代謝工程改造大腸桿菌增產(chǎn)酪醇
2021-12-03 03:09:14曾嬌嬌余世琴周景文
食品與發(fā)酵工業(yè) 2021年22期
曾嬌嬌,余世琴,周景文*
1(江南大學(xué) 生物工程學(xué)院,工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無(wú)錫,214122) 2(未來(lái)食品科學(xué)中心(江南大學(xué)),江蘇 無(wú)錫,214122)
酪醇[2-(4-羥基苯基)乙醇,tyrosol]是一種天然的多酚類物質(zhì),廣泛存在于紅景天、女貞、橄欖等植物中,具有抗氧化、消炎等多種藥理活性[1]。目前酪醇的合成主要有植物提取、化學(xué)合成及微生物代謝生產(chǎn)3種途徑。由于植物提取率低、化學(xué)合成反應(yīng)復(fù)雜等問(wèn)題的存在,微生物細(xì)胞工廠的優(yōu)勢(shì)逐漸凸顯。在微生物代謝中,酪醇是酪氨酸代謝途徑的下游產(chǎn)物,是橄欖苦苷、羥基酪醇、紅景天苷等重要天然產(chǎn)物的關(guān)鍵前體。微生物中酪醇的合成途徑主要有3種:(1) SATOH等[2]在大腸桿菌中表達(dá)植物罌粟(Papaversomniferum)來(lái)源的酪氨酸脫羧酶(tyrosine decarboxylase,TDC)和藤黃微球菌(Micrococcusluteus)來(lái)源的酪胺氧化酶(tyramine oxidase,TYO),并結(jié)合大腸桿菌內(nèi)源的乙醇脫氫酶(alcohol dehydrogenase,ADH),實(shí)現(xiàn)了酪醇的從頭合成,產(chǎn)量達(dá)到69 mg/L;(2) YANG等[3]在大腸桿菌中過(guò)量表達(dá)來(lái)源于釀酒酵母的苯丙酮酸脫羧酶(phenylpyruvate decarboxylase,ARO10),得到了800.40 mg/L 酪醇;(3) CHUNG等[4]對(duì)擬南芥(Arabidopsisthaliana)、矮牽牛(Petuniahybrid)和荷蘭芹(Petroselinumcrispum)3種來(lái)源的芳香醛合成酶(aromatic aldehyde synthase,AAS)進(jìn)行了檢測(cè),發(fā)現(xiàn)表達(dá)PcAAS菌株的產(chǎn)量最高,達(dá)12.9 mg/L。在這3條途徑中,僅AAS可以將酪氨酸一步轉(zhuǎn)化為生產(chǎn)酪醇所需的直接前體物質(zhì)。因此,本研究將從此途徑入手開(kāi)展酪醇的生物合成研究。
ZHOU等[5-6]在大腸桿菌基因組中挖掘得到一系列梯度強(qiáng)度的組成型啟動(dòng)子,并用于強(qiáng)化己二酸的生產(chǎn)。為了提升酪醇的產(chǎn)量,本研究以E.coliBL21 (DE3)為出發(fā)菌株,使用組成型啟動(dòng)子PcspA[5]取代PT7啟動(dòng)子,并運(yùn)用該質(zhì)粒過(guò)量表達(dá)PcAAS來(lái)構(gòu)建酪醇合成的生物途徑。將調(diào)控分支代謝途徑基因敲除阻斷競(jìng)爭(zhēng)途徑和過(guò)量表達(dá)乙醇脫氫酶來(lái)強(qiáng)化代謝流的方法結(jié)合起來(lái),通過(guò)使用高、中、低3種不同強(qiáng)度的組成型啟動(dòng)子對(duì)途徑中的關(guān)鍵酶AAS和ADH進(jìn)行組合調(diào)控,實(shí)現(xiàn)了大腸桿菌合成酪醇的代謝途徑優(yōu)化。研究結(jié)果為后續(xù)采用發(fā)酵法生產(chǎn)酪醇和相關(guān)的下游產(chǎn)物奠定了良好的基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 菌株及質(zhì)粒
本研究使用及構(gòu)建的菌株和質(zhì)粒分別見(jiàn)表1和表2。
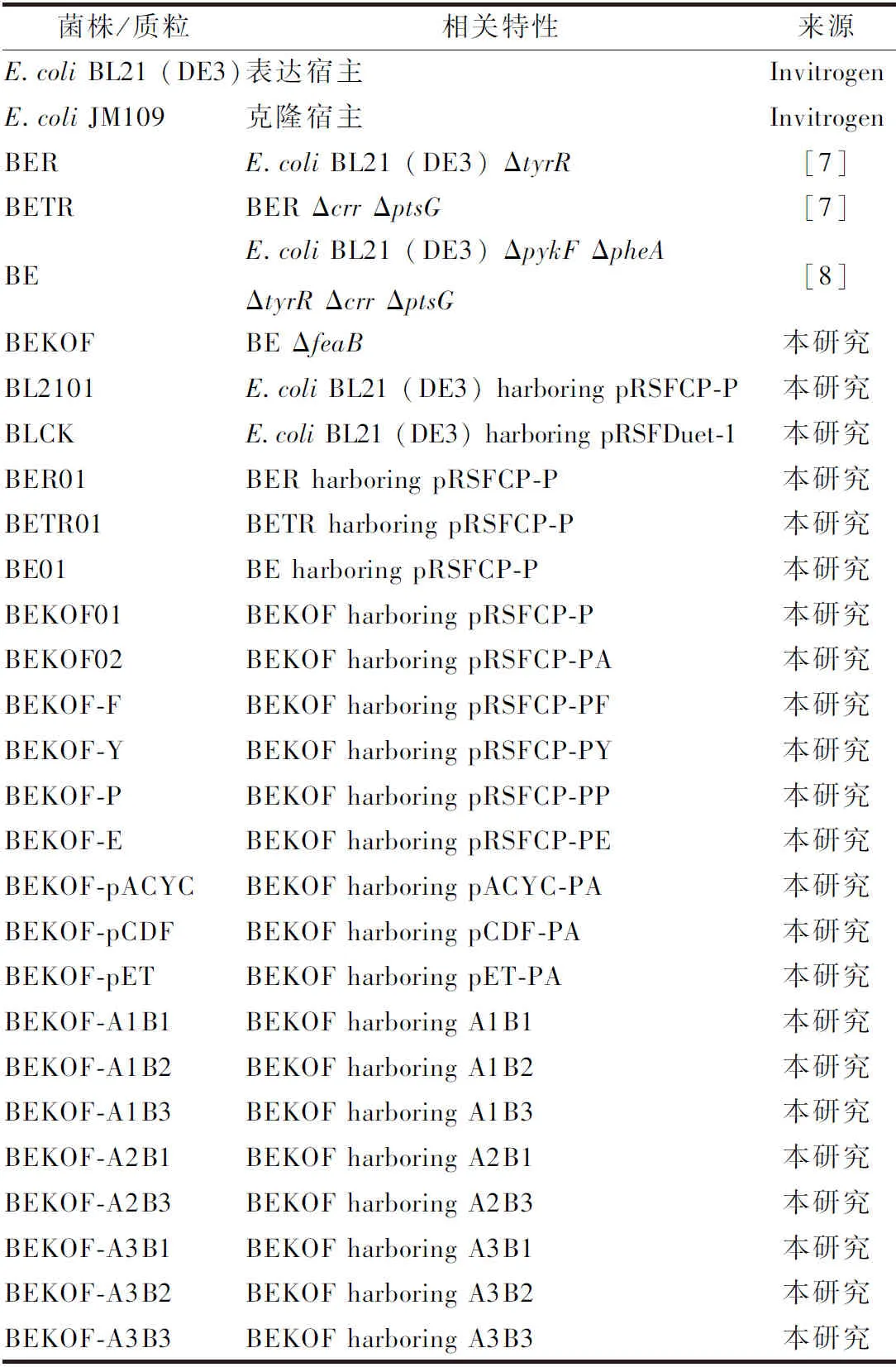
表1 本研究使用的菌株

表2 本研究使用的質(zhì)粒
1.1.2 培養(yǎng)基及試劑
種子培養(yǎng)基(g/L):胰蛋白胨10.0,酵母粉5.0,NaCl 10.0。
發(fā)酵培養(yǎng)基F1(g/L):葡萄糖 25.0,甘油 10.0,(NH4)2SO47.5,K2HPO4·3H2O 3.0,KH2PO42.0,MgSO4·7H2O 1.0,檸檬酸鈉 1.0,維生素B10.1,酵母粉 7.0,微量元素營(yíng)養(yǎng)液1 mL/L,依據(jù)需求加入適量的抗生素。
微量元素營(yíng)養(yǎng)液(g/L):Al2(SO4)3·18H2O 2.0,CoSO4·7H2O 0.75,CuSO4·5H2O 2.5,H3BO30.5,MnSO4·H2O 24.0,NiSO4·6H2O 2.5,ZnSO4·7H2O 15.0。
酵母粉,Oxoid公司;胰蛋白胨及所有使用的抗生素粉末,上海生工有限公司;酪醇標(biāo)準(zhǔn)品,阿拉丁;其余試劑購(gòu)自國(guó)藥集團(tuán)。
1.2 實(shí)驗(yàn)方法
1.2.1 基因組編輯方法
基因組編輯方法參考JIANG等[9]的研究。敲除feaB基因(phenylacetaldehyde dehydrogenase gene)所用的sgRNA序列為5′-CGATCTTTGATCCGGCCACC-3′,其PAM序列為GGG,所用相關(guān)引物如表3所示。

表3 基因敲除使用的引物
1.2.2 重組質(zhì)粒的構(gòu)建方法
質(zhì)粒構(gòu)建相關(guān)引物如表4所示。質(zhì)粒構(gòu)建采用Gibson組裝的方法進(jìn)行。首先通過(guò)簡(jiǎn)單PCR獲得目的片段,對(duì)得到的PCR產(chǎn)物純化后使用Gibson組裝酶進(jìn)行連接。將連接產(chǎn)物采用熱激法轉(zhuǎn)化E.coliJM109,次日挑取轉(zhuǎn)化子進(jìn)行菌落PCR并將得到的陽(yáng)性克隆進(jìn)行Sanger測(cè)序。
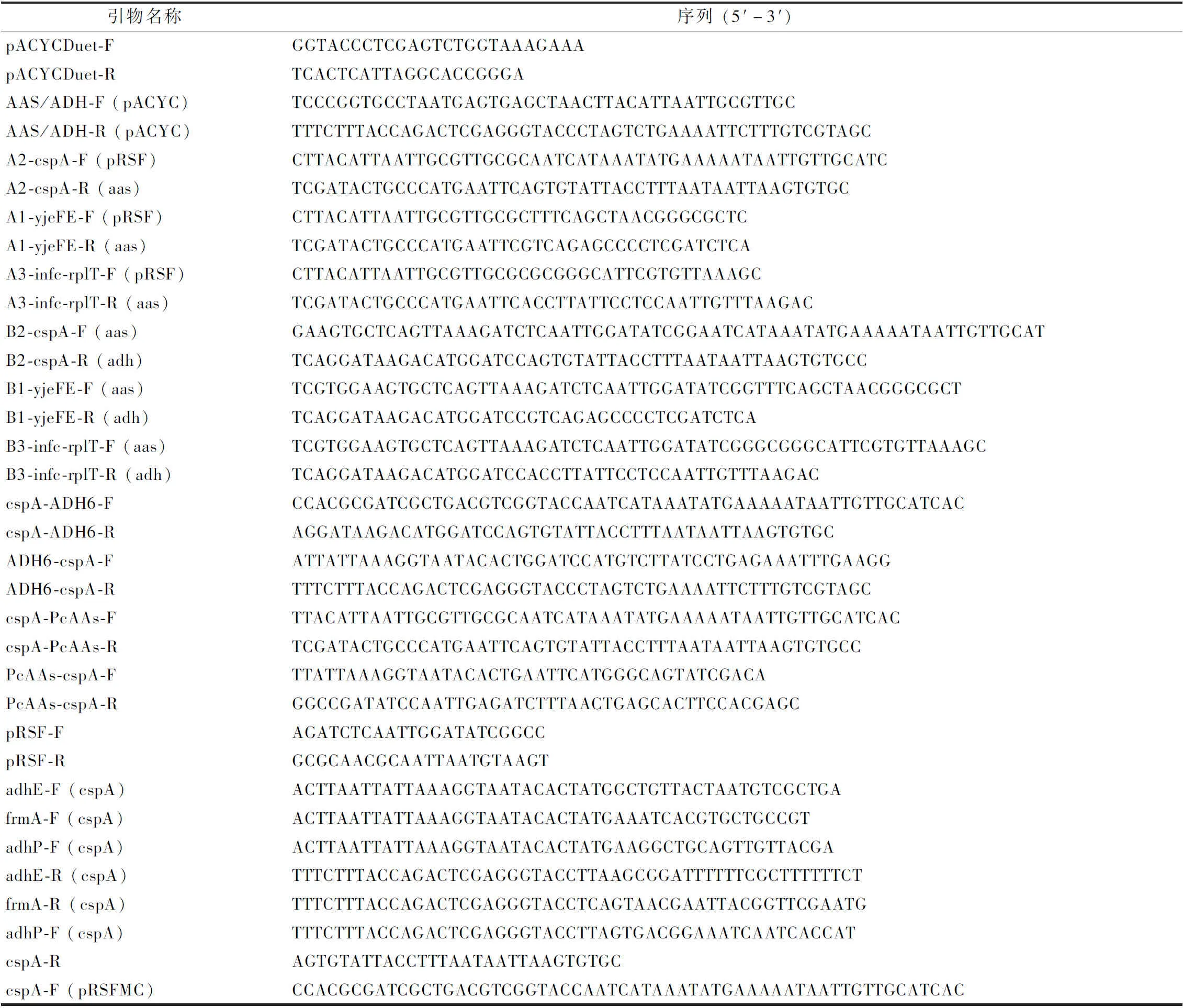
表4 質(zhì)粒構(gòu)建使用的引物
1.2.3 搖瓶發(fā)酵方法
在LB平板上劃線分離得到單菌落,將其接種于LB培養(yǎng)基中并加入適量抗生素,在37 ℃,220 r/min的搖床中過(guò)夜培養(yǎng)(10~14 h)。以3%的接種量轉(zhuǎn)接至發(fā)酵培養(yǎng)基F1中,將裝有20 mL培養(yǎng)基的250 mL 錐形瓶置于30 ℃,220 r/min的搖床中振蕩培養(yǎng),適時(shí)取樣。
1.2.4 樣品處理及HPLC檢測(cè)方法
由于酪氨酸微溶于水,利用酪氨酸的官能團(tuán)—NH2和H+可進(jìn)行化學(xué)反應(yīng)這一特性,用2 mol/L的HCl將發(fā)酵液樣品按體積比2∶1稀釋,以確保酪氨酸完全溶解。用去離子水將發(fā)酵液樣品進(jìn)一步稀釋,使最終產(chǎn)物濃度適宜分析方法的檢測(cè)范圍。稀釋完成后,12 000 r/min離心10 min。收集上清液并過(guò)濾處理得到用于HPLC測(cè)定的樣品。
HPLC測(cè)定的色譜柱為C18柱ODS-2 Hypersil(250 mm×4.6 mm,5 μm),紫外檢測(cè)器的檢測(cè)波長(zhǎng)為280 nm,A相為含有0.1%的三氟乙酸的去離子水,B相為含有0.1%三氟乙酸的乙腈,使用1.0 mL/min的流速進(jìn)行梯度洗脫,流速為1.0 mL/min,柱溫40 ℃, 進(jìn)樣量10 μL。酪氨酸和酪醇的出峰時(shí)間分別約為10.0、12.5 min。
2 結(jié)果與分析
2.1 酪醇合成途徑的構(gòu)建
本研究采用的代謝調(diào)控策略如圖1所示,選擇E.coliBL21 (DE3)作為出發(fā)菌株,利用化學(xué)轉(zhuǎn)化的方法將構(gòu)建的重組質(zhì)粒pRSFCP-P(攜帶有芳香醛合成酶PcAAS的基因)導(dǎo)入其中,得到菌株BL2101,以轉(zhuǎn)入空質(zhì)粒pRSFDuet-1的菌株作為對(duì)照菌株(菌株BLCK)。結(jié)果表明,24 h時(shí)菌株BL2101發(fā)酵液中有酪醇產(chǎn)生,但隨著發(fā)酵時(shí)間的延長(zhǎng),產(chǎn)量并未有明顯增加,菌株BL2101的最終產(chǎn)量為105.97 mg/L。液相-質(zhì)譜分析結(jié)果表明該產(chǎn)物為酪醇(圖2)。
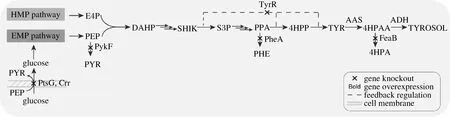
圖1 本研究相關(guān)代謝工程策略示意圖
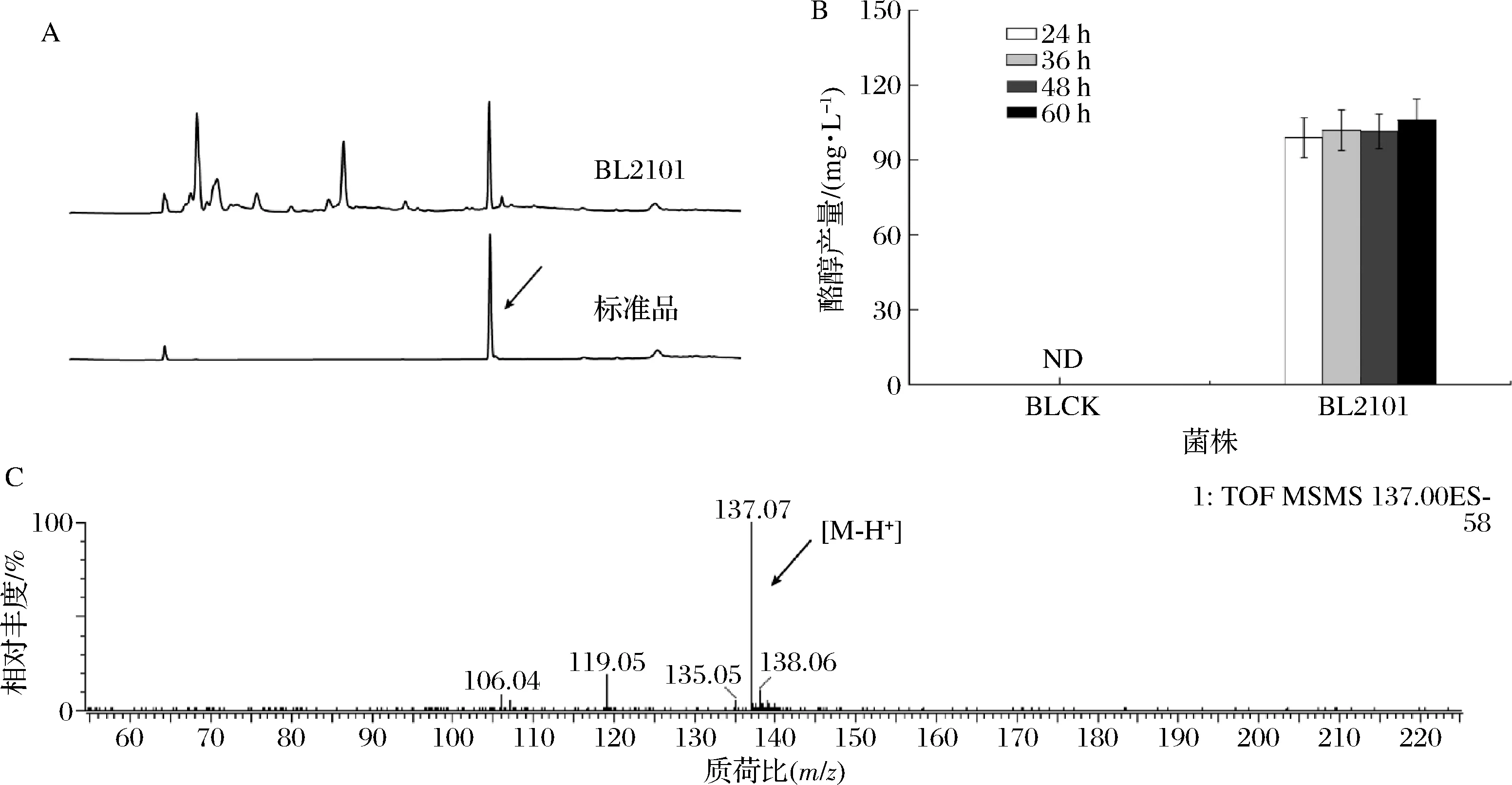
A-發(fā)酵24 h樣品及分析標(biāo)準(zhǔn)品液相檢測(cè)圖;B-從頭合成發(fā)酵產(chǎn)生的酪醇產(chǎn)量;C-發(fā)酵樣品質(zhì)譜圖
2.2 分支代謝途徑的阻遏
酪醇合成依賴的莽草酸途徑是苯丙氨酸、色氨酸、酪氨酸以及相關(guān)衍生物合成的重要代謝途徑。下調(diào)支路表達(dá)可以有效增強(qiáng)流向產(chǎn)物的代謝流。首先,在缺失反饋抑制基因(transcriptional regulatory protein gene,tyrR)的菌株BER中轉(zhuǎn)入質(zhì)粒pRSFCP-P獲得重組菌株BER01,酪醇產(chǎn)量為60.97 mg/L。在菌株BER的基礎(chǔ)上,繼續(xù)敲除在葡萄糖轉(zhuǎn)運(yùn)過(guò)程中會(huì)使磷酸烯醇式丙酮酸轉(zhuǎn)變?yōu)楸岬幕騪tsG和crr(均為葡萄糖轉(zhuǎn)運(yùn)過(guò)程相關(guān)基因),并通過(guò)質(zhì)粒過(guò)量表達(dá)PcAAS獲得的菌株BETR01,酪醇產(chǎn)量為76.24 mg/L。在此基礎(chǔ)上,敲除苯丙氨酸競(jìng)爭(zhēng)途徑的基因pheA(prephenate dehydratase gene)以及會(huì)使磷酸烯醇式丙酮酸流向丙酮酸的基因pykF(pyruvate kinase),并表達(dá)重組質(zhì)粒pRSFCP-P獲得菌株BE01,其酪醇產(chǎn)量達(dá)131.22 mg/L,相比其他2株重組菌株,酪醇產(chǎn)量分別提升了115.22%及72.11%。另外,通過(guò)敲除會(huì)使代謝流流向副產(chǎn)物對(duì)羥基苯乙酸的基因feaB,并在該菌株中過(guò)量表達(dá)PcAAS獲得菌株BEKOF01,酪醇產(chǎn)量達(dá)到1 055.36 mg/L(圖3)。相比菌株BE01,酪醇產(chǎn)量提升了704.2%;相比在E.coliBL21 (DE3)中過(guò)量表達(dá)PcAAS,酪醇產(chǎn)量提升了896.23%。
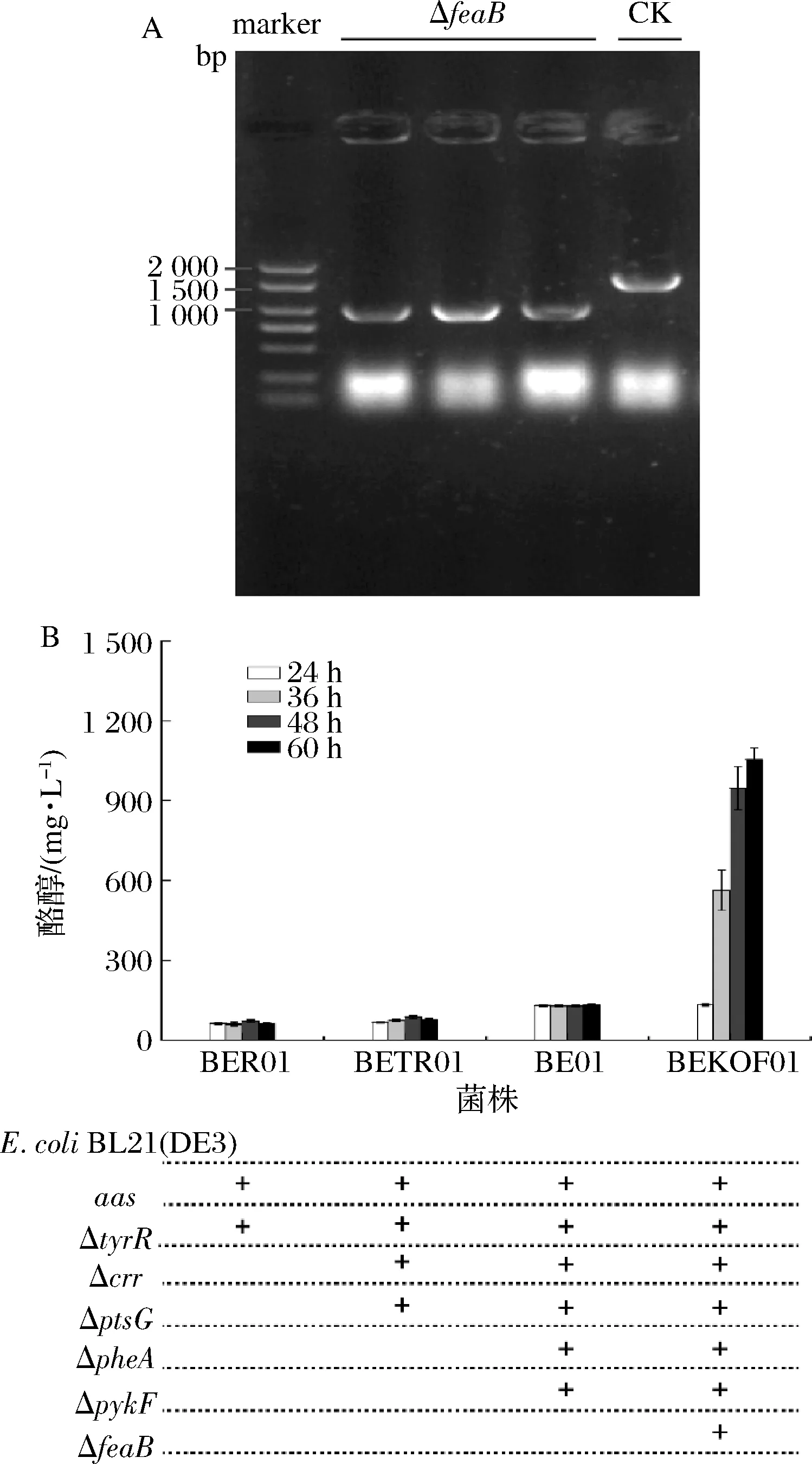
A-feaB敲除電泳圖;B-基因缺失對(duì)酪醇產(chǎn)量的影響
2.3 乙醇脫氫酶的篩選
對(duì)羥基苯乙醛的還原反應(yīng)是酪醇合成代謝途徑的最后一步,對(duì)酪醇生產(chǎn)具有重要影響。本研究利用大腸桿菌內(nèi)源的乙醇脫氫酶實(shí)現(xiàn)該步反應(yīng),酪醇產(chǎn)量達(dá)到1 055.36 mg/L。為了深入研究乙醇脫氫酶活性對(duì)酪醇積累的影響,本研究考察了4種來(lái)源于大腸桿菌的乙醇脫氫酶(AdhE[10]、AdhP[11]、YahK[12]及FrmA[13])及1種來(lái)源于釀酒酵母的乙醇脫氫酶(Adh6)[14-15](圖4)。結(jié)果表明,過(guò)量表達(dá)大腸桿菌來(lái)源的乙醇脫氫酶效果不佳;過(guò)量表達(dá)FrmA僅獲得516.92 mg/L的酪醇;過(guò)量表達(dá)YahK僅產(chǎn)生了96.33 mg/L的酪醇;過(guò)量表達(dá)AdhE的菌株發(fā)酵液及過(guò)量表達(dá)AdhP的菌株發(fā)酵液并未檢測(cè)到酪醇;過(guò)量表達(dá)釀酒酵母來(lái)源的Adh6效果顯著(菌株BEKOF02),酪醇產(chǎn)量達(dá)1 516.86 mg/L。 相比于未表達(dá)Adh6的菌株(菌株BEKOF01),酪醇產(chǎn)量提升了43.73%。
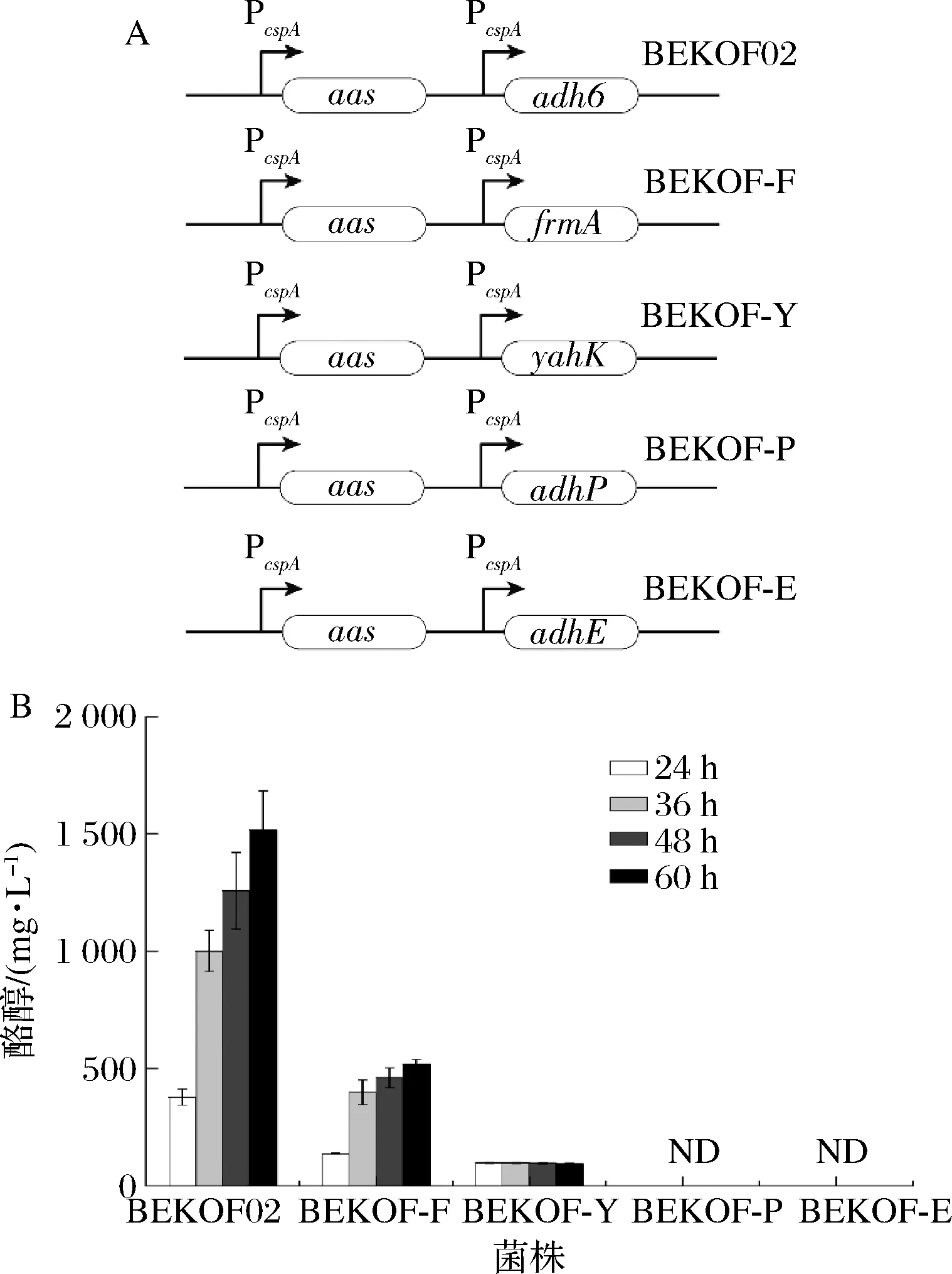
A-乙醇脫氫酶篩選;B-酪醇產(chǎn)量
2.4 關(guān)鍵酶PcAAS和ScADH6的組合調(diào)控
酶表達(dá)量的調(diào)節(jié)是代謝工程改造過(guò)程中的重要方法。在本研究中,通過(guò)調(diào)整質(zhì)粒拷貝數(shù)的方法,嘗試平衡酪醇生產(chǎn)的代謝流。質(zhì)粒pRSFDuet-1、pETDuet-1、pACYCDuet-1和pCDFDuet-1含有的復(fù)制子分別是RSF、pBR322、p15A和CDF,其拷貝數(shù)分別為100、40、20和10[16]。由于本研究已使用pRSFDuet-1作為載體,因此將PcAAS及ScADH6的表達(dá)框克隆至其余3個(gè)質(zhì)粒中,獲得菌株BEKOF-pET、BEKOF-pACYC及BEKOF-pCDF。3株菌株產(chǎn)量均低于50.00 mg/L (圖5、表5)。
因此,為了進(jìn)一步優(yōu)化代謝流,選擇具有不同表達(dá)強(qiáng)度的啟動(dòng)子PyjeFE(低強(qiáng)度)、PcspA(中強(qiáng)度)、Pinfc-rplT(高強(qiáng)度)[5]分別用于調(diào)節(jié)PcAAS及ScADH6的表達(dá)強(qiáng)度。結(jié)果表明,A2B1(使用啟動(dòng)子PcspA表達(dá)PcAAS,啟動(dòng)子PyjeFE表達(dá)ADH6)的組合產(chǎn)量較BEKOF02(使用啟動(dòng)子PcspA表達(dá)PcAAS,啟動(dòng)子PcspA表達(dá)ADH6)產(chǎn)量提升了293.60 mg/L,酪醇產(chǎn)量達(dá)到1 810.46 mg/L(圖5、表5)。
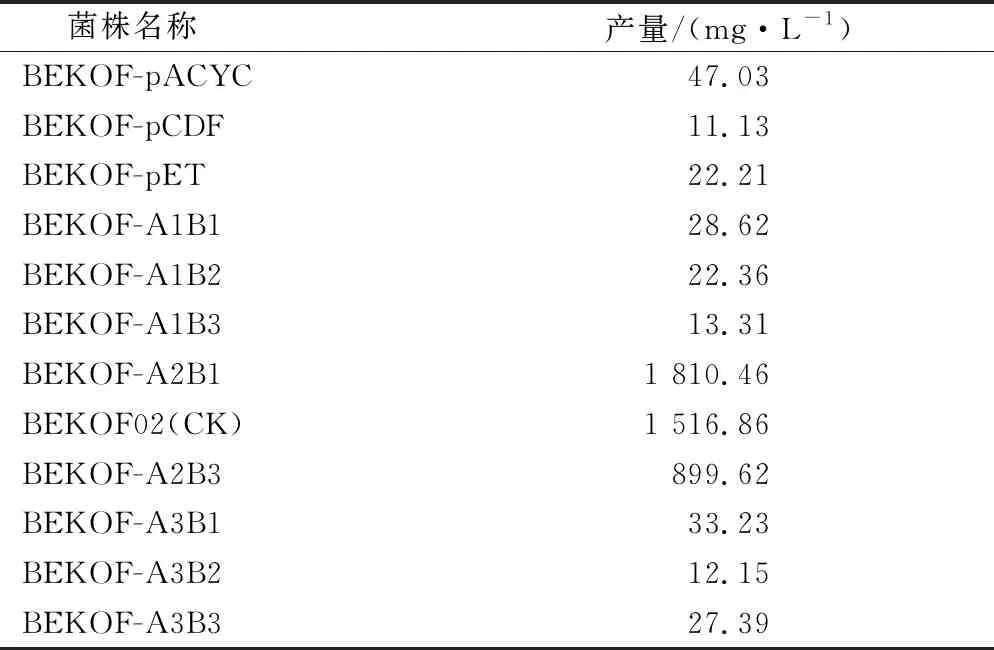
表5 復(fù)制子或啟動(dòng)子對(duì)酪醇產(chǎn)量的影響
A-酪醇產(chǎn)量;B-復(fù)制子篩選;C-啟動(dòng)子組合表達(dá)篩選
2.5 最佳菌株BEKOF-A2B1的發(fā)酵優(yōu)化
葡萄糖和甘油是大腸桿菌培養(yǎng)過(guò)程中常用的碳源,在細(xì)胞生長(zhǎng)過(guò)程中發(fā)揮著重要作用。此外,葡萄糖和甘油也是細(xì)胞糖酵解途徑的底物,其含量的高低直接決定了代謝所產(chǎn)生的產(chǎn)物含量。本研究中的出發(fā)培養(yǎng)基中含有葡萄糖25 g/L及甘油10 g/L,以此為基礎(chǔ),進(jìn)行了葡萄糖和甘油濃度的調(diào)整。圖6表明了不同配比下發(fā)酵60 h時(shí)酪醇的產(chǎn)量、菌體OD600以及發(fā)酵液pH。25 g/L葡萄糖和20 g/L甘油的組合具有優(yōu)勢(shì)(圖6-A),但發(fā)酵液pH較低,導(dǎo)致大腸桿菌難以進(jìn)行正常的代謝活動(dòng)。因此,通過(guò)提高培養(yǎng)基中氮源濃度的方法或外源添加來(lái)調(diào)節(jié)發(fā)酵過(guò)程中的pH,結(jié)果表明外源添加5 g/L碳酸鈣效果最佳,酪醇產(chǎn)量達(dá)到2 120.58 mg/L。
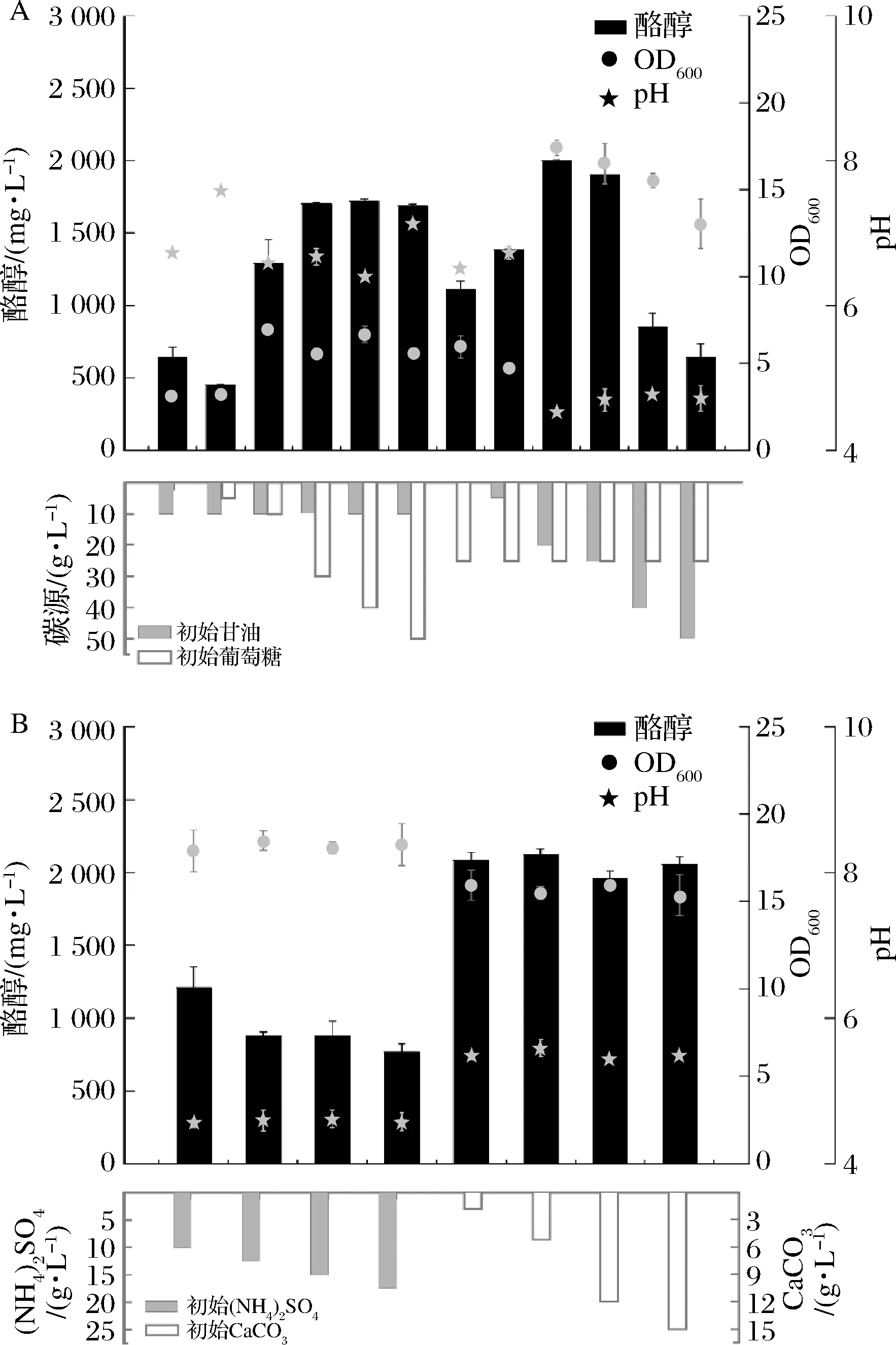
A-碳源濃度優(yōu)化;B-氮源濃度優(yōu)化及CaCO3濃度優(yōu)化
3 討論
酪醇因具有顯著的抗氧化、抗腫瘤等功能藥效成為近年來(lái)的研究熱點(diǎn)[17]。釀酒酵母由于具有完整的Ehrlich途徑,成為研究者們較為青睞的表達(dá)宿主。但是,由于較長(zhǎng)的發(fā)酵周期使得釀酒酵母作為生產(chǎn)菌株具有了一定的局限性,如LIU等[18]改造的對(duì)香豆酸生產(chǎn)菌株1個(gè)發(fā)酵周期需要96 h,GUO等[19]改造獲得的具有酪醇生產(chǎn)能力的釀酒酵母的發(fā)酵周期為196 h。大腸桿菌因發(fā)酵周期較短且易于基因工程改造等優(yōu)點(diǎn)成為了另一種選擇。研究表明,在宿主細(xì)胞中表達(dá)來(lái)源荷蘭芹的芳香醛合成酶可以構(gòu)建酪醇生物合成途徑[4,19-20]。本研究將經(jīng)密碼子優(yōu)化的PcAAS在E.coliBL21 (DE3)中過(guò)量表達(dá),酪醇產(chǎn)量達(dá)到105.97 mg/L。此外,在缺失了不同分支代謝途徑調(diào)控基因的菌株中過(guò)量表達(dá)PcAAS,以及敲除對(duì)酪醇積累具有顯著抑制作用的feaB基因,酪醇積累量達(dá)到1 055.36 mg/L,而未缺失feaB基因的菌株產(chǎn)量?jī)H為131.22 mg/L,研究結(jié)果與此前的報(bào)道類似[3, 13, 21]。
在多數(shù)研究中,醛基到羥基的還原反應(yīng)依賴于大腸桿菌中天然存在的醇脫氫酶[4, 13]。過(guò)量表達(dá)乙醇脫氫酶可以強(qiáng)化酪醇合成途徑的代謝流。GUO等[20]在釀酒酵母中表達(dá)來(lái)自大腸桿菌的乙醇脫氫酶,提高了酪醇的產(chǎn)量。本研究過(guò)量表達(dá)ScADH6使得酪醇的產(chǎn)量由1 055.36 mg/L提高至1 516.86 mg/L。另外,LIU等[18]通過(guò)同時(shí)優(yōu)化生物合成途徑中關(guān)鍵酶的啟動(dòng)子,顯著提高了對(duì)香豆酸的產(chǎn)量。在本研究使用中等強(qiáng)度啟動(dòng)子啟動(dòng)PcAAS與低強(qiáng)度啟動(dòng)子啟動(dòng)ScADH6的組合表現(xiàn)出較大潛力,酪醇產(chǎn)量達(dá)1 810.46 mg/L。 在此基礎(chǔ)上,通過(guò)優(yōu)化碳氮源初始濃度以及加入CaCO3改善發(fā)酵過(guò)程pH穩(wěn)定性,酪醇產(chǎn)量提高至2120.58 mg/L。在搖瓶發(fā)酵水平上,較XU等[13]研究中大腸桿菌YMG5A*R的產(chǎn)量1 506.96 mg/L及LIU等[22]研究中釀酒酵母LYTY16的產(chǎn)量702.30 mg/L,酪醇分別提升了40.72%和201.95%。但是,在發(fā)酵罐水平上,對(duì)比XU等[13]研究中的3.90 g/L和LIU等[22]研究中的9.90 g/L,仍存在一定差距。因此,在后續(xù)研究中,可以結(jié)合對(duì)前體供給途徑的系統(tǒng)改造和發(fā)酵過(guò)程優(yōu)化,使其成為羥基酪醇等高價(jià)值化合物合成的良好平臺(tái)菌株。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
科技傳播(2019年22期)2020-01-14 03:06:54
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36