小白菜種子休眠破除方法研究
2021-11-29 07:07:20耿啟華宋莉萍林處發張余洋汪愛華
湖北農業科學 2021年21期
耿啟華,宋莉萍,林處發,張余洋,汪愛華
(1.武漢市農業科學院,武漢 430065;2.華中農業大學,武漢 430070)
種子休眠是指具有生活力的種子在各種外在或內在因素影響下,即使在適宜的萌發條件下也無法正常發芽的現象[1-5]。大多數種子均存在休眠現象,主要是由于種皮限制、存在外源抑制物質、胚未完全成熟等因素所導致[2,3,6,7]。青格樂等[2]、李楠[4]、楊鷺生等[5]、王亞茹等[8]研究認為,休眠是植物在長期的系統發育過程中所獲得的用于抵抗不良環境的一種適應性,是植物所特有的、重要的環境適應性之一;但是,種子休眠對于農業的發展、生產以及實驗研究來說卻存在很不利的影響,若種子在播種時仍處于休眠狀態則無法確定種子的播種量和播種期,會對生產和實驗研究造成很大影響[2,9]。因此,解決種子休眠的問題對于農業生產以及科研來說極為重要。目前,已有很多有關種子休眠方面的研究,但由于影響種子休眠的因素眾多,影響機制復雜,因此需要進一步探索打破種子休眠的方法[2,4,10]。
小白菜(Brassica rapassp.Chinensis)是十字花科主要蔬菜之一,在大部分地區均有種植。對于種子貯藏來說,休眠可以有效延長小白菜種子的貯藏期限;但對于當年采收的商品性種子來說,休眠會導致種子延遲上市,影響種子的銷售以及種植[9]。在發芽預試驗中發現,在實驗室提供的眾多種子中多數品種均存在休眠現象,但M62和漢優休眠較為嚴重,且小白菜種子的休眠會隨著貯藏時間的延長而解除。為此,試驗通過不同的物理、化學及激素處理方法探尋快速打破小白菜種子休眠的方法,以加快試驗研究的進度。
1 材料與方法
1.1 試驗材料
試驗用種子由武漢市農業科學院蔬菜研究所十字花科研究室提供。種子于2020年4月27日采收,選取飽滿純凈的漢優、M62兩種小白菜種子。
所用藥劑與儀器包括氫氧化鈉(國產,分析純)、純濃硫酸、硝酸鉀(國產,分析純)、過氧化氫(國產)、赤霉素(GA,Coolaber)、6-BA(SIGMA)、油菜素內酯(BR,SIGMA);烘干箱PYL-70(天津市萊玻特瑞儀器設備有限公司)、RTOP型智能型人工氣候箱(浙江托普儀器有限公司)、容聲冰箱(BCD-249WD11DY)、培養皿(biosharp,90 mm×15 mm)、9 mm濾紙。
1.2 試驗方法
種子發芽試驗于武漢市農業科學院北部園區院蔬菜研究所十字花科研究室實驗室開展,所有處理均于2020年4月29日開始進行。利用培養皿法,將各處理后的種子放置于鋪有濾紙的培養皿中,加入去離子水保持濾紙處于濕潤狀態,然后將培養皿置于室溫下培養,每天統計種子發芽數,共4 d[11]。以未經處理的種子為對照,每個處理100粒,設3次重復。
1.2.1 化學處理
1)H2O2浸種處理:分別以濃度1%、2%、4%、6%、8%、10%、12%的H2O2浸種24 h,沖洗5次以上進行種子發芽試驗。
2)KNO3浸 種 處 理:設 置0.1%、0.2%、0.4%、0.8%、1.6%、3.2%濃度梯度,分別浸種12、24 h,沖洗5次以上進行種子發芽試驗。
3)純濃硫酸浸種處理:用濃硫酸分別浸種5、10、15、20、25 min,沖洗至pH約為7后進行種子發芽試驗。
4)赤霉素浸種處理:設置200、400、800、1 000、2 000、2 500、3 000 mg/L的GA溶液,分別浸種12、24、48 h,沖洗干凈進行發芽試驗。
5)6-BA浸種處理:設置不同的濃度(50、100、200、300 mg/L),分別浸種24、48 h,沖洗后進行發芽試驗。
1.2.2 引發處理 以400 mg/L赤霉素、0.05%油菜素內酯(BR)、10% H2O2、10%NaOH、1% KNO3分別浸種24 h,沖洗干凈后30℃烘干12 h,烘干后進行發芽試驗。
1.2.3 變溫處理 先將兩種小白菜種子在4、10、20℃條件下貯藏7 d,取出后分別在40、45、50℃條件下處理1 h,之后繼續放置在4、10、20℃條件下貯藏48 h,取出后進行發芽試驗。
1.3 數據統計與處理
利用Excel對數據進行匯總,計算種子發芽率并繪制圖表,發芽率=發芽數/供試種子數×100%;同時利用SPSS 23進行數據分析,用LSD法進行多重比較。
2 結果與分析
2.1 化學處理種子發芽情況
2.1.1 H2O2浸種處理種子發芽情況從圖1可以看出,H2O2浸種24 h種子發芽率整體呈現先升高后降低的趨勢,H2O2濃度為2%和6%時可有效打破M62、漢優兩種小白菜種子的休眠,種子發芽率達到80%以上,但2%的濃度對打破兩種種子休眠的影響更為均衡;當H2O2溶液濃度高于8%時,種子發芽率逐漸降低。
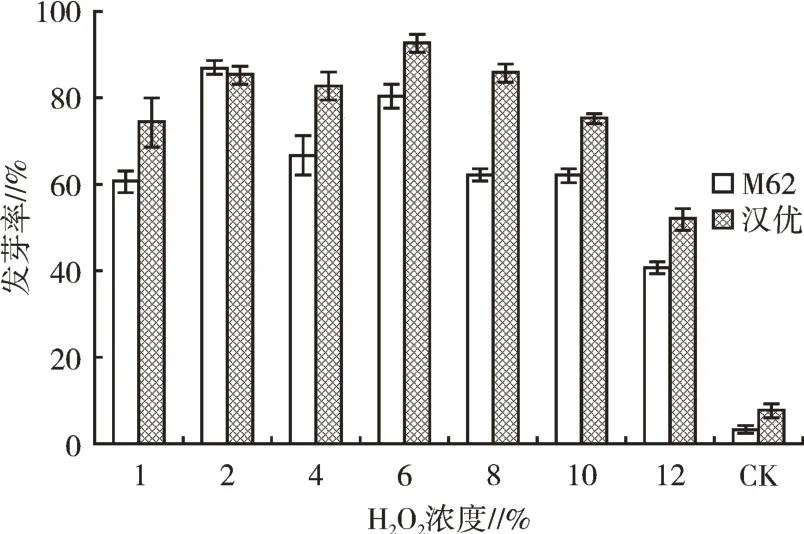
圖1 H2O2浸種24 h對種子發芽的影響
2.1.2 KNO3浸種處理種子發芽情況方差分析結果(表1)顯示,在一定濃度下,相同濃度KNO3浸種12、24 h,種子發芽率差異存在明顯差異,但觀察發現處理12 h與處理24 h的種子生長并無明顯區別;不同濃度KNO3浸種相同時間后其發芽率存在明顯差異,兩種種子發芽率均呈現先升高后降低的趨勢,當KNO3濃度為0.8%、1.6%時對打破兩種小白菜種子的休眠效果最顯著,種子發芽率達到95%以上;而3.2%的KNO3溶液打破休眠的效果較差。

表1 KNO3浸種對M62和漢優種子發芽的影響
2.1.3 濃硫酸浸種處理種子發芽情況試驗(圖2)發現,經濃硫酸浸泡的種子種皮大量破裂,種子發芽率達到98%以上,說明濃硫酸浸種可在不傷及種胚的情況下有效打破小白菜種子休眠。但是觀察種子后續生長情況發現,濃硫酸處理后的種子發芽后長勢較弱,有些幼苗子葉易爛;由此可見,雖然濃硫酸可以快速有效地打破小白菜種皮的限制,解除休眠,但對種子的后續生長不利,因此濃硫酸處理并不是打破小白菜種子休眠的最佳方法。
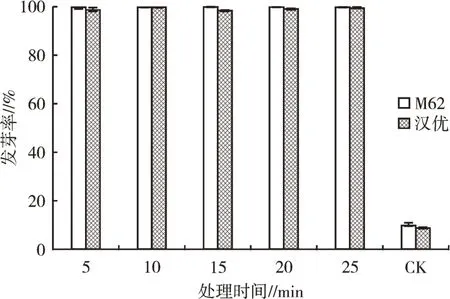
圖2 濃硫酸不同處理時間對種子發芽的影響
2.1.4 赤霉素浸種處理種子發芽情況 試驗結果(表2)表明,在一定濃度范圍內,赤霉素可有效打破漢優、M62兩種小白菜種子的休眠,不同處理時間以及不同處理濃度對種子發芽的影響均有明顯差異。但經多重比較發現,在一定濃度范圍內,相同濃度下處理12 h和24 h種子發芽率差異并不明顯;赤霉素濃度低于1 000 mg/L時,浸種12、24 h均可達到較好打破休眠的效果,幼苗長勢優良,無明顯差異;隨著赤霉素溶液濃度的升高,兩種種子發芽率均有所降低,其中浸種12 h的種子發芽率快速降低,而浸種24 h的兩種種子在赤霉素溶液濃度為2 000 mg/L時發芽率均達到90%以上,高于浸種12、48 h種子發芽率,且發芽整齊,幼苗長勢健壯,子葉濃綠。赤霉素浸種48 h打破休眠效果較差,易生霉、爛種。綜合對比發現,浸種24 h,且赤霉素濃度低于1 000 mg/L時打破小白菜種子休眠效果最佳。

表2 赤霉素處理對漢優和M62種子發芽的影響
2.1.5 6-BA浸種處理種子發芽情況從圖3可以看出,6-BA濃度為50 mg/L時,浸種24、48 h處理下,M62種子和漢優種子發芽率均較高,且兩種種子幼苗長勢良好;隨著6-BA濃度的增大,兩種小白菜種子發芽率呈現明顯的下降趨勢,當6-BA濃度為300 mg/L時,兩種種子發芽率均為0。綜上所述,50 mg/L 6-BA浸種處理可有效打破M62和漢優種子的休眠。
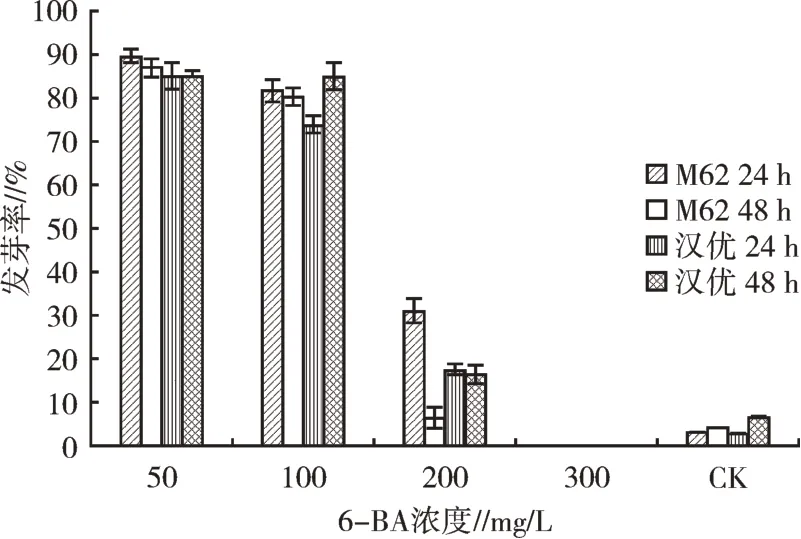
圖3 6-BA浸種對種子發芽的影響
2.2 引發處理種子發芽情況
從圖4可以看出,經400 mg/L赤霉素溶液引發處理的M62種子發芽率達到98%,漢優種子發芽率達到100%;1% KNO3溶液處理的兩種小白菜種子發芽率均達到100%,且幼苗長勢健壯;10% H2O2溶液也可有效打破休眠,M62種子發芽達到70%,漢優則達到87%;而經0.05%BR、10%NaOH處理的種子則完全未發芽;綜合引發處理種子發芽情況,400 mg/L赤霉素溶液、1% KNO3溶液浸種24 h,沖洗干凈后30℃烘干12 h可有效打破小白菜種子的休眠。
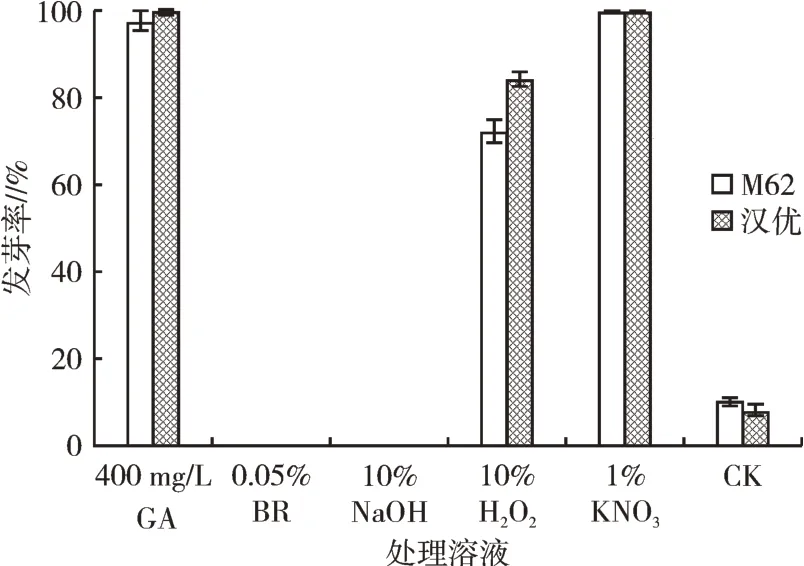
圖4 引發處理對種子發芽的影響
2.3 變溫處理種子發芽情況
從圖5可以看出,將兩種小白菜種子置于20℃條件下貯藏7 d,取出后在50℃條件下處理1 h,之后繼續放置于20℃條件下貯藏48 h可有效打破小白菜種子休眠,兩種種子發芽率均達到80%以上;4℃和10℃變溫處理打破休眠效果較差。
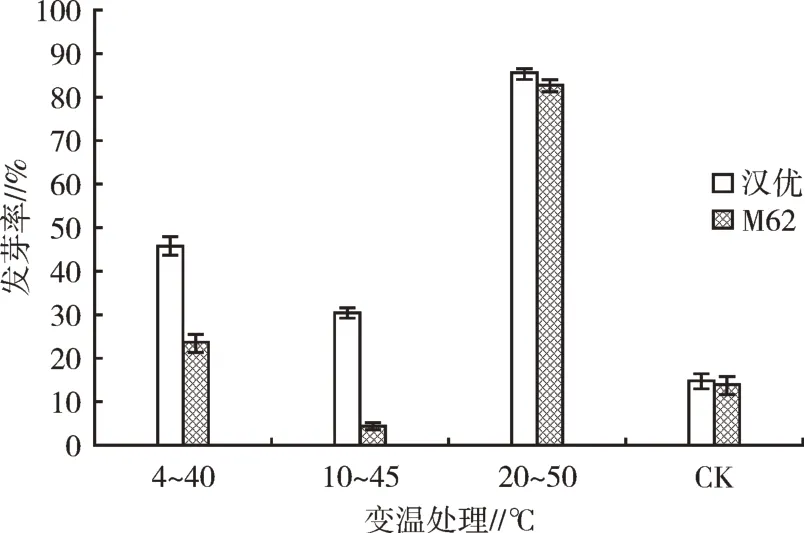
圖5 變溫處理對種子發芽的影響
3 小結與討論
試驗結果表明,通過不同的化學處理、物理處理以及激素處理可有效打破小白菜種子的休眠。其中KNO3浸種12、24 h均可使種子發芽率達到95%以上,且幼苗長勢旺盛;GA濃度低于1 000 mg/L時,浸種24 h可完全打破休眠;在引發處理中發現,400 mg/L GA和1% KNO3可完全打破休眠,發芽率達到100%;濃硫酸浸種可在不傷及胚的情況下有效破除種皮限制,可使M62種子發芽率達到100%,漢優發芽率達到95%以上,但幼苗后期生長不良,出芽后存在生長停止現象。50 mg/L 6-BA浸種處理也可有效打破M62和漢優種子的休眠。但是綜合對比發現KNO3與GA浸種打破休眠效果更優,且對幼苗生長無不良影響,因此可廣泛用于打破小白菜種子的休眠。
造成種子休眠的原因主要有生理性休眠(胚休眠)、存在抑制物和種皮限制,種子要想快速整齊地萌發也需要理想的內部條件和適宜的外部環境[12]。在調控種子休眠與萌發的過程中,植物激素是最重要的內部因素。1975年,Khan[13]提出了著名的三因子學說,其中“三因子”是指GA、CK和發芽抑制物(如ABA)。Khan[13]、屈海泳等[14]認為激素對休眠的調控是由于抑制物質可抵消促進細胞分裂和生長發育激素的作用,并且導致種子休眠的原因可能不僅是由于抑制物質的存在,赤霉素和細胞分裂素的缺乏也可能會導致種子休眠。大量研究結果表明,參與種子休眠與萌發過程調控的激素中脫落酸(ABA)和赤霉素(GA)是最主要的兩種調控激素,ABA參與誘導停止種胚細胞分裂、啟動并維持種子的休眠,而GA則起到啟動和促進種子萌發的作用,但GA主要在胚根突出時發生作用,且兩者在調節種子休眠和萌發過程中屬于相互拮抗的關系,它們的相對含量及敏感性在種子的休眠釋放和萌發過程中往往起到決定性作用[4,10,11,15-18]。目前赤霉素廣泛應用于打破植物種子休眠與調節植物衰老,但其具體作用機制還需要進一步研究。
KNO3促進種子萌發的作用機理較為復雜,對于KNO3打破休眠促進種子萌發的機制研究目前還比較缺乏。楊婷婷等[19]曾用KNO3處理芥菜,發現不同品種芥菜發芽率均有不同程度的提高。陳小奇等[20]用不同濃度KNO3分別浸泡稗草種子12、24 h,對稗草種子的萌發均有明顯促進作用。李敏等[11]的試驗表明,用1% KNO3溶液浸泡稻稗種子24 h解除休眠效果最好。對于KNO3促進種子萌發的機理,研究者們普遍認為硝酸鉀中的K+作為多種酶的激活劑,在一定程度上增強了有關酶的活性,從而提高了種子的生活力,促進種子發芽[19];K+作為營養元素可被種子吸收從而促進種子萌發[11];另外,KNO3具有強氧化性,作為一種強氧化劑,它主要通過強迫性供氧以促進代謝的進行,從而促進種子的萌發[21];但是對于KNO3打破休眠,促進種子萌發的具體機制還需要進一步的研究。
有生活力的種子發芽一般需要充足的氧氣、水分以及適宜的溫度,種子必須吸收充足的水分才能引發一系列酶的活性,從而促進種子的萌發。而一定濃度的強酸可以有效軟化或腐蝕種皮,以此增加種皮的透性,使種子吸收足夠的氧氣與水分;也可將種子內的還原性物質氧化變性、增強種子代謝,從而打破種子的休眠[11,22]。李敏等[11]用濃硫酸處理稻稗種子發現濃硫酸對種子萌發具有很好的促進作用;陳小奇等[20]發現在一定的時間范圍內,稗草種子的發芽率隨著濃硫酸處理時間的延長而提高;魏朝陽[23]用濃硫酸處理冬青種子發現濃硫酸酸蝕可促進種子萌發并且可以顯著提高發芽率,但種子腐爛嚴重;宋彬等[24]的研究也證實了濃硫酸處理可有效解除錦雞兒種子的休眠;多個研究均證明了濃硫酸可在不傷及種胚的情況下快速有效地打破種皮對發芽的限制,促進種子萌發。但必須控制其處理時間,處理時間過長會對種子造成損傷,時間過短則種皮軟化或腐蝕效果差,無法打破種子休眠。
