內(nèi)蒙古荒漠灌木根際解磷菌多樣性及其解磷和產(chǎn)鐵載體能力
2021-11-22 08:33:58孟建宇楊鴻儒賈麗娟
環(huán)境科學研究 2021年11期
關(guān)鍵詞:能力
孟建宇, 李 蘅*, 楊鴻儒, 賈麗娟,2
1.內(nèi)蒙古農(nóng)業(yè)大學生命科學學院, 內(nèi)蒙古 呼和浩特 010011
2.中國農(nóng)業(yè)科學院草原研究所, 內(nèi)蒙古 呼和浩特 010010
磷是植物體中蛋白質(zhì)、核酸和ATP的重要組分,參與多種理化反應的調(diào)節(jié)過程,是其生長發(fā)育過程中必需的營養(yǎng)元素[1]. 土壤中95%以上的磷是不溶性的磷酸鹽形式,不具生物有效性[2-3]. 為解決土壤含磷量不足、補充植物生長所需,在實際生產(chǎn)過程需施用大量磷肥,但這會引起食物安全、土壤環(huán)境污染等問題[4-5].
解磷細菌(phosphate-solubilizing bacteria,PSB)是土壤中可將不溶性磷素轉(zhuǎn)化為生物能直接利用的磷的一類功能微生物類群,通過分泌多種有機酸、酶類來活化不溶性磷,提高有效的生物可利用磷含量,促進植物的生長和發(fā)育[6]. 研究發(fā)現(xiàn),接種PSB后能顯著增加土壤及植株的磷、鉀、鎂等礦質(zhì)元素含量[7-9],而且PSB還可改善根際微生物群落結(jié)構(gòu),促進土壤叢枝菌根對植物根系的侵染,提高植物生長的品質(zhì)[10]. 畢銀麗等[11]將分離得到的PGPR(plant growth promoting rhizobacteria)制成的菌肥施加到荒漠植物的土壤中,不僅提高了有效磷含量,土質(zhì)由酸性變?yōu)橹行裕⑶乙苍鰪娏怂嵝粤姿崦富钚裕罱K加快了植物的生長發(fā)育. 邢芳芳等[12]分離得到的解磷菌枯草芽孢桿菌(Bacillussubtilis)對白菜有明顯的促生作用,經(jīng)其處理后,白菜葉片數(shù)、葉綠素以及產(chǎn)量都有明顯提高. 不同類型土壤和植物根際土壤中廣泛分布有PSB,其種群、功能和遺傳特性等豐富多樣[13-14]. 目前對土壤PSB的研究主要集中于玉米、番茄、小麥和油菜等農(nóng)作物[15],但對荒漠根際PSB的報道還較少.
內(nèi)蒙古荒漠區(qū)分布廣泛,其面積約占全區(qū)總面積的51.50%,植被主要有霸王(Zygophyllumxanthonylon)、四合木(Tetraenatnongolica)、沙冬青(Ammopiptanthusmongolicus)、半日花(Helianthemumsoongoricum)和白刺(Nitrariatangutorum)等強旱生、古老孑遺灌木. 它們在保持土壤穩(wěn)定、改善生態(tài)環(huán)境方面起著非常重要的作用,而能產(chǎn)鐵載體的PSB對灌木生長又有著積極的促進作用. 因此,分析內(nèi)蒙古荒漠灌木根際PSB類群,認識荒漠灌木植物PSB多樣性及其促生特性,豐富功能根際促生細菌資源庫以及開發(fā)新的改善土壤磷素營養(yǎng)途徑,對于改良和保護荒漠地區(qū)的生態(tài)環(huán)境具有積極而重大的意義.
1 材料與方法
1.1 試驗材料
1.1.1樣品采集
樣品為內(nèi)蒙古西鄂爾多斯荒漠的霸王、半日花、四合木、白刺和沙冬青5種灌木. 采樣方法:先用工具鏟除去落葉層,再用鐵鏟挖去植物基部的上層覆土,剪下一部分主根和須根,用刷子刮下粘在根段上的土壤,裝入無菌聚乙烯袋.
1.1.2培養(yǎng)基
牛肉膏蛋白胨培養(yǎng)基:牛肉膏3.0 g/L,蛋白胨10.0 g/L,氯化鈉5.0 g/L.
LB培養(yǎng)基:胰蛋白胨10.0 g/L, 酵母粉5.0 g/L,氯化鈉5.0 g/L,pH為7.2.
無機磷培養(yǎng)基:磷酸鈣10.0 g/L,葡萄糖10.0 g/L,硫酸鎂0.3 g/L,氯化鉀0.3 g/L,硫酸銨0.5 g/L,氯化鈉0.3 g/L,硫酸亞鐵0.03 g/L,硫酸錳0.03 g/L,pH為7.0~7.5.
PVK培養(yǎng)基:葡萄糖10.0 g/L,磷酸鈣5.0 g/L,硫酸銨0.5 g/L,氯化鈉0.2 g/L,硫酸鎂0.1 g/L,氯化鉀0.2 g/L,酵母粉0.5 g/L,硫酸錳0.002 g/L,硫酸亞鐵0.002 g/L,0.4%溴酚藍0.6 g/L,瓊脂18.0 g/L,pH為7.0~7.2.
有機磷培養(yǎng)基:葡萄糖10.0 g/L,硫酸銨0.2 g/L,氯化鎂5.0 g/L,硫酸鎂0.5 g/L,氯化鉀0.1 g/L,植酸鈣2.0 g/L.
液體發(fā)酵培養(yǎng)基:蛋白胨3.0 g/L,葡萄糖15.0 g/L,可溶性淀粉20.0 g/L,硫酸銨0.5 g/L,氯化鉀0.5 g/L,硫酸鎂0.03 g/L,硫酸錳0.03 g/L,硫酸亞鐵0.04 g/L,磷酸二氫鉀0.02 g/L,pH為5.5.
MSA(蔗糖-天冬氨酸)培養(yǎng)基:蔗糖20.0 g/L,天冬酰胺2.0 g/L,磷酸氫二鉀1.0 g/L,硫酸鎂0.5 g/L,pH為7.0.
1.2 試驗方法
1.2.1根際解磷菌的分離
稱取植物根際土壤0.5 g放入49.5 mL無菌水的三角瓶中振蕩搖勻,按十倍稀釋法用無菌水稀釋成10-2~10-6梯度的菌懸液,分別涂布于無機磷培養(yǎng)基,每個梯度設(shè)3個平行,26 ℃下培養(yǎng). 5 d后挑取形態(tài)各異的單菌落進一步劃線純化. 將純化的菌株接種到PVK培養(yǎng)基上篩選解無機磷細菌,接種到有機磷培養(yǎng)基上篩選解有機磷細菌.
1.2.2菌株的16S rRNA鑒定及其同源性分析
用CTAB法提取菌株的DNA,16S rRNA基因擴增引物為27F和1492R. PCR擴增體系:5.0 μL 10×PCR Buffer,4.0 μL 2.5 mmol/L的dNTPs,0.5 μL 20 μmol/L的正反引物,0.3 μL 5 U/μL的Easy Taq DNA聚合酶,2.5 μL的DNA模板,37.7 μL ddH2O. 反應條件:94 ℃預變性3 min,94 ℃變性1 min,55 ℃退火1 min,72 ℃延伸1 min,30個循環(huán);72 ℃最終延伸5 min. 擴增產(chǎn)物用1.0%瓊脂糖凝膠電泳檢測后送至生工生物工程(上海)股份有限公司測序,測序的序列上傳NCBI,通過Blast-n與EX-TAXON數(shù)據(jù)庫中同源序列進行比對,然后用MEGA 6.0構(gòu)建系統(tǒng)發(fā)育樹.
1.2.3解無機磷能力的測定
將獲得的純培養(yǎng)物在PVK培養(yǎng)基上劃線培養(yǎng)3 d,挑出劃線部分呈藍色的菌株,采用鉬銻抗比色法測定其解無機磷能力. 將初篩菌株接種在LB培養(yǎng)基中,28 ℃下?lián)u床振蕩過夜. 離心獲得菌體后制成一定濃度的菌懸液,取0.5 mL接種于50 mL無機磷培養(yǎng)基中,置于30 ℃、170 r/min下培養(yǎng)7 d. 然后在 10 000 r/min下離心10 min,并測定上清液的pH. 取上清液1.25 mL于比色管中,再加入2.5 mL鉬銻抗比色劑,用去離子水定容,搖勻后靜置30 min,測定OD730 nm值,根據(jù)標準曲線y=2.733x+0.010 7(R2=0.997 5)計算含磷量.
1.2.4解有機磷能力的測定
將菌株接種到LB培養(yǎng)基中,28 ℃下?lián)u床振蕩過夜,離心獲得菌體后制成一定濃度的菌懸液,取0.5 mL接種到液體發(fā)酵培養(yǎng)基中,置于30 ℃、170 r/min下培養(yǎng)72 h. 發(fā)酵液于 10 000 r/min下離心10 min,取上清液1 mL,加入5 mmol/L的植酸鈉1 mL,37 ℃下保溫30 min,添加3 mL終止液中止反應,然后測定OD415 nm值. 參照磷標準曲線y=24.389x+0.036 6(R2=0.994 7)計算酶活性. 酶活性定義:在37 ℃、pH為5.5的條件下,從濃度為8.4 g/L的植酸鈉溶液中每分鐘釋放出1 μmol無機磷所需要的植酸酶量為1 IU/mL.
1.2.5產(chǎn)鐵載體能力分析
用滅菌的牙簽挑取單菌落,點接在CAS檢測平板上,28 ℃下恒溫培養(yǎng)96 h. 記錄橙色暈圈的有無,并對暈圈直徑(D)及菌落直徑(d)進行測定,計算二者比值(D/d),該值越大,表明產(chǎn)鐵載體能力越強,對篩選到的產(chǎn)鐵載體能力較強菌株參考Payne[16]的方法進行定量分析. 將可產(chǎn)生暈圈的菌株接種到MSA培養(yǎng)基中,置于30 ℃、15 r/min下培養(yǎng)48 h. 取2 mL 菌液于離心管中,6 000 r/min下離心10 min,取1.5 mL上清液加上1.5 mL CAS檢測液,充分混勻靜置1 h 后測定OD630 nm值(As),以雙蒸水作為對照. 另取1.5 mL CAS檢測液加1.5 mL未接菌的MSA培養(yǎng)基上清液混勻后測定OD630 nm值,作為參比值(Ar). 用As/Ar表示鐵載體的相對含量,數(shù)值越低,表示鐵載體的含量越大.As/Ar值在0~1.0之間以0.2為間隔,每減小0.2,增加一個“+”,As/Ar值低于0.5時產(chǎn)鐵載體能力較強(+++)[17].
2 結(jié)果與分析
2.1 根際解磷菌的分離及多樣性分析
從內(nèi)蒙古西鄂爾多斯荒漠5種灌木根際土壤中分離到62株細菌,其中半日花15株、白刺3株、霸王9株、四合木26株,沙冬青9株. 根際解磷菌分類類群(見表1)和系統(tǒng)發(fā)育分析(見圖1)表明,62株解磷細菌分屬于α-變形菌綱(α-proteobacteria,占比為22.58%)、β-變形菌綱(β-proteobacteria,占比為17.74%)、γ-變形菌綱(γ-proteobacteria,占比為8.07%)、芽孢桿菌綱(Bacilli,占比為17.74%)、放線菌綱(Actinobacteria,占比為32.26%)和噬纖維菌綱(Cytophagia,占比為1.61%). 其中,放線菌綱為最優(yōu)勢類群,α-變形菌綱是次優(yōu)勢類群. 在屬水平上,分屬于21個屬,分別為短波單胞菌屬(Brevundimonas,1株)、鞘氨醇單胞菌屬(Sphingomonas,5株)、蒼白桿菌屬(Ochrobactrum,3株)、中華根瘤菌屬(Sinorhizobium,4株)、根瘤菌屬(Rhizobium,1株)、馬賽菌屬(Massilia,9株)、Prolinoborus(1株)、產(chǎn)堿桿菌屬(Alcaligenes,1株)、單胞菌屬(Stenotrophomonas,2株)、假單胞菌屬(Psedumonas,3株)、芽孢桿菌屬(Bacillus,11株)、鏈霉菌屬(Streptomyces,9株)、紅球菌屬(Rhodococcus,3株)、分枝桿菌屬(Mycobacterium,1株)、冢村氏菌屬(Tsukamurella,1株)、諾卡氏菌屬(Nocardia,1株)、Nocardioide(2株)、短桿菌屬(Brevibacterium,1株)、細桿菌屬(Microbacterium,1株)、韓國生工屬(Kribbella,1株)和Dyadobacter(1株). 其中,芽孢桿菌屬為第一優(yōu)勢菌屬,占總菌數(shù)的17.74%,其次為馬賽菌屬和鏈霉菌屬,均分別占總菌數(shù)的14.52%. 在不同灌木中分離到的PSB多樣性差異較明顯,如半日花的優(yōu)勢菌屬為鞘氨醇單胞菌屬和馬賽菌屬(各占20.00%),霸王的優(yōu)勢菌屬是紅球菌屬和中華根瘤菌屬(各占22.22%),沙冬青的優(yōu)勢菌屬為馬賽菌屬和芽孢桿菌屬(各占22.22%),四合木的優(yōu)勢菌屬為芽孢桿菌屬和鏈霉菌屬(各占23.07%). 其中,芽孢桿菌屬和馬賽菌屬分布較廣,分別存在于4種灌木的根際土壤中;有些屬僅分布于一種灌木的根際土壤中,如短波單胞菌屬、韓國生工屬和Dyadobacter只在半日花根際土壤中分離到,產(chǎn)堿桿菌屬、根瘤菌屬、冢村氏菌屬和分枝桿菌屬只在四合木根際土壤中分離到,細桿菌屬和Prolinoborus只在霸王根際土壤中分離到,短桿菌屬和諾卡氏菌屬只在沙冬青根際土壤中分離到.
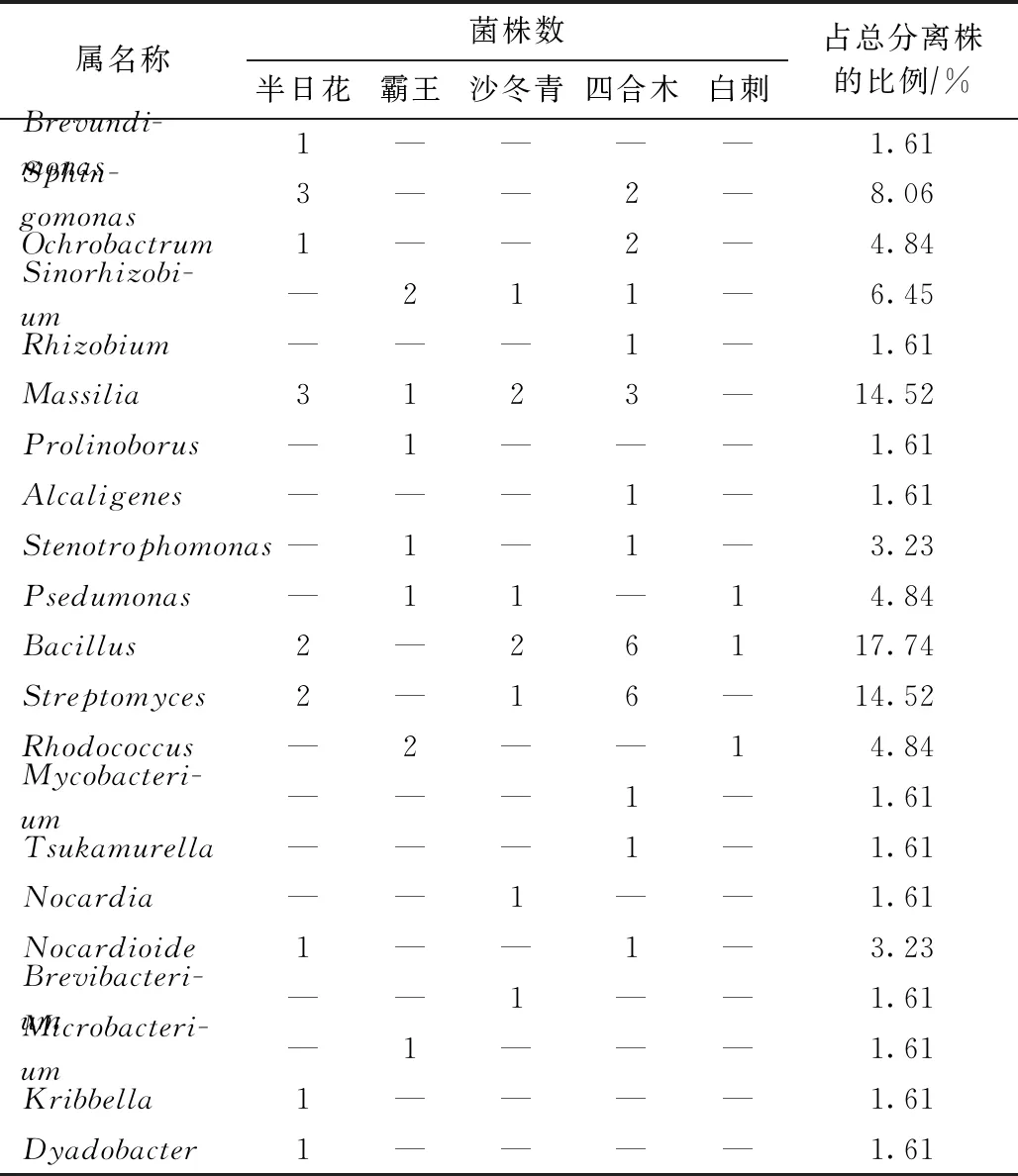
表1 分離所得根際解磷菌屬水平上的歸類
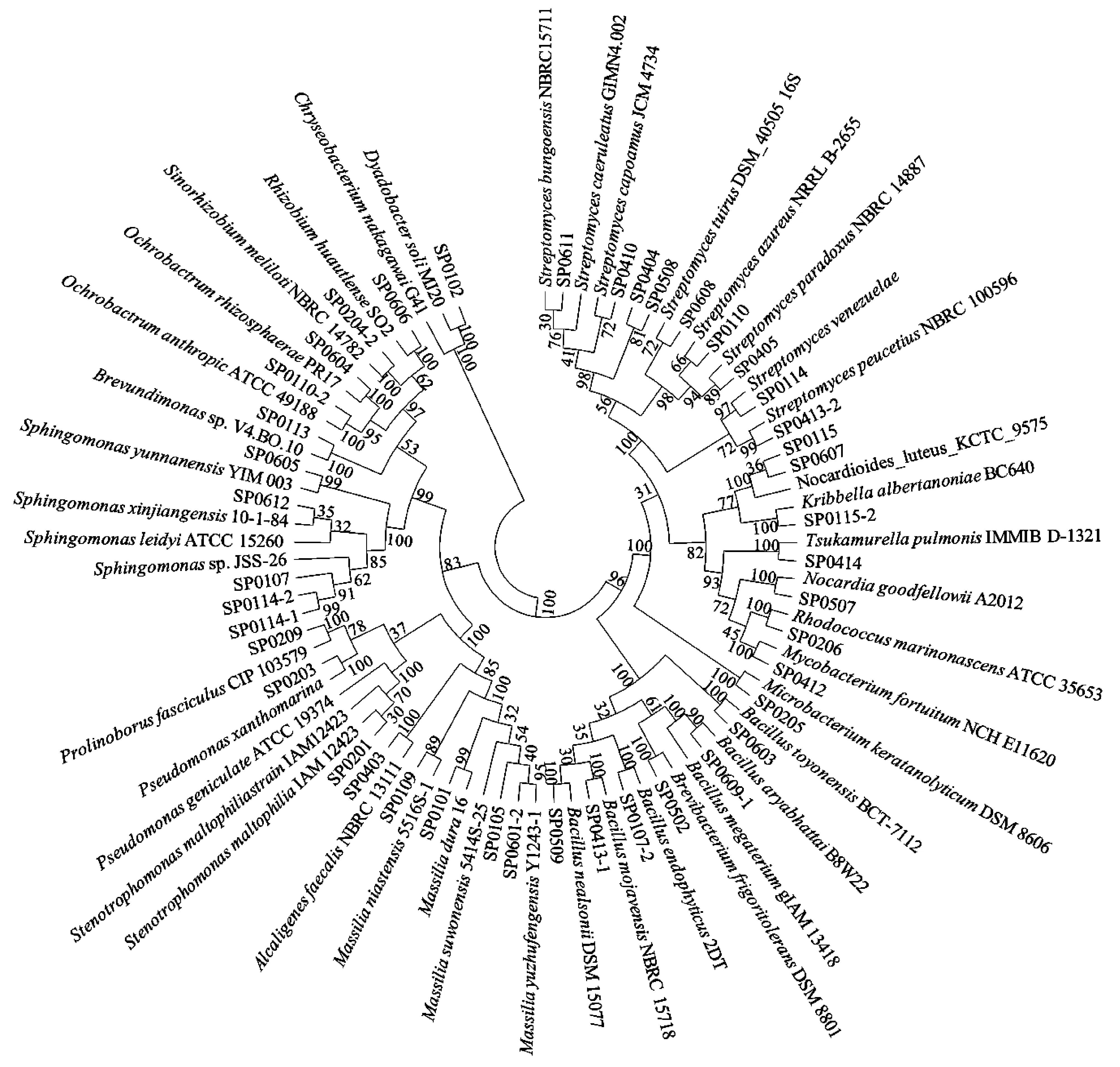
圖1 基于16S rRNA基因序列的根際解磷菌分離株的系統(tǒng)發(fā)育分析
2.2 解無機磷能力
對能使PVK平板變色的23株細菌的解無機磷能力進行測定,各菌株的解無機磷能力為4.20~33.99 μg/mL(見表2). 其中,有8株菌的解無機磷能力大于20 μg/mL,占菌株總數(shù)的30.8%;13株菌的解無機磷能力為10~20 μg/mL,占50%;其余5株的解無機磷能力為4~10 μg/mL,占19.23%;解無機磷能力最強的是霸王SP0205,為33.99 μg/mL.
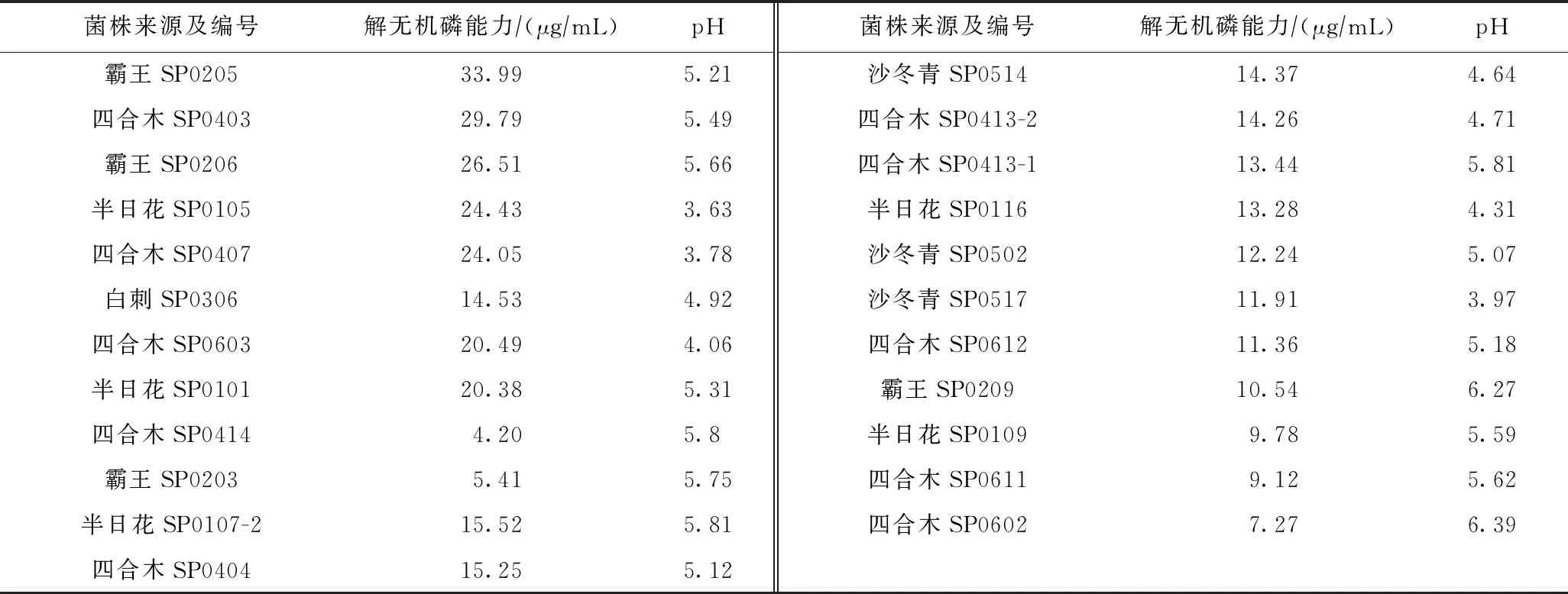
表2 培養(yǎng)液pH及解無機磷能力
2.3 解有機磷能力
該研究中,既能在無機磷培養(yǎng)基上生長,又能在有機磷培養(yǎng)基上生長的PSB共有32株,分屬于鏈霉菌屬(5株)、馬西利亞菌屬(4株)、芽孢桿菌屬(4株)、假單胞菌屬(3株)、蒼白桿菌屬(2株)、中華根瘤菌屬(2株)、寡養(yǎng)單胞菌屬(2株)、產(chǎn)檢桿菌屬(1株)、根瘤菌屬(1株)、鞘氨醇單胞菌屬(1株)、分枝桿菌屬(1株)、細桿菌屬(1株)、紅球菌屬(1株)、韓國生工屬(1株)、類諾卡氏菌屬(1株)、短波單胞菌屬(1株)和短桿菌屬(1株). 其中,鏈霉菌占比(15.62%)較高,其次是芽孢桿菌和馬西利亞菌,各占12.5%. 這32株解有機磷菌的解有機磷能力如表3所示,有4株菌的解有機磷能力高于100 IU,占菌株總數(shù)的10.53%;有7株菌的解有機磷能力為50~100 IU,占18.4%;有17株菌的解有機磷能力為10~50 IU,占44.74%. 其中,解有機磷能力最高的菌株是SP0110-2,高達191.03 IU,具有很好的應用價值.
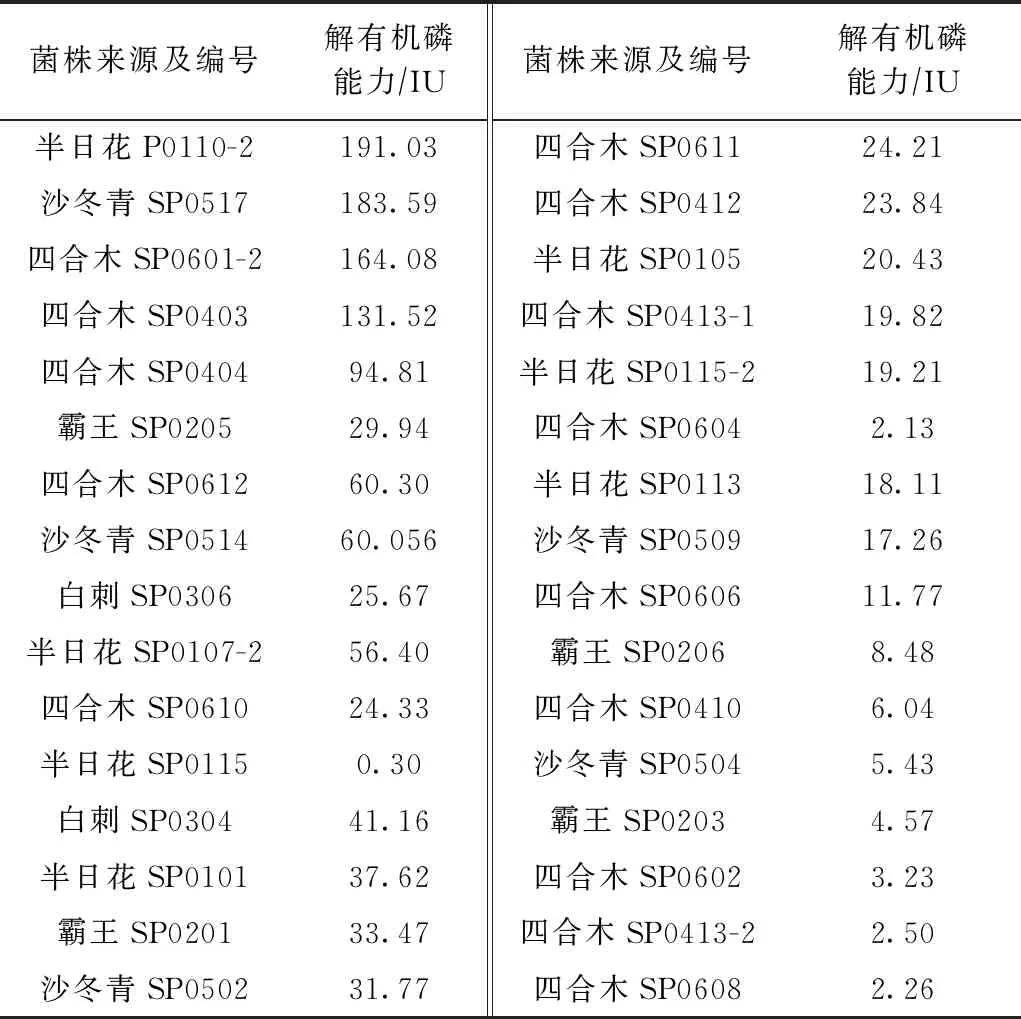
表3 菌株的解有機磷能力
2.4 產(chǎn)鐵載體分析
利用CAS檢測平板對篩選出的根際PSB的產(chǎn)鐵載體能力進行定性測定,其中有22株P(guān)SB的菌落有明顯的橙黃色暈圈,SP0412的D/d值最大,為4.83. 定量測定結(jié)果(見表4)顯示,這些菌株的As/Ar范圍為0.47~0.99;As/Ar為0.4~0.6的有7株(SP0412、SP0403、SP0610、SP0115-2、SP0306、SP0203和 SP0201),產(chǎn)鐵載體能力較強;As/Ar為0.6~0.8的有10株,占供試菌總數(shù)的45.45%,產(chǎn)鐵載體能力中等;As/Ar為0.8~1.0的有5株,占供試菌株總數(shù)的22.73%,產(chǎn)鐵載體能力比低. 產(chǎn)鐵載體菌株占所分離PSB菌株的35.48%,產(chǎn)鐵載體能力較強者的比例為31.82%.

表4 產(chǎn)鐵載體的根際解磷菌
3 討論
細菌是根際土壤微生物中數(shù)量最多的類群,在維持根際土壤的生態(tài)循環(huán)方面起著重要作用[18]. 由于植物根系的分泌物和根的脫落物均會影響植物根部的微生物群落,致使不同植物根際的微生物組成也有很大差別,這也勢必影響PSB的種群結(jié)構(gòu)[19]. 目前已報道的PSB約有30多個菌屬[10]. 該文從內(nèi)蒙古荒漠灌木植物根際分離到62株P(guān)SB,分屬6綱21屬,說明內(nèi)蒙古荒漠灌木有著多樣性高、種類豐富的根際微生物資源. 根際土壤中一些常見的屬種,如芽孢桿菌屬、假單胞菌屬、根瘤菌屬、中華根瘤菌屬、鞘氨醇單胞菌屬及馬西利亞菌屬等在所分離的PSB菌株中都有發(fā)現(xiàn),大部分都是已報道過對植物有促進生長作用的細菌類群,包括解磷、產(chǎn)鐵載體的屬種[10,20];也有較少報道的屬種,如韓國生工屬、冢村氏菌屬. 有研究[21-23]表明,不同植物根際土壤微生物優(yōu)勢菌門(如變形菌門、放線菌門等)組成具有高度相似性. 筆者也發(fā)現(xiàn),放線菌綱是內(nèi)蒙古荒漠灌木根際解磷菌的最優(yōu)勢類群,但放線菌的數(shù)量則很少,這與程冬梅等[20]對新疆胡楊林根際微生物類群的研究結(jié)果一致. 該研究中,芽孢桿菌屬是最優(yōu)勢菌屬,占總分離菌株總數(shù)的17.74%,這與已有報道[24-25]的根際優(yōu)勢菌屬相同,該屬種的菌株是最早發(fā)現(xiàn)的也是解磷效果最好的并且應用最廣泛的PSB類群[26]. Kumar等[27]發(fā)現(xiàn),印度Haryana干旱半干旱地區(qū)的優(yōu)勢菌類群分別屬于厚壁菌門(Firmicutes)和芽孢桿菌屬,這與干旱區(qū)的內(nèi)蒙古荒漠灌木根際PSB菌群有一定差異,而且不同灌木間根際PSB組成差異明顯,這或許與荒漠灌木種類及所處的自然環(huán)境和生態(tài)條件有關(guān),但尚需進一步驗證.
PSB具有較強的根際效應分布,不同土壤類型下PSB的數(shù)量及其解磷能力也不同. 與其他生境植物發(fā)現(xiàn)的根際PSB相比,內(nèi)蒙古荒漠灌木根際PSB的解無機磷能力處于中低等水平,遠低于相關(guān)報道結(jié)果[28-29]. 筆者發(fā)現(xiàn),PSB的解磷量與培養(yǎng)液的pH雖然呈負相關(guān),但相關(guān)性并不顯著,這與已有報道結(jié)果[30]一致,說明非酸類代謝物質(zhì)可能是所分離菌株解磷的主要機制[31]. 筆者還發(fā)現(xiàn),雖然有一些PSB能在無機磷培養(yǎng)基上產(chǎn)生比較明顯的溶磷圈,但定量測定的溶磷量則較低甚至出現(xiàn)負數(shù),這可能是PSB在分解難溶的Ca3(PO4)2過程中,由于溶解產(chǎn)生的可溶性磷被PSB自身細胞吸收或貯藏利用所導致.
對于有機磷降解菌的報道多為芽孢桿菌和假單胞菌[32]. 該研究篩選出的32株解有機磷菌中鏈霉菌最多(15.62%),其次為芽孢桿菌和假單胞菌(12.5%),與上述報道相似. 這32株解有機磷PSB的解有機磷能力高于100 IU的有4株,在50~100 IU之間的有7株,解有機磷能力最強的菌株是SP0110-2,高達191.03 IU,遠高于相關(guān)報道結(jié)果[33-34],處于較高水平. 該研究中發(fā)現(xiàn)具有解有機磷能力的菌株有32株,達到所篩菌株總數(shù)的51.62%,共分屬于17個菌屬,表明荒漠環(huán)境下的解有機磷PGPR具有豐富的多樣性,這與荒漠灌木的耐貧瘠、耐干旱的能力息息相關(guān).
在低鐵環(huán)境中,許多PGPR可產(chǎn)生鐵載體,可以螯合環(huán)境中溶解的鐵加以吸收,有效限制了土壤中可溶性鐵的供應,致使病原真菌產(chǎn)生鐵缺乏,引發(fā)麥角甾醇合成途徑受阻,抑制細胞膜合成,產(chǎn)生物質(zhì)滲漏[35],使其死亡. 所以具有產(chǎn)鐵載體能力的細菌不僅具有植物促生能力,還能增強植物抗病原菌能力. 對具有產(chǎn)鐵載體能力的PSB的研究表明,來自不同灌木植物根際的產(chǎn)鐵載體PSB有著明顯的多樣性. 在系統(tǒng)發(fā)育上,22株產(chǎn)鐵載體菌株分屬于14個菌屬,占全部供試菌總數(shù)的35.48%,芽孢桿菌屬有6株,占總產(chǎn)鐵載體菌株數(shù)量的27.27%,為優(yōu)勢菌屬,不同于朱彭玲等[36]的研究結(jié)果. 目前,有關(guān)解有機磷細菌具有產(chǎn)鐵載體能力的報道還較少[32],而筆者篩選到的具產(chǎn)鐵載體能力的有機磷降解菌多達18株,占菌株總數(shù)的81.82%,且產(chǎn)鐵載體能力較強的菌株(占總數(shù)的31.82%)全部是解有機磷細菌. 這些研究結(jié)果表明,內(nèi)蒙古荒漠可成為具有多種植物較高促生長能力的解有機磷根際促生菌的重要來源.
4 結(jié)論
a) 內(nèi)蒙古荒漠地區(qū)蘊藏著較為豐富的PSB菌種資源. 從西鄂爾多斯荒漠5種灌木根際分離到62株P(guān)SB,分屬α-變形菌綱、β-變形菌綱、γ-變形菌綱、芽孢桿菌綱、放線菌綱和噬纖維菌綱,其中放線菌綱為最優(yōu)勢菌綱;分屬21個菌屬,其中芽孢桿菌屬為最優(yōu)勢菌屬;不同灌木間PSB組成差異比較明顯,四合木根際PSB種類最為豐富,而白刺根際PSB種類最少.
b) 分離到的PSB解無機磷能力處于中低等水平,解無機磷能力在4.20~33.99 μg/mL之間;解有機磷的PSB菌株占比較大,且大部分(解有機磷能力高于50 IU的占比為47.83%)解有機磷能力較強,最高可達191.03 IU,處于較高等水平;有22株具有產(chǎn)鐵載體能力,且產(chǎn)鐵載體能力較強的菌株(占比為31.82%)全部為解有機磷細菌,是解有機磷PGPR的重要來源之一.
猜你喜歡
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學生數(shù)理化·七年級數(shù)學人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(shè)(2018年6期)2018-08-16 07:23:10
新高考(英語進階)(2018年1期)2018-04-18 14:00:11
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
