束毛藻發現到1989年PCR技術應用于海洋固氮研究領域之間將近30年間, 海洋生物固氮的研究并沒有取得里程碑式的進展。然而, 二十世紀九十年代分子生物學和15N2同位素示蹤法應用于固氮研究領域, 取得了一些突破性進展。在空間分布方面, 生物固氮的研究從傳統上認為寡營養鹽的低緯度地區拓展到上升流、河口及海灣等富營養鹽海域[18, 29, 31, 34–35, 40, 106, 116–121], 從高溫高鹽的熱帶亞熱帶低緯度區域到高緯度寒冷海域[16, 155–156], 從海洋表層到上千米的深層和沉積物[40, 157]。在對固氮生物多樣性的認識方面, 從束毛藻到單細胞固氮藍藻的發現[15, 32, 34, 38–39, 40, 42, 106, 158–159], 再到發現異養固氮細菌的重要性[6–7, 48–50, 160–161]。這些發現, 拓展了對固氮生物及其固氮的傳統認識, 表明以前基于大洋寡營養鹽區域束毛藻為主要固氮生物估算的固氮量可能低估了生物固氮在全球海洋生物地球化學循環中的地位[38–44]。
固氮作用與反硝化作用分別是海洋氮最重要的源與匯, 這兩個過程決定了海洋氮庫的收支平衡[162]。然而, 基于已發表的實測固氮速率的數據顯示, 海洋中反硝化速率超過固氮速率, 結合態氮的虧損速率高達近200 Tg N·yr-1[163]。但是, 基于模型和沉積物同位素的結果表明, 全球氮收支處于相對平衡狀態[164–165], 這就引起人們重新反思對全球海洋生物固氮的再認識, 是否過去的方法存在低估的情況[166]。首先, 在檢測方法上, 過去應用廣泛的15N2同位素示蹤氣泡方法比新的海水添加法低估了固氮速率[13, 21], 如在束毛藻為主要固氮者的海域, 該方法可能低估了62%的固氮速率, 而在單細胞固氮藍藻、共生固氮藍藻和異養固氮菌為主導的固氮生物群落結構中, 基于海水添加法的固氮量結果是氣泡法的6倍之多[124]。基于氣泡法和海水添加法的計算世界大洋固氮速率可以從原先估計的103± 8Tg N·yr-1上升到177±8 Tg N·yr-1[124]。其次, 過去對全球海洋生物固氮量的估計都是基于已發現的大洋區域, 較少涉及到近岸如河口、上升流和海灣等高營養鹽區域, 高營養鹽區域如河口、上升流和沿岸區域是否具有一些曾被忽視的關鍵類群?以及適應這種環境的機制是什么?迫切需要用現代分子生物學方法繼續探究其機制, 有必要對這些區域的生物固氮進行重新評估和再認識。第三, 盡管發現了異養固氮細菌在某些區域占據優勢地位[5, 6, 51–53], 但是其固氮活性至今并沒有進行有效的評估, 而且一般都不認為異養固氮細菌對固氮具有重要性[7]。相信隨著NanoSIMS技術的應用, 這一問題可能會得到一定程度的認知。同時, 近年來組學技術(基因組、轉錄組、蛋白質組和代謝組)也逐漸應用到海洋生物固氮領域, 可以從基因和代謝網絡水平上了解固氮生物與環境之間適應的內在機制。
[1] KARL D, MICHAELS A, BERGMAN B, et al. Dinitrogen fixation in the world's oceans[J]. Biogeochemistry, 2002, 57(1): 47–98.
[2] GLIBERT P M, HEIL C A, HOLLANDER D, et al. Evidence for dissolved organic nitrogen and phosphorus uptake during a cyanobacterial bloom in Florida Bay[J]. Marine ecology Progress series, 2004, 280: 73–83.
[3] ALTABET M A, FRANCOIS R, MURRAY D W, et al. Climate-related variations in denitrification in the Arabian sea from sediment15N/14N ratios[J]. Nature, 1995, 373(6514): 506–509.
[4] DUGDALE R C, MENZEL D W, RYTHER J H. Nitrogen fixation in the Sargasso sea[J]. Deep-Sea Research, 1961, 7(4): 297–300.
[5] HALM H, LAM P, FERDELMAN T G, et al. Heterotrophic organisms dominate nitrogen fixation in the South Pacific Gyre[J]. Isme Journal, 2012, 6(6): 1238–1249.
[6] SHIOZAKI T, IJICHI M, KODAMA T, et al. Heterotrophic bacteria as major nitrogen fixers in the euphotic zone of the Indian Ocean[J]. Global Biogeochemical Cycles, 2014, 28(10): 1096–1110.
[7] TURK-KUBO K A, KARAMCHANDANI M, CAPONE D G, et al. The paradox of marine heterotrophic nitrogen fixation: abundances of heterotrophic diazotrophs do not account for nitrogen fixation rates in the Eastern Tropical South Pacific[J]. Environmental Microbiology, 2014, 16(10): 3095–3114.
[8] STEWART W, FITZGERALD G, BURRIS N. In situ studies on N2fixation using the acetylene reduction technique[J]. Proceedings of the National Academy of Sciences of the United States of America, 1967, 58(5): 2071.
[9] DILWORTH M J. Acetylene reduction by nitrogen-fixing preparations from[J]. Biochi-mica et Biophysica Acta (BBA) - General Subjects, 1966, 127(2): 285–294.
[10] MULLIS K B, FALOONA F A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction[J]. Methods in Enzymology, 1987, 155: 335–350.
[11] ZEHR J P, MCREYNOLDS L A. Use of degenerate oligonucleotides for amplification of thegene from the marine cyanobacterium[J]. Applied and Environmental Microbiology, 1989, 55(10): 2522–2526.
[12] VILLAREAL T A. Laboratory culture and preliminary characterization of the nitrogen-fixing-symbiosis[J]. Marine Ecology, 1990, 11(2): 117– 132.
[13] MONTOYA J P, VOSS M, KAHLER P, et al. A Simple, high-precision, high-sensitivity tracer assay for nitrogen fixation[J]. Applied and Environmental Microbiology, 1996, 62(3): 986–993.
[14] ZEHR J P, MELLON M T, ZANI S. New nitrogen-fixing microorganisms detected in oligotrophic oceans by amplification of nitrogenase () genes[J]. Applied and Environmental Microbiology, 1998, 64(9): 3444–3450.
[15] ZEHR J P, WATERBURY J B, TURNER P J, et al. Unicellular cyanobacteria fix N2in the subtropical North Pacific Ocean[J]. Nature, 2001, 412(6847): 635–638.
[16] ZIELKE M, EKKER A S, OLSEN R A, et al. The influence of abiotic factors on biological nitrogen fixation in different types of vegetation in the High Arctic, Svalbard[J]. Arctic, Antarctic, and Alpine Research, 2002, 34(3): 293–299.
[17] VOSS M, CROOT P, LOCHTE K, et al. Patterns of nitrogen fixation along 10ON in the tropical Atlantic[J]. Geophysical Research Letters, 2004, 31(23): L23S09.
[18] VOSS M, BOMBAR D, LOICK N, et al. Riverine influence on nitrogen fixation in the upwelling region off Vietnam, South China Sea[J]. Geophysical Research Letters, 2006, 33(7): L07604.
[19] ZEHR J P, BENCH S R, MONDRAGON E A, et al. Low genomic diversity in tropical oceanic N2-fixing cyanobacteria[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(45): 17807–17812.
[20] HEWSON I, PORETSKY R S, BEINART R A, et al. In situ transcriptomic analysis of the globally important keystone N2-fixing taxon[J]. The ISME Journal, 2009, 3(5): 618–631.
[21] MOHR W, GROSSKOPF T, WALLACE D W R, et al. Methodological underestimation of oceanic nitrogen fixation rates[J]. Plos One, 2010, 5(9): e12583.
[22] SANDH G, RAN L, XU L, et al. Comparative proteomic profiles of the marine cyanobacteriumIMS101 under different nitrogen regimes[J]. Proteomics, 2011, 11(3): 406–419.
[23] BONNET S, BERTHELOT H, TURK‐KUBO K, et al. Diazotroph derived nitrogen supports diatom growth in the South West Pacific: a quantitative study using nanoSIMS[J]. Limnology and Oceanography, 2016, 61(5): 1549–1562.
[24] ZEHR J P, KUDELA R M. Nitrogen cycle of the open ocean: from genes to ecosystems[J]. Annual Review of Marine Science, 2011, 3(1): 197–225.
[25] SOHM J A, WEBB E A, CAPONE D G. Emerging patterns of marine nitrogen fixation[J]. Nature Reviews Microbiology, 2011, 9(7): 499–508.
[26] CAPONE D G, ZEHR J P, PAERL H W, et al., a globally significant marine cyanobacterium[J]. Science, 1997, 276(5316): 1221–1229.
[27] LAROCHE J, BREITBARTH E. Importance of the diazotrophs as a source of new nitrogen in the ocean[J]. Journal of Sea Research, 2005, 53(1-2): 67–91.
[28] CHURCH M J, JENKINS B D, KARL D M, et al. Vertical distributions of nitrogen-fixing phylotypes at Stn ALOHA in the oligotrophic North Pacific Ocean[J]. Aquatic Microbial Ecology, 2005, 38(1): 3–14.
[29] FOSTER R A, SUBRAMANIAM A, MAHAFFEY C, et al. Influence of the Amazon River plume on distributions of free-living and symbiotic cyanobacteria in the western tropical north Atlantic Ocean[J]. Limnology and Oceanography, 2007, 52(2): 517–532.
[30] CARPENTER E J, MONTOYA J P, BURNS J, et al. Extensive bloom of a N2-fixing diatom/cyanobacterial association in the tropical Atlantic Ocean[J]. Marine Ecology Progress Series, 1999, 185: 273–283.
[31] SUBRAMANIAM A, YAGER P L, CARPENTER E J, et al. Amazon River enhances diazotrophy and carbon sequestration in the tropical North Atlantic Ocean[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(30): 10460–10465.
[32] LUO Y W, DONEY S C, ANDERSON L A, et al. Database of diazotrophs in global ocean: abundance, biomass and nitrogen fixation rates[J]. Earth System Science Data, 2012, 4(1): 47–73.
[33] FOSTER R A, KUYPERS M M M, VAGNER T, et al. Nitrogen fixation and transfer in open ocean diatom-cyanobacterial symbioses[J]. Isme Journal, 2011, 5(9): 1484–1493.
[34] SOHM J A, HILTON J A, NOBLE A E, et al. Nitrogen fixation in the South Atlantic Gyre and the Benguela Upwelling System[J]. Geophysical Research Letters, 2011, 38: L16608.
[35] YEUNG L Y, BERELSON W M, YOUNG E D, et al. Impact of diatom-diazotroph associations on carbon export in the Amazon River plume[J]. Geophysical Research Letters, 2012, 39: L18609.
[36] EICHNER M, ROST B, KRANZ S A. Diversity of ocean acidification effects on marine N2fixers[J]. Journal of Experimental Marine Biology and Ecology, 2014, 457(2014): 199–207.
[37] LUO Y W, LIMA I D, KARL D M, et al. Data-based assessment of environmental controls on global marine nitrogen fixation[J]. Biogeosciences, 2014, 11(3): 691–708.
[38] MONTOYA J P, HOLL C M, ZEHR J P, et al. High rates of N2fixation by unicellular diazotrophs in the oligotrophic Pacific Ocean[J]. Nature, 2004, 430(7003): 1027–1031.
[39] LANGLOIS R J, LAROCHE J, RAAB P A. Diazotrophic diversity and distribution in the tropical and subtropical Atlantic ocean[J]. Applied and Environmental Micro-biology, 2005, 71(12): 7910–7919.
[40] MOISANDER P H, BEINART R A, HEWSON I, et al. Unicellular cyanobacterial distributions broaden the oceanic N2fixation domain[J]. Science, 2010, 327(5972): 1512–1514.
[41] ZEHR J P, BENCH S R, CARTER B J, et al. Globally distributed uncultivated oceanic N2-Fixing cyanobacteria lack oxygenic photosystem II[J]. Science, 2008, 322(5904): 1110–1112.
[42] THOMPSON A W, FOSTER R A, KRUPKE A, et al. Unicellular cyanobacterium symbiotic with a single-celled eukaryotic alga[J]. Science, 2012, 337(6101): 1546–1550.
[43] TRIPP H J, BENCH S R, TURK K A, et al. Metabolic streamlining in an open-ocean nitrogen-fixing cyanobac-terium[J]. Nature, 2010, 464(7285): 90–94.
[44] FALCON L I, CARPENTER E J, CIPRIANO F, et al. N2fixation by unicellular Bacterioplankton from the Atlantic and Pacific oceans: Phylogeny and In Situ Rates[J]. Applied and Environmental Microbiology, 2004, 70(2): 765–770.
[45] WEBB E A, EHRENREICH I M, BROWN S L, et al. Phenotypic and genotypic characterization of multiple strains of the diazotrophic cyanobacterium,, isolated from the open ocean[J]. Environmental Microbiology, 2009, 11(2): 338–348.
[46] BERNSTEIN H C, CHARANIA M A, MCCLURE R S, et al. Multi-omic dynamics associate oxygenic photosynthesis with nitrogenase-mediated H2production insp. ATCC 51142[J]. Scientific Reports, 2015, 5: 16004.
[47] BOSTR?M K H, RIEMANN L, KüHL M, et al. Isolation and gene quantification of heterotrophic N2-fixing bacterioplankton in the Baltic Sea[J]. Environmental Microbiology, 2007, 9(1): 152–164.
[48] MARTINEZ-GARCIA S, FERNANDEZ E, CALVO-DIAZ A, et al. Response of heterotrophic and autotrophic microbial plankton to inorganic and organic inputs along a latitudinal transect in the Atlantic Ocean[J]. Biogeosciences, 2010, 7(5): 1701–1713.
[49] BIRD C, WYMAN M. Transcriptionally active hetero-trophic diazotrophs are widespread in the upper water column of the Arabian Sea[J]. FEMS Microbiology Ecology, 2013, 84(1): 189–200.
[50] FARNELID H, HARDER J, BENTZON-TILIA M, et al. Isolation of heterotrophic diazotrophic bacteria from estuarine surface waters[J]. Environmental Microbiology, 2014, 16(10): 3072–3082.
[51] KUMAR P K, SINGH A, RAMESH R, et al. N2fixation in the Eastern Arabian Sea: Probable role of heterotrophic diazotrophs[J]. Frontiers in Marine Science, 2017, 4: 80.
[52] WU C, KAN J, LIU H, et al. Heterotrophic bacteria dominate the diazotrophic community in the eastern Indian Ocean (EIO) during pre-southwest monsoon[J]. Microbial Ecology, 2019, 78(2019): 804–819.
[53] CHEN T-Y, CHEN Y-L L, SHEU D-S, et al. Community and abundance of heterotrophic diazotrophs in the northern South China Sea: Revealing the potential importance of a new alphaproteobacterium in N2fixation[J]. Deep-sea Research Part I-oceanographic Research Papers, 2019, 143(2019): 104–114.
[54] STAAL M, MEYSMAN F J, STAL L J. Temperature excludes N2-fixing heterocystous cyanobacteria in the tropical oceans[J]. Nature, 2003, 425(6957): 504–507.
[55] RIEMANN L, FARNELID H, STEWARD G F. Nitro-genase genes in non-cyanobacterial plankton: prevalence, diversity and regulation in marine waters[J]. Aquatic Microbial Ecology, 2010, 61(3): 225–237.
[56] CARPENTER E. Physiology and ecology of marine plank-tonic Oscillatoria ()[J]. Marine Biology Letters, 1983, 4(2): 69–85.
[57] MCCARTHY J J, CARPENTER E J. Oscillatoriacyanophyta in the central north atlantic ocean[J]. Journal of Phycology, 1979, 15(1): 75–82.
[58] BREITBARTH E, OSCHLIES A, LAROCHE J. Physiolo-gical constraints on the global distribution of- effect of temperature on diazotrophy[J]. Biogeosciences (BG), 2007, 4(1): 53–61.
[59] BRAUER V S, STOMP M, ROSSO C, et al. Low temperature delays timing and enhances the cost of nitrogen fixation in the unicellular cyanobacterium[J]. Isme Journal, 2013, 7(11): 2105–2115.
[60] FU F X, BELL P R F. Factors affecting N2fixation by the cyanobacteriumsp GBR-TRLI101[J]. Fems Microbiology Ecology, 2003, 45(2): 203–209.
[61] GROSSE J, BOMBAR D, HAI N D, et al. The Mekong River plume fuels nitrogen fixation and determines phytoplankton species distribution in the South China Sea during low- and high-discharge season[J]. Limnology and Oceanography, 2010, 55(4): 1668–1680.
[62] GALLON J. N2fixation in phototrophs: adaptation to a specialized way of life[J]. Plant and Soil, 2001, 230(1): 39–48.
[63] SA?UDO-WILHELMY S A, KUSTKA A B, GOBLER C J, et al. Phosphorus limitation of nitrogen fixation byin the central Atlantic Ocean[J]. Nature, 2001, 411(6833): 66–69.
[64] LU Y, WEN Z, SHI D, et al. Effect of light on N2fixation and net nitrogen release ofin a field study[J]. Biogeosciences, 2018, 15(1): 1–12.
[65] HO T-Y, CHU T-H, HU C-L J F I M. Interrelated influence of light and Ni ongrowth[J]. Fiontiers in Microbiology, 2013, 4: Article 139.
[66] GARCIA N S, FU F X, BREENE C L, et al. Combined effects of CO2and light on large and small isolates of the unicellular N2-fixing cyanobacteriumfrom the western tropical Atlantic Ocean[J]. European Journal of Phycology, 2013, 48(1): 128–139.
[67] RABOUILLE S, VAN DE WAAL D B, MATTHIJS H C P, et al. Nitrogen fixation and respiratory electron transport in the cyanobacteriumunder different light/dark cycles[J]. Fems Microbiology Ecology, 2014, 87(3): 630– 638.
[68] CAI X N, HUTCHINS D A, FU F X, et al. Effects of ultraviolet radiation on photosynthetic performance and N2fixation inIMS 101[J]. Biogeosciences, 2017, 14(19): 4455–4466.
[69] GALLON J R, HAMADI A F. Studies on the effects of oxygen on acetylene reduction (nitrogen fixation) insp. ATCC 27152[J]. Microbiology, 1984, 130(3): 495–503.
[70] GALLON J. Reconciling the incompatible: N2fixation and O2[J]. New Phytologist, 1992, 122(4): 571–609.
[71] RAVEN J A. The iron and molybdenum use efficiencies of plant growth with different energy, carbon and nitrogen sources[J]. New Phytologist, 1988, 109(3): 279–287.
[72] JICKELLS T D, AN Z S, ANDERSEN K K, et al. Global iron connections between desert dust, ocean biogeochemistry, and climate[J]. Science, 2005, 308(5718): 67–71.
[73] MOORE C M, MILLS M M, ACHTERBERG E P, et al. Large-scale distribution of Atlantic nitrogen fixation controlled by iron availability[J]. Nature Geoscience, 2009, 2(12): 867–871.
[74] WU J, CHUNG S W, WEN L S, et al. Dissolved inorganic phosphorus, dissolved iron, andin the oligotrophic South China Sea[J]. Global Biogeochemical Cycles, 2003, 17(1): 8-1-8-10.
[75] CHAPPELL P D, MOFFETT J W, HYNES A M, et al. Molecular evidence of iron limitation and availability in the global diazotroph[J]. Isme Journal, 2012, 6(9): 1728–1739.
[76] TURK-KUBO K A, ACHILLES K M, SERROS T R C, et al. Nitrogenase () gene expression in diazotrophic cyanobacteria in the Tropical North Atlantic in response to nutrient amendments[J]. Frontiers in Microbiology, 2012, 3: Article 386.
[77] BERMAN-FRANK I, CULLEN J T, SHAKED Y, et al. Iron availability, cellular iron quotas, and nitrogen fixation in[J]. Limnology and Oceanography, 2001, 46(6): 1249–1260.
[78] WHITTAKER S, BIDLE K D, KUSTKA A B, et al. Quantification of nitrogenase inIMS 101: implications for iron limitation of nitrogen fixation in the ocean[J]. Environmental Microbiology Reports, 2011, 3(1): 54–58.
[79] KUEPPER H, SETLIK I, SEIBERT S, et al. Iron limitation in the marine cyanobacteriumreveals new insights into regulation of photosynthesis and nitrogen fixation[J]. New Phytologist, 2008, 179(3): 784–798.
[80] WEBB E A, MOFFETT J W, WATERBURY J B. Iron Stress in open-ocean cyanobacteria (,, andspp.): Identification of the IdiA protein[J]. Applied and Environmental Micro-biology, 2001, 67(12): 5444–5452.
[81] JACQ V, RIDAME C, L'HELGUEN S, et al. Response of the unicellular diazotrophic cyanobacteriumto iron limitation[J]. PloS One, 2014, 9(1): e86749.
[82] WU J F, SUNDA W, BOYLE E A, et al. Phosphate depletion in the western North Atlantic Ocean[J]. Science, 2000, 289(5480): 759–762.
[83] HYNES A M, CHAPPELL P D, DYHRMAN S T, et al. Cross-basin comparison of phosphorus stress and nitrogen fixation in[J]. Limnology and Oceano-graphy, 2009, 54(5): 1438–1448.
[84] WEBB E A, JAKUBA R W, MOFFETT J W, et al. Molecular assessment of phosphorus and iron physiology inpopulations from the western Central and western South Atlantic[J]. Limnology and Oceanography, 2007, 52(5): 2221–2232.
[85] ZHANG R, CHEN M, YANG Q, et al. Physical-biological coupling of N2fixation in the northwestern South China Sea coastal upwelling during summer[J]. Limnology and Oceanography, 2015, 60(4): 1411–1425.
[86] MOISANDER P H, STEPPE T F, HALL N S, et al. Variability in nitrogen and phosphorus limitation for Baltic Sea phytoplankton during nitrogen-fixing cyanobacterial blooms[J]. Marine Ecology Progress Series, 2003, 262: 81–95.
[87] VILLAREAL T, CARPENTER E. Buoyancy regulation and the potential for vertical migration in the oceanic cyanobacterium[J]. Microbial Ecology, 2003, 45(1): 1–10.
[88] DYHRMAN S T, CHAPPELL P D, HALEY S T, et al. Phosphonate utilization by the globally important marine diazotroph[J]. Nature, 2006, 439(7072): 68–71.
[89] DYHRMAN S T, HALEY S T. Phosphorus scavenging in the unicellular marine diazotroph[J]. Applied and Environmental Microbiology, 2006, 72(2): 1452–1458.
[90] PEREIRA N, SHILOVA I N, ZEHR J P. Use of the high-affinity phosphate transporter gene,, as an indicator for phosphorus stress in the marine diazotroph(Chroococcales, Cyanobacteria)[J]. Journal of Phycology, 2019, 55(2019): 752–761.
[91] MILLS M M, RIDAME C, DAVEY M, et al. Iron and phosphorus co-limit nitrogen fixation in the eastern tropical North Atlantic[J]. Nature, 2004, 429(6989): 292–294.
[92] CURATTI L, HERNANDEZ J A, IGARASHI R Y, et al. In vitro synthesis of the iron molybdenum cofactor of nitrogenase from iron, sulfur, molybdenum, and homocitrate using purified proteins[J]. Proceedings of the National Academy of Sciences, 2007, 104(45): 17626– 17631.
[93] MULHOLLAND M R, CAPONE D G. Nitrogen fixation, uptake and metabolism in natural and cultured populations ofspp.[J]. Marine Ecology Progress Series, 1999, 188: 33–49.
[94] KNAPP A N, DEKAEZEMACKER J, BONNET S, et al. Sensitivity ofandabundance and N2fixation rates to varying NO3-and PO43-concentrations in batch cultures[J]. Aquatic Microbial Ecology, 2012, 66(3): 223–236.
[95] WARD B A, DUTKIEWICZ S, MOORE C M, et al. Iron, phosphorus, and nitrogen supply ratios define the biogeography of nitrogen fixation[J]. Limnology and Oceanography, 2013, 58(6): 2059–2075.
[96] WEN Z Z, LIN W F, SHEN R, et al. Nitrogen fixation in two coastal upwelling regions of the Taiwan Strait[J]. Scientific Reports, 2017, 7: 17601.
[97] MCGILLICUDDY D. Mechanisms of physical–biological– biogeochemical interactions at the oceanic mesoscale[J]. Annual Review of Marine Science, 2016, 8: 13.11–13.36.
[98] MCGILLICUDDY D J, ROBINSON A R, SIEGEL D A, et al. Influence of mesoscale eddies on new production in the Sargasso Sea[J]. Nature, 1998, 394(6690): 263–266.
[99] BENITEZ-NELSON C R, BIDIGARE R R, DICKEY T D, et al. Mesoscale eddies drive increased silica export in the subtropical Pacific Ocean[J]. Science, 2007, 316(5827): 1017–1021.
[100]CARPENTER E J, MCCARTHY J J. Nitrogen fixation and uptake of combined nitrogenous nutrients by Oscillatoriain the western Sargasso Sea[J]. Limnology and Oceanography, 1975, 20(3): 389–401.
[101]CHEN Y-L L, CHEN H-Y, TUO S-H, et al. Seasonal dynamics of new production fromN2fixation and nitrate uptake in the upstream Kuroshio and South China Sea basin[J]. Limnology and Oceanography, 2008, 53(5): 1705.
[102]DAVIS C S, MCGILLICUDDY D J, JR. Transatlantic abundance of the N2-fixing colonial cyanobacterium[J]. Science, 2006, 312(5779): 1517–1520.
[103]OLSON E M, MCGILLICUDDY D J, FLIERL G R, et al. Mesoscale eddies andspp. distributions in the southwestern North Atlantic[J]. Journal of Geophysical Research: Oceans, 2015, 120(6): 4129–4150
[104]FONG A A, KARL D M, LUKAS R, et al. Nitrogen fixation in an anticyclonic eddy in the oligotrophic North Pacific Ocean[J]. Isme Journal, 2008, 2(6): 663–676.
[105]CHURCH M J, MAHAFFEY C, LETELIER R M, et al. Physical forcing of nitrogen fixation and diazotroph community structure in the North Pacific subtropical gyre[J]. Global Biogeochemical Cycles, 2009, 23(2): GB2020.
[106]HOLL C M, WAITE A M, PESANT S, et al. Unicellular diazotrophy as a source of nitrogen to Leeuwin Current coastal eddies[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2007, 54(8): 1045–1054.
[107]RAHAV E, HERUT B, STAMBLER N, et al. Uncoupling between dinitrogen fixation and primary productivity in the eastern Mediterranean Sea[J]. Journal of Geophysical Research-Biogeosciences, 2013, 118(1): 195–202.
[108]LOSCHER C R, BANGE H W, SCHMITZ R A, et al. Water column biogeochemistry of oxygen minimum zones in the eastern tropical North Atlantic and eastern tropical South Pacific oceans[J]. Biogeosciences, 2016, 13(12): 3585–3606.
[109]LIU J, ZHOU L, LI J, et al. Effect of mesoscale eddies on diazotroph community structure and nitrogen fixation rates in the South China Sea[J]. Regional Studies in Marine Science, 2020, 35: 101106.
[110]ZHANG Y, ZHAO Z, SUN J, et al. Diversity and distribution of diazotrophic communities in the South China Sea deep basin with mesoscale cyclonic eddy perturbations[J]. Fems Microbiology Ecology, 2011, 78(3): 417–427.
[111]GONZALEZ TABOADA F, GONZALEZ GIL R, HOEFER J, et al.spp. population structure in the eastern North Atlantic subtropical gyre[J]. Deep-sea Research Part I-oceanographic Research Papers, 2010, 57(1): 65–77.
[112]LOSCHER C R, BOURBONNAIS A, DEKAEZEMACKER J, et al. N2fixation in eddies of the eastern tropical South Pacific Ocean[J]. Biogeosciences, 2016, 13(10): 2889–2899.
[113]SPILLING K, CAMARENA-GOMEZ M-T, LIPSEWERS T, et al. Impacts of reduced inorganic N:P ratio on three distinct plankton communities in the Humboldt upwelling system[J]. Marine Biology, 2019, 166:114.
[114]CAPONE D G, HUTCHINS D A. Microbial biogeo-chemistry of coastal upwelling regimes in a changing ocean[J]. Nature Geoscience, 2013, 6(9): 711–717.
[115]KNAPP A N. The sensitivity of marine N2fixation to dissolved inorganic nitrogen[J]. Frontiers in Microbiology, 2012, 3: Article 374.
[116]SUBRAMANIAM A, MAHAFFEY C, JOHNS W, et al. Equatorial upwelling enhances nitrogen fixation in the Atlantic Ocean[J]. Geophysical Research Letters, 2013, 40(9): 1766–1771.
[117]TOVAR-SANCHEZ A, SANUDO-WILHELMY S A. Influence of the Amazon River on dissolved and intra-cellular metal concentrations incolonies along the western boundary of the sub-tropical North Atlantic Ocean[J]. Biogeosciences, 2011, 8(1): 217–225.
[118]FERREIRA P A A, BOMFETI C A, SOARES B L, et al. Efficient nitrogen-fixing Rhizobium strains isolated from amazonian soils are highly tolerant to acidity and aluminium[J]. World Journal of Microbiology & Biotechnology, 2012, 28(5): 1947–1959.
[119]HILTON J A, SATINSKY B M, DOHERTY M, et al. Metatranscriptomics of N2-fixing cyanobacteria in the Amazon River plume[J]. Isme Journal, 2015, 9(7): 1557–1569.
[120]WASMUND N, STRUCK U, HANSEN A, et al. Missing nitrogen fixation in the Benguela region[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2015, 106(2015): 30–41.
[121]FERRANDO L, SCAVINO A F. Strong shift in the diazotrophic endophytic bacterial community inhabiting rice () plants after flooding[J]. Fems Microbiology Ecology, 2015, 91(9): fv104.
[122]FERNANDEZ C, FARíAS L, ULLOA O. Nitrogen fixation in denitrified marine waters[J]. PloS One, 2011, 6(6): e20539.
[123]FERNANDEZ C, GONZALEZ M L, MUNOZ C, et al. Temporal and spatial variability of biological nitrogen fixation off the upwelling system of central Chile (35-38.5 degrees S)[J]. Journal of Geophysical Research-Oceans, 2015, 120(5): 3330–3349.
[124]GROSSKOPF T, MOHR W, BAUSTIAN T, et al. Doubling of marine dinitrogen-fixation rates based on direct measurements[J]. Nature, 2012, 488(7411): 361–364.
[125]WILSON S T, BOTTJER D, CHURCH M J, et al. Comparative assessment of nitrogen fixation methodologies, conducted in the oligotrophic north pacific ocean[J]. Applied and Environmental Microbiology, 2012, 78(18): 6516–6523.
[126]呂娥. 中國邊緣海海洋固氮作用的研究—乙炔還原法的應用[D]. 廈門: 廈門大學, 2007.
[127]MAGUE T H, WEARE N M, HOLM-HANSEN O. Nitrogen fixation in the North Pacific Ocean[J]. Marine Biology, 1974, 24(2): 109–119.
[128]SROGA G E, LANDEGREN U, BERGMAN B, et al. Isolation ofand part ofby modified capture polymerase chain reaction from a natural population of the marine cyanobacteriumsp.[J]. Fems Microbiology Letters, 1996, 136(2): 137–145.
[129]ZEHR J P, BRAUN S, CHEN Y B, et al. Nitrogen fixation in the marine environment: Relating genetic potential to nitrogenase activity[J]. Journal of Experimental Marine Biology and Ecology, 1996, 203(1): 61–73.
[130]ZEHR J P, MONTOYA J P, JENKINS B D, et al. Experiments linking nitrogenase gene expression to nitrogen fixation in the North Pacific subtropical gyre[J]. Limnology and Oceanography, 2007, 52(1): 169–183.
[131]陳晨, 柏耀輝, 梁金松, 等. FISH-NanoSIMS技術在環境微生物生態學上的應用研究[J]. 環境科學, 2015, (01): 244–251.
[132]CHEN Y F L. Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea[J]. Deep-sea Research Part I-oceanographic Research Papers, 2005, 52(2): 319–340.
[133]KONG L, JING H, KATAOKA T, et al. Phylogenetic diversity and spatio-temporal distribution of nitrogenase genes () in the northern South China Sea[J]. Aquatic Microbial Ecology, 2011, 65(1): 15–27.
[134]DONG J, ZHANG Y, WANG Y, et al. Spatial and seasonal variations of cyanobacteria and their nitrogen fixation rates in Sanya Bay, South China Sea[J]. Scientia Marina, 2008, 72(2): 239–251.
[135]李濤, 劉勝, 黃良民, 等. 大亞灣紅海束毛藻赤潮生消過程研究[J]. 海洋環境科學, 2008, 27(03): 224–227.
[136]WEN Z, LIN W, SHEN R, et al. Nitrogen fixation in two coastal upwelling regions of the Taiwan Strait[J]. Scientific Reports, 2017, 7(1): 17601.
[137]MOISANDER P H, BEINART R A, VOSS M, et al. Diversity and abundance of diazotrophic microorganisms in the South China Sea during intermonsoon[J]. The ISME Journal, 2008, 2(9): 954–967.
[138]BOMBAR D, MOISANDER P H, DIPPNER J W, et al. Distribution of diazotrophic microorganisms andgene expression in the Mekong River plume during intermon-soon[J]. Marine Ecology Progress Series, 2011, 424: 39–52.
[139]CHEN Y-L L, CHEN H-Y, LIN Y-H. Distribution and downward flux ofin the South China Sea as influenced by the transport from the Kuroshio Current[J]. Marine Ecology Progress Series, 2003, 259: 47–57.
[140]CHEN Y L L, CHEN H Y, KARL D M, et al. Nitrogen modulates phytoplankton growth in spring in the South China Sea[J]. Continental Shelf Research, 2004, 24(4-5): 527–541.
[141]SHIOZAKI T, CHEN Y-L L, LIN Y-H, et al. Seasonal variations of unicellular diazotroph groups A and B, andin the northern South China Sea and neighboring upstream Kuroshio Current[J]. Continental Shelf Research, 2014, 80(2014): 20–31.
[142]SHEN P P, TAN Y H, HUANG L M, et al. Occurrence of brackish water phytoplankton species at a closed coral reef in Nansha Islands, South China Sea[J]. Marine Pollution Bulletin, 2010, 60(10): 1718–1725.
[143]劉甲星, 周林濱, 李剛, 等. 秋季南海東北部表層水體固氮及其對初級生產力貢獻[J]. 熱帶海洋學報, 2016, 35(5): 38–47.
[144]CHOU W C, CHEN Y L L, SHEU D D, et al. Estimated net community production during the summertime at the SEATS time-series study site, northern South China Sea: Implications for nitrogen fixation[J]. Geophysical Research Letters, 2006, 33(22): L22610.
[145]CHEN Y-L L, CHEN H-Y, LIN Y-H, et al. The relative contributions of unicellular and filamentous diazotrophs to N2fixation in the South China Sea and the upstream Kuroshio[J]. Deep-sea Research Part I-oceanographic Research Papers, 2014, 85(2014): 56–71.
[146]DANG H, YANG J, LI J, et al. Environment-Dependent Distribution of the Sediment-Harboring Microbiota in the Northern South China Sea[J]. Applied and Environmental Microbiology, 2013, 79(1): 121–132.
[147]YIN J-P, LING J, DONG J-D, et al. Vertical variations of bacterial community composition in south china sea determined by DGGE fingerprinting and multivariate analysis[J]. Polish Journal of Environmental Studies, 2014, 23(5): 1787–1796.
[148]XIAO P, JIANG Y, LIU Y, et al. Re-evaluation of the diversity and distribution of diazotrophs in the South China Sea by pyrosequencing thegene[J]. Marine and Freshwater Research, 2015, 66(8): 681–691.
[149]LI D, LIU J, ZHANG R, et al. N2fixation impacted by carbon fixation via dissolved organic carbon in the changing Daya Bay, South China Sea[J]. Science of the Total Environment, 2019, 674(2019): 592–602.
[150]林峰, 陳敏, 楊偉鋒, 等. 夏季臺灣海峽南部海域上層水體的生物固氮作用[J]. 生態學報, 2013, 33(19): 6354– 6362.
[151]李麗. 廣東沿海典型赤潮高發區海洋環境生態研究[D]. 廣州: 暨南大學, 2009.
[152]石榮貴, 連忠廉, 龍愛民, 等. 北部灣冬季的生物固氮作用[J]. 海洋環境科學, 2012, 31(004): 524–528.
[153]ZHANG R, ZHU X, YANG C, et al. Distribution of dissolved iron in the Pearl River (Zhujiang) Estuary and the northern continental slope of the South China Sea[J]. Deep Sea Research Part II: Topical Studies in Oceano-graphy, 2018, 167(2019): 14–24.
[154]LOICK N, DIPPNER J, DOAN H N, et al. Pelagic nitrogen dynamics in the Vietnamese upwelling area according to stable nitrogen and carbon isotope data[J]. Deep-sea Research Part I-oceanographic Research Papers, 2007, 54(4): 596–607.
[155]BLAIS M, TREMBLAY J é, JUNGBLUT A D, et al. Nitrogen fixation and identification of potential diazotrophs in the Canadian Arctic[J]. Global Biogeoche-mical Cycles, 2012, 26(3): GB3022.
[156]DíEZ B, BERGMAN B, PEDRóS-ALIó C, et al. High cyanobacterialgene diversity in Arctic seawater and sea ice brine[J]. Environmental Microbiology Reports, 2012, 4(3): 360–366.
[157]RAHAV E, HERUT B, MULHOLLAND M R, et al. Heterotrophic and autotrophic contribution to dinitrogen fixation in the Gulf of Aqaba[J]. Marine Ecology Progress Series, 2015, 522: 67–77.
[158]ZEHR J P. Nitrogen fixation by marine cyanobacteria[J]. Trends in Microbiology, 2011, 19(4): 162–173.
[159]LE MOAL M, BIEGALA I C. Diazotrophic unicellular cyanobacteria in the northwestern Mediterranean Sea: a seasonal cycle[J]. Limnology and Oceanography, 2009, 54(3): 845–855.
[160]BOSTROM K H, RIEMANN L, KUHL M, et al. Isolation and gene quantification of heterotrophic N2-fixing bacterioplankton in the Baltic Sea[J]. Environmental Microbiology, 2007, 9(1): 152–164.
[161]RAHAV E, GIANNETTO M J, BAR-ZEEV E. Contri-bution of mono and polysaccharides to heterotrophic N2fixation at the eastern Mediterranean coastline[J]. Scientific Reports, 2016, 6: 27858.
[162]DEUTSCH C, SARMIENTO J L, SIGMAN D M, et al. Spatial coupling of nitrogen inputs and losses in the ocean[J]. Nature, 2007, 445(7124): 163–167.
[163]CODISPOTI L. An oceanic fixed nitrogen sink exceeding 400 Tg N a-1vs the concept of homeostasis in the fixed- nitrogen inventory[J]. Biogeosciences, 2007, 4(2): 233–253.
[164]GRUBER N, SARMIENTO J L. Global patterns of marine nitrogen fixation and denitrification[J]. Global Biogeochemical Cycles, 1997, 11(2): 235–266.
[165]ALTABET M. Constraints on oceanic N balance/ imbalance from sedimentary15N records[J]. Biogeos-ciences, 2006, 3(4): 1121–1155.
[166]MAHAFFEY C, MICHAELS A F, CAPONE D G. The conundrum of marine N2fixation[J]. American Journal of Science, 2005, 305(6-8): 546–595.
Research progress in marine biological nitrogen fixation
LI Zhihong1, LI Jinyou2, LIU Jiaxing3, 4, *
1. Development Promotion Center of Marine Fishery Science and Technology, Ocean Development Experimental Zone of Wanshan, Zhuhai 519005, China 2. School of Data Science, City University of Hong Kong, Hong Kong, China 3. Key Laboratory of Tropical Marine Bio-resources and Ecology, Guangdong Provincial Key Laboratory of Applied Marine Biology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China 4. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China
Marine nitrogen fixation plays an important role in the global carbon and nitrogen cycles because it can support the nitrogen required for primary production. The importance of unicellular nitrogen-fixing cyanobacteria and heterotrophic nitrogen-fixing bacteria has gradually been known since the application of molecular biology and15N2isotope tracing in nitrogen fixation study in the 1990s. Firstly, it is suggested that the nitrogen fixation amount estimated based onmay beunderestimated. Secondly, traditional study of marine biological nitrogen fixation is limited to the tropical and subtropical oligotrophic regions, while less attention has been paid to high nutrient regions such as upwelling and estuaries. Therefore, it is necessary to re-evaluate and re-understand the nitrogen fixation in these regions. This article reviewed the recent progress of nitrogen fixation, including biodiversity and distribution of diazotrophs, limiting factors, methods, and existing question. In addition, the latest progress and question of nitrogen fixation in the South China Sea were also reviewed.
marine biological nitrogen fixation; unicellular nitrogen-fixing cyanobacteria; heterotrophic nitrogen-fixing bacteria; molecular biology technology; high nutrient regions; South China Sea
10.14108/j.cnki.1008-8873.2021.05.026
P735
A
1008-8873(2021)05-215-16
2020-02-26;
2020-03-29
科技基礎資源調查專項(2018FY100105); 南方海洋科學與工程廣東省實驗室(廣州)人才團隊引進重大專項(GML2019ZD0401); 廣東省促進經濟高質量發展專項資金海洋經濟發展項目(GDOE[2019]A32); 國家自然科學基金(41806198, 31971432); 廣東省基礎與應用基礎研究基金(2019A1515010896); 廣州市科技計劃項目(202102020279)
李志紅 (1970—), 女, 湖南雙峰縣人, 碩士, 從事海洋漁業及生態學方面工作, E-mail: 799491187@qq.com
通信作者:劉甲星, 男, 博士, 助理研究員, 研究方向為海洋生態學, E-mail: ljx2ljx@sisio.ac.cn
李志紅, 李勁尤, 劉甲星. 海洋生物固氮研究進展[J]. 生態科學, 2021, 40(5): 215–230.
LI Zhihong, LI Jinyou, LIU Jiaxing. Research progress in marine biological nitrogen fixation[J]. Ecological Science, 2021, 40(5): 215–230.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
