甘肅省5 種昆蟲病原線蟲對草地貪夜蛾的致病力測定
2021-11-22 09:50:04楊淋凱朱小芳錢秀娟劉衛紅姜紅霞劉長仲
草業科學 2021年10期
楊淋凱,朱小芳,錢秀娟,劉衛紅,姜紅霞,劉長仲
(1. 甘肅農業大學植物保護學院,甘肅 蘭州 730070;2. 甘肅省植保植檢站,甘肅 蘭州 730020)
草地貪夜蛾(Spodoptera frugiperda)屬鱗翅目夜蛾科灰翅夜蛾屬[1-3],是一種原發生于美洲熱帶和亞熱帶地區的多食性及遷飛性農業害蟲,因其生存能力強,擴展速度快,目前已侵入非洲和亞洲多個國家和地區[4]。我國于2019 年1 月在云南首次發現草地貪夜蛾,此后迅速蔓延至多個省份。甘肅省于2019年7 月2 日首次發現草地貪夜蛾,截止2019 年年底已在甘肅東部28 個縣區發生,為害玉米(Zea mays)約266.67 hm2[5-6]。有研究證明入侵我國的草地貪夜蛾均為“玉米型”[7],主要危害玉米,其次是高粱(Sorghum bicolor)、谷子(Setaria italica)、小麥(Triticum aestivum)、甘蔗(Saccharum officinarum)等15 種作物,甚至也能在皇竹草(Pennisetum sinese)、馬唐(Digitaria sanguinalis)、牛筋草(Eleusine indica)、蘇丹草(Sorghum sudanense)等禾本科草類植物上生存[8]。玉米是我國主要的糧食和飼料作物,而高粱的營養成分類似于玉米,且成本更低,經加工可做成優良的飼料,減緩草地畜牧業飼草供應緊缺的壓力[9]。玉米和高粱是草地貪夜蛾的主要寄主,其大規模危害會造成玉米和高粱的嚴重減產,這無疑對糧食安全生產和草地畜牧業發展構成了極大威脅。目前國內對該害蟲的防治以幾種殺蟲活性強的新型化學殺蟲劑為主,如甲氨基阿維菌素苯甲酸鹽、氯蟲苯甲酰胺、乙基多殺菌素及相關的復合制劑,并在生產上取得了一定的防治效果[10-12],但長期大量施用化學農藥不僅污染環境、害蟲容易產生抗藥性,同時還增加了飼料農藥超標和畜產品品質下降的風險。在生物防治方面,草地貪夜蛾也已經對轉Bt 毒蛋白基因的作物及Bt 殺蟲制劑產生抗性[13],因此尋求新的生防制劑具有重要的意義。
昆蟲病原線蟲(entomopathogenic nematodes, EPN)包括斯氏屬(Steinernema)和異小桿屬(Heterorhabditis)兩類[14],具有寄主范圍廣及主動搜尋能力、對環境無污染、對人畜安全、易培養、成本低、可與農藥混配施用等優點[15]。作為重要的生防資源,昆蟲病原線蟲已被用于防治蠐螬(Scarabacidae)、韭蛆(Bradysia odoriphaga)、小地老虎(Agrotis ypsilon)等多種農林、草地、花卉以及衛生害蟲[16-17]。使用昆蟲病原線蟲防控草地貪夜蛾已成為國內外的研究熱點。昆蟲病原線蟲對寄主昆蟲有很強的專化性,不同種類甚至品系對相同寄主的致病力不同[18],因此正確選擇適宜種類及品系是防治成功的重要因素。
甘肅氣候干燥,少雨且晝夜溫差大,外地引進的昆蟲病原線蟲難以適應當地的環境,而土著昆蟲病原線蟲早已適應當地的條件。因此,應用土著昆蟲病原線蟲具有短時間內完成定殖、建立穩定種群、更好地發揮持續高效控害等特點。為探索甘肅省昆蟲病原線蟲資源對防控草地貪夜蛾的潛力,拓展其在害蟲生物防治中的應用,本研究利用從甘肅省采集分離獲得的5 種10 個昆蟲病原線蟲品系,通過室內生物測定,篩選出對草地貪夜蛾具有高效生防潛力的昆蟲病原線蟲品系,為有效利用甘肅省昆蟲病原線蟲資源防治草地貪夜蛾提供理論依據。
1 材料與方法
1.1 材料
供試昆蟲:草地貪夜蛾采自甘肅省白銀市靖遠縣,在甘肅農業大學昆蟲生態實驗室飼養。
供試線蟲:夜蛾斯氏線蟲(Steinernema feltiae)0639C、0657YM、0661YM、0663PG、0663TD、0619HT品系,斯氏線蟲(Steinernema karil) 0663YC 品系,卡森斯氏線蟲(Steinernema krussei) 0657L 品系,短尾異小桿線蟲(Heterorhabditis brevicaudis) 0641TY 品系,大異小桿線蟲(Heterorhabditis megidis) 0627M 品系。線蟲10 個品系均采自甘肅省內西峰市、天水市、武威市民勤縣、白銀市靖遠縣和景泰縣等具有代表性的市縣[18],采用大蠟螟(Galleria mellonella)誘集法分離并收集線蟲[19],經品系鑒定后保存于甘肅農業大學昆蟲生態實驗室。定期以韭蛆(Bradysia odoriphaga)為寄主于無菌土壤中復壯以保證品系活力。試驗所用線蟲懸浮液為新鮮培養,由大蠟螟活體擴繁而得,保存于4 ℃冰箱內待用。
1.2 致病力測定
以草地貪夜蛾2 齡幼蟲作為供試昆蟲,篩選出致病力較高的昆蟲病原線蟲種類和品系,用致病力較高的昆蟲病原線蟲侵染草地貪夜蛾,并測定其對草地貪夜蛾的致病力。
1.2.1 甘肅省昆蟲病原線蟲種類(品系)篩選
在24 孔細胞培養板中處理,每個孔加入1 頭草地貪夜蛾2 齡幼蟲并用脫脂棉和濾紙進行保濕,12 個孔為一個處理,用移液槍吸取200 頭線蟲侵染的溶液滴加在每個孔內,1 個品系線蟲為1 個處理,共10 個線蟲處理,同時以等量無菌水作對照,每處理重復3 次,孔板內放置足量的草地貪夜蛾人工飼料[20]。然后將24 孔板放回人工氣候箱,每隔12 h 檢查草地貪夜蛾的死亡情況并記錄。控制條件為溫度24~26 ℃,相對濕度70%,光照 ∶ 黑暗16 h ∶ 8 h。
1.2.2 高效昆蟲病原線蟲品系對草地貪夜蛾2 齡幼蟲的致病力測定
對篩選出的高效品系進行致病力測定,在24 孔細胞培養板中處理,每個孔加入1 頭草地貪夜蛾2 齡幼蟲并用脫脂棉和濾紙保濕,12 個孔為一個處理。每種線蟲設6 個數量梯度,使線蟲與草地貪夜蛾2 齡幼蟲數量比分別為5 ∶ 1、10 ∶ 1、20 ∶ 1、40 ∶ 1、80 ∶ 1、160 ∶ 1,同時以等量無菌水處理作對照,每處理重復3 次,孔板內放置足量的草地貪夜蛾人工飼料。然后將24 孔板放回人工氣候箱,每隔12 h 檢查草地貪夜蛾的死亡情況并記錄。控制條件為溫度24~26 ℃,相對濕度70%,光照 ∶ 黑暗16 h ∶ 8 h。
1.2.3 高效昆蟲病原線蟲品系對草地貪夜蛾預蛹的致病力測定
在9 cm 的培養皿中墊2 層濾紙,加入適量蒸餾水保持濕潤,每皿放入1 頭草地貪夜蛾預蛹,設200、400、800、1 600、3 200 IJs·H?15 個線蟲數量梯度,用移液槍吸取各濃度的線蟲直接滴加到預蛹體壁上,每處理10 頭,3 次重復,以無菌水處理作對照。然后將培養皿放回人工氣候箱,檢查化蛹情況并記錄。
1.2.4 高效昆蟲病原線蟲品系對草地貪夜蛾蛹的致病力測定
在帶蓋的小塑料盒內裝入適量蛭石并加蒸餾水保持濕潤,每個盒子中埋入1 頭正常化蛹的供試昆蟲蛹,用移液槍滴加各梯度的線蟲,使線蟲與草地貪夜蛾蛹的數量比分別為800 ∶ 1、1 600 ∶ 1、3 200 ∶ 1、6 400 ∶ 1、12 800 ∶ 1,每處理10 頭,3 次重復,以無菌水處理作對照,操作完成后放回人工氣候箱觀察羽化情況并記錄。
1.3 數據分析
采用SPSS 26.0 軟件對所測數據進行統計分析,用平均值 ± 標準誤表示測定結果。使用 One-Sample Kolmogorov-Smirnov test 對數據進行正態分布檢驗。符合正態分布的使用 Lev-ene’s test 進行方差齊性檢驗,若齊性不存在,則對百分率數據進行反正弦轉換。對本試驗數據進行檢驗表明均符合方差齊性,直接進行單因素方差分析,并使用 LSD test 進行多重比較。
2 結果與分析
2.1 甘肅省昆蟲病原線蟲優良種類(品系)的篩選
不同昆蟲病原線蟲種類間對草地貪夜蛾2 齡幼蟲的致死率差異較大(表1),夜蛾斯氏線蟲對草地貪夜蛾2 齡幼蟲致病力最強,卡森斯氏線蟲次之。幼蟲被昆蟲病原線蟲各品系侵染24 h 后均有一定的死亡率,其中夜蛾斯氏線蟲0663PG 為63.90%,顯著高于其他昆蟲病原線蟲種類(P< 0.05)。夜蛾斯氏線蟲的0657YM、0663TD、卡森斯氏線蟲0657L 和斯氏線蟲(S. karil) 0663YC 的致病死亡率在31%~43%,但其間差異不顯著(P> 0.05)。侵染48 h 后,各品系對寄主的致死作用均有所增加,其中夜蛾斯氏線蟲 0663PG 為88.90%,與其他昆蟲病原線蟲種類存在顯著差異(P< 0.05)。侵染72 h 后,除夜蛾斯氏線蟲0619HT 和短尾異小桿線蟲0641TY 外,其余所有昆蟲病原線蟲品系的累積死亡率均在50%以上。大異小桿線蟲 0627M 和夜蛾斯氏線蟲0663TD的致死率為75%和72.23%,兩者間差異不顯著(P>0.05)。夜蛾斯氏線蟲 0663PG 和卡森斯氏線蟲 0657L的致死率分別達到100%和83.30%,兩者間差異不顯著(P> 0.05)。可見,夜蛾斯氏線蟲 0663PG 和卡森斯氏線蟲0657L 為對草地貪夜蛾幼蟲致病力較強的昆蟲病原線蟲品系。
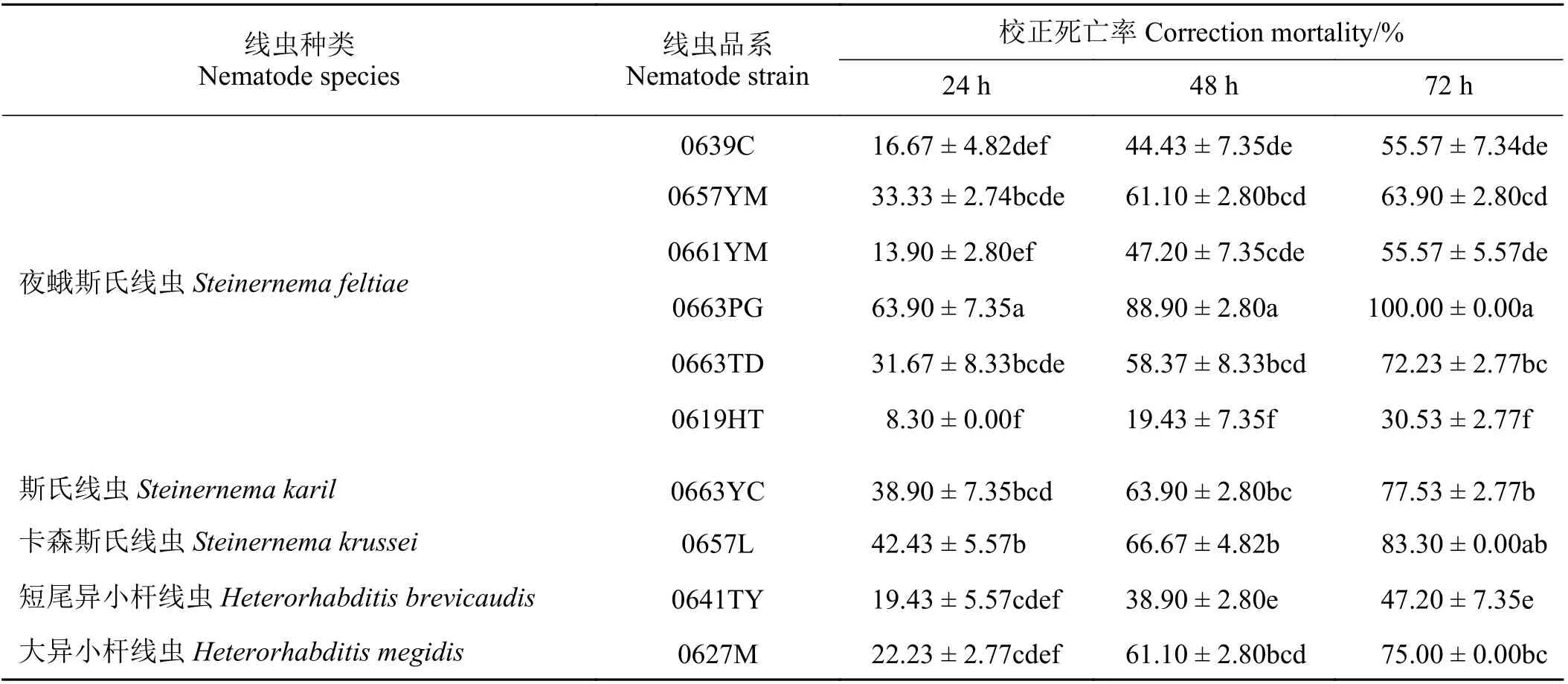
表1 10 個昆蟲病原線蟲品系對草地貪夜蛾2 齡幼蟲的致病力Table 1 Virulence of 10 entomopathogenic nematodes strains on 2nd instar larvae of S. frugiperda
夜蛾斯氏線蟲的不同品系對草地貪夜蛾2 齡幼蟲的致死率存在明顯差異。在夜蛾斯氏線蟲的6 個供試品系中,侵染24 h 后,0663PG 的致死率為63.90%,與其余品系之間存在顯著差異(P< 0.05)。侵染72 h 后,0663PG 的累計死亡率達到100%,0663TD、0657YM、0639C、0661YM 和0619HT 分 別為75.00%、63.90%、55.57%、55.57%和30.53%。
2.2 2 種高效昆蟲病原線蟲對草地貪夜蛾的致病力
草地貪夜蛾2 齡幼蟲被侵染24 h 后,夜蛾斯氏線蟲0663PG 與幼蟲數量比為160 ∶ 1 和80 ∶ 1 時,其致死率均為38.90%,顯著高于其他線蟲濃度(P<0.05)。侵染48 h 時,數量比為160 ∶ 1 和80 ∶ 1 時死亡率分別為63.90%和52.77%,兩者差異不顯著(P>0.05)。侵染72 h 時,其死亡率分別達到100%和61.10%,兩者間存在顯著差異(P< 0.05),也顯著高于其他線蟲濃度(P< 0.05) (表2)。
卡森斯氏線蟲0657L 侵染24 h 后,數量比為160 ∶ 1 和80 ∶ 1 時,其致死率均為27.77%。侵染48 h后,數量比為160 ∶ 1 和80 ∶ 1 死亡率較高,分別為61.10%和50.57%;處理72 h 后其死亡率分別為80.53%和58.33%,兩者間差異顯著(P< 0.05) (表2)。
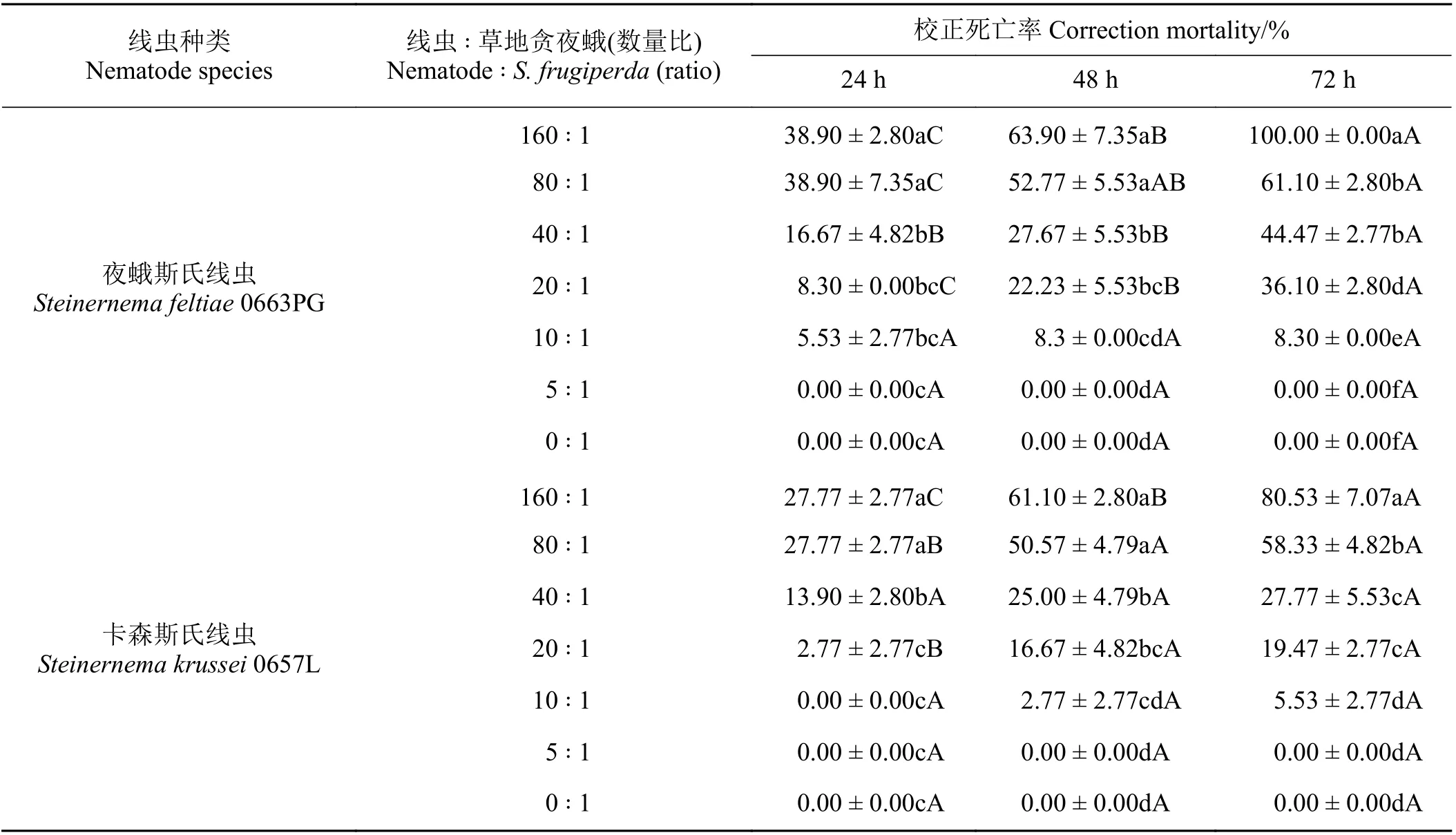
表2 夜蛾斯氏線蟲0663PG 品系和卡森斯氏線蟲0657L 品系對草地貪夜蛾 2 齡幼蟲的致死作用Table 2 Steinernema feltiae 0663PG and Steinernema krussei 0657L against Spodoptera frugiperda 2nd instar larvae
2.3 夜蛾斯氏線蟲0663PG 品系對草地貪夜蛾預蛹的致病力測定
5 個數量梯度處理下的預蛹2 d 后均可成功化蛹,侵染13 d 后處理組和對照組開始出現草地貪夜蛾蛹羽化,處理15 d 后,夜蛾斯氏線蟲0663PG 與草地貪夜蛾預蛹數量比為1 600 ∶ 1 和3 200 ∶ 1 時,蛹的羽化率為96.67%,其余梯度處理下的羽化率均為100% (表3)。
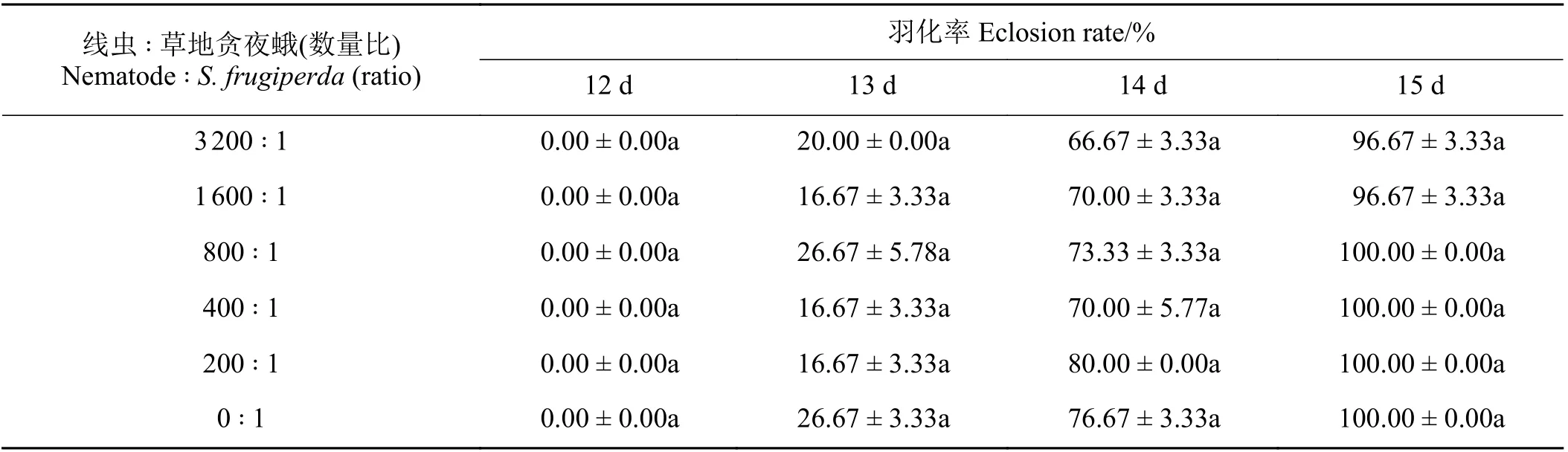
表3 夜蛾斯氏線蟲0663PG 品系對草地貪夜蛾預蛹的致病力Table 3 Steinernema feltiae 0663PG against Spodoptera frugiperda prepupa
2.4 夜蛾斯氏線蟲0663PG 品系對草地貪夜蛾蛹的致病力測定
5 個數量梯度下夜蛾斯氏線蟲0663PG 在侵染13 d 后對草地貪夜蛾蛹的致死作用不明顯,數量比為6 400 ∶ 1 和12 800 ∶ 1 時,蛹的羽化率為93.33%。對未羽化的蛹解剖發現確有線蟲寄生,但各梯度下的線蟲對蛹的致死率低于10%。觀察結果表明,昆蟲病原線蟲不適合在蛹期進行防治,且線蟲對草地貪夜蛾蛹的羽化時間并沒有影響(表4)。
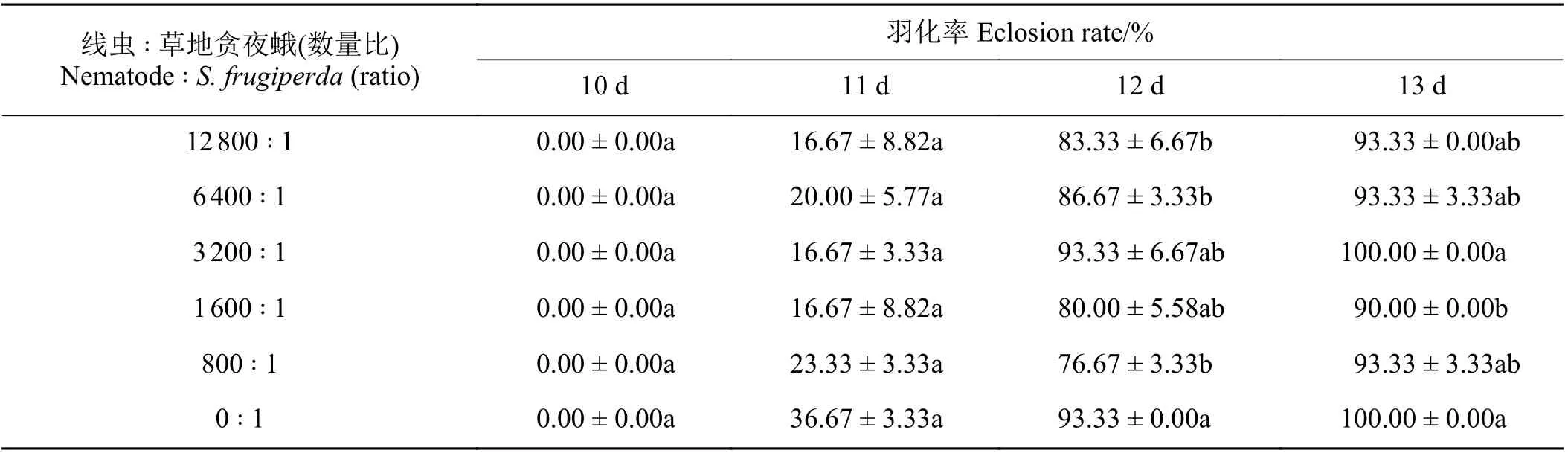
表4 夜蛾斯氏線蟲0663PG 品系對草地貪夜蛾蛹的致病力Table 4 Steinernema feltiae 0663PG against Spodoptera frugiperda pupa
3 討論與結論
本研究發現,夜蛾斯氏線蟲0663PG 品系對草地貪夜蛾的致病力較高,處理48 h 時,侵染數量比為160 ∶ 1 時,其死亡率達到63.90%。梁銘榮等[21]研究發現,小卷蛾斯氏線蟲AⅡ和長尾斯氏線蟲X-7品系處理36 h 時,侵染數量比為50 ∶ 1 時,對草地貪夜蛾2 齡幼蟲的致死作用達到92%和80%。通過結果比對分析,夜蛾斯氏線蟲0663PG 致病力低于小卷蛾斯氏線蟲AⅡ和長尾斯氏線蟲X-7 品系,說明不同種類及品系對同一種寄主昆蟲的致病力存在差異。這可能與不同種間線蟲的侵染方式有關,夜蛾斯氏線蟲0663PG 屬于中間型線蟲[22],而小卷蛾斯氏線蟲AⅡ和長尾斯氏線蟲X-7 品系屬于伏擊型侵染策略的線蟲,草地貪夜蛾2 齡幼蟲比較活躍,行動速度快,更適合伏擊型線蟲侵染。錢秀娟等[23]在室內測定下,夜蛾斯氏線蟲0619HT 對大蠟螟的致死作用最強,處理72 h 時,侵染數量比為60 ∶ 1 時,累計死亡率為96.7%。而本研究發現,0619HT在處理72 h 時,侵染數量比為200 ∶ 1 時,累計死亡率為30.53%,與供試的其他9 個線蟲品系相比致病力最低,說明同種昆蟲病原線蟲品系對不同的寄主昆蟲致病力存在差異。昆蟲病原線蟲的致病力會因寄主昆蟲的改變而不同,因此,昆蟲病原線蟲的室內篩選和致病力評價在生產應用之前很有必要。
國外對于昆蟲病原線蟲防控草地貪夜蛾的研究早于國內,許多研究表明昆蟲病原線蟲具有防治草地貪夜蛾的應用潛力。Lezama-Gutierrez 等[24]和Molina-Ochoa 等[25]調查發現,斯氏線蟲是土壤中分布最廣的草地貪夜蛾天然病原物。Andaló等[26]在室內條件下測定了17 個巴西本地線蟲品系對草地貪夜蛾的致病力, 最終得到S. arenarium和Heterorhabditissp. RSC02 兩個高效品系。本研究以甘肅本地10 個昆蟲病原線蟲品系進行室內篩選和致病力評測,發現夜蛾斯氏線蟲0663PG 的毒力最高,當0663PG 與草地貪夜蛾數量比為80 ∶ 1 時,72 h 后草地貪夜蛾幼蟲死亡率在60%以上,當侵染劑量增加到160 ∶ 1時,草地貪夜蛾的死亡率達到100%,本研究認為,夜蛾斯氏線蟲0663PG 品系在草地貪夜蛾的生物防治方面有著巨大潛力,可以在后續研究中與化學藥劑組合使用,彌補線蟲單獨使用時見效慢的缺陷[27]。基于此,尋求高效低毒的殺蟲劑與夜蛾斯氏線蟲0663PG 混配防治草地貪夜蛾還有待研究。
本研究在室內測定了甘肅省5 種10 個線蟲品系對草地貪夜蛾2 齡幼蟲的致病力,篩選出致病力較好的夜蛾斯氏線蟲0663PG 和卡森斯氏線蟲0657L 兩個品系。在低劑量下對以上2 種線蟲進行致病力測定,夜蛾斯氏線蟲0663PG 與草地貪夜蛾數量比為160 ∶ 1 時,處理48 h 后致死率為63.90%,與卡森斯氏線蟲0657L 的致死率相近,但處理72 h后草地貪夜蛾2 齡幼蟲死亡率達到100%,同樣數量比下卡森斯氏線蟲0657L 對2 齡幼蟲的致死率為80%,夜蛾斯氏線蟲0663PG 的致病力高于卡森斯氏線蟲0657L。在大劑量下用夜蛾斯氏線蟲0663PG侵染草地貪夜蛾預蛹和蛹,預蛹在線蟲處理下2 d后成功化蛹,預蛹和蛹分別在侵染15 d 和13 d 后完成羽化,但夜蛾斯氏線蟲0663PG 對預蛹和蛹的致死率低于10%。本研究結果表明,草地貪夜蛾預蛹和蛹對昆蟲病原線蟲不敏感,線蟲侵染后對蛹的羽化時間沒有顯著影響(P> 0.05),故昆蟲病原線蟲并不適合在草地貪夜蛾預蛹和蛹期進行防治。
