鏈霉菌和殼聚糖對(duì)淮山土壤微生物活性與群落的影響
2021-12-08 07:36:30陸建明,黃小龍,吳文嬙,許云,夏薇,張榮萍
熱帶作物學(xué)報(bào) 2021年10期
關(guān)鍵詞:殼聚糖
陸建明,黃小龍,吳文嬙,許云,夏薇,張榮萍

摘? 要:為探究鏈霉菌30702和殼聚糖對(duì)淮山土壤微生物活性與群落的影響,采用比色法和滴定法觀測(cè)土壤酶活性的變化,采用高通量測(cè)序技術(shù)和Trimmomatic等軟件,分析土壤微生物群落中細(xì)菌和真菌的發(fā)展變化。結(jié)果表明,鏈霉菌、殼聚糖、土壤原始微生物和培養(yǎng)時(shí)間等4個(gè)因素對(duì)淮山土壤的脲酶和過氧化氫酶活性均有顯著影響,殼聚糖濃度在0~10.0 g/kg內(nèi),土壤酶活性隨殼聚糖濃度的增大而提高;隨時(shí)間的延長,先升再降;殼聚糖與鏈霉菌之間存在交互效應(yīng)。鏈霉菌和殼聚糖能增加淮山土壤細(xì)菌的物種豐度,減少細(xì)菌的多樣性,增加艾德昂菌屬(Ideonella)和纖維弧菌(Cellvibrio)等有益菌屬的相對(duì)豐度,而對(duì)真菌的物種豐度和多樣性的影響較小,與對(duì)照之間的差異不顯著,土壤優(yōu)勢(shì)菌屬為腐質(zhì)霉屬(Humicola)、枝孢屬(Cladosporium)、鞘氨醇單胞菌屬(Sphingomonas)和uncultured_bacterium_ c_Subgroup_6。其中,鏈霉菌與2.5 g/kg殼聚糖復(fù)合處理的細(xì)菌物種豐度最大,細(xì)菌多樣性最少。
關(guān)鍵詞:鏈霉菌;殼聚糖;淮山;土壤微生物活性;微生物群落;交互效應(yīng)
中圖分類號(hào):S156? ? ? 文獻(xiàn)標(biāo)識(shí)碼:A
Effect of Streptomyces and Chitosan on the Soil Microbial Activities and Microbiome in Dioscorea oppositifolia L.
LU Jianming, HUANG Xiaolong, WU Wenqiang, XU Yun, XIA Wei, ZHANG Rongping*
Hainan University, Haikou, Hainan 570228, China
Abstract: In order to explore the effect of Streptomyces 30702 and chitosan on the soil microbial activity and microbiome in Dioscorea oppositifolia L., the soil enzyme activity development was measured by colorimetric and titration methods, the change of the bacteria and fungi in soil microbiome was detected using the high-throughput sequencing technology and softwares such as Trimmomatic. The results showed that Streptomyces, chitosan, soil original microorganisms and culture time all affected the activity of urease and catalase in the soil significantly. The soil enzyme activity increased as chitosan concentration rose from 0 to 10.0 g/kg. As growing time went on, the soil enzyme activity increased first and then decreased obviously. Chitosan and Streptomyces interacted obviously. Streptomyces and chitosan increased the species abundance of soil bacteria and reduced the diversity of bacteria, increased the relative abundance of beneficial bacteria such as Ideonella and Cellvibrio, but had little effect on the species abundance and diversity of fungi and there was no significant difference between them and CK, the dominant bacteria genera in the soil were Humicola, Cladosporium, Sphingomonas and uncultured_bacterium_c_subgroup_6. Among them, the species abundance of bacteria in the Streptomyces 30702 2.5 g/kg treatment combined with chitosan was the largest, and the diversity of bacteria in the treatment was the least.
Keywords: Streptomyces sp.; chitosan; Dioscorea oppositifolia L.; soil microbial activity; soil microbiome; interaction effect
DOI: 10.3969/j.issn.1000-2561.2021.10.038
淮山(Dioscorea oppositifolia L.),又名薯蕷,為薯蕷科薯蕷屬作物,單子葉纏繞性藤本植物[1],多產(chǎn)于河南、山西、廣東、海南等地。淮山塊莖富含淀粉和各種營養(yǎng)成分以及Fe、Mn、Zn、Cu等豐富的微量元素及18種氨基酸,具有較高的食用和藥用價(jià)值[2]。近年來隨著各地政府的大力支持,我國淮山栽培面積越來越大,但淮山是忌連作作物,連作后產(chǎn)量下降、品質(zhì)降低、病蟲害加重[3],連作第2年,一般減產(chǎn)20%~30%,重者甚至枯死[4],制約了淮山產(chǎn)業(yè)的健康持續(xù)發(fā)展。研究表明,大豆和雙孢蘑菇的連作促使土壤微生物區(qū)系從高肥的“細(xì)菌型”土壤向低肥的“真菌型”土壤轉(zhuǎn)化,且土壤微生物總數(shù)減少[5-6],重茬根際土壤真菌的優(yōu)勢(shì)種群對(duì)作物根系的生長發(fā)育和養(yǎng)分吸收起不良作用,這是重茬作物生長發(fā)育障礙的重要原因。
鏈霉菌屬(Streptomyces)是產(chǎn)生抗生素最多的放線菌菌屬,鏈霉菌30702為課題組從藥用植物海南粗榧(Cephalotaxus hainanensis)根際分離得到[7],屬于紫黑鏈霉菌(Streptomyces viol-aceusniger)。該分枝的放線菌普遍可產(chǎn)生豐富的活性物質(zhì),已大量應(yīng)用于農(nóng)用抗生素等方面。王玉[8]研究發(fā)現(xiàn),在增加菌劑的番茄結(jié)果后期,土壤真菌較對(duì)照減少,而細(xì)菌數(shù)量增多,與連作0 a土壤微生物結(jié)構(gòu)相似;馬云艷等[9]研究發(fā)現(xiàn)生防放線菌改變了土壤中優(yōu)勢(shì)微生物的數(shù)量和比例,但是在實(shí)驗(yàn)室有明顯抑菌作用的生防菌盆栽和田間處理時(shí),受生防菌本身、病原菌和環(huán)境因子(如土壤中營養(yǎng)源、溫度、滲透性、pH)等影響較大,從而導(dǎo)致生防菌在土壤中的定殖能力和防病效果不穩(wěn)定[10]。經(jīng)試驗(yàn)證明,鏈霉菌30702不僅產(chǎn)生抗真菌作用的抗生素,還產(chǎn)生抑制病菌生長發(fā)育的蛋白酶、幾丁質(zhì)酶、纖維素酶和β-葡聚糖苷酶等水解酶[11],在防治山藥炭疽病方面也表現(xiàn)出良好的效果,有廣譜的抗菌效果[12-13],為本研究抑制土傳病害、改善土壤微生物群落結(jié)構(gòu)奠定了基礎(chǔ)。
殼聚糖(chitosan, CTS)是一種高分子量的陽性多糖,為線性β-(1→4)-N-乙酰-D-葡萄糖胺聚合物[14],是由幾丁質(zhì)脫乙酰所得。在農(nóng)業(yè)中,殼聚糖可增加土壤中有益的微生物放線菌[15],減少鐮刀菌、立枯絲核菌等病原菌,并可抑制根結(jié)線蟲,增加作物產(chǎn)量,提高品質(zhì)等[16],減少蔬菜土傳病害的發(fā)生[17]。而殼聚糖和微生物制劑對(duì)淮山土壤的調(diào)節(jié)研究較少報(bào)道,本試驗(yàn)采用光照培養(yǎng)箱的離體栽培方式,研究殼聚糖與鏈霉菌30702不同施用量處理下,淮山土壤微生物活性和微生物群落結(jié)構(gòu)的發(fā)展變化,為殼聚糖與鏈霉菌30702對(duì)淮山病害及其土壤的調(diào)控作用與機(jī)理研究奠定基礎(chǔ)。
1? 材料與方法
1.1? 材料
供試土壤為海南大學(xué)基地連作淮山5 a以上土壤(磚紅壤土,pH 5.38,有機(jī)質(zhì)3.4%,全氮0.12%),采用五點(diǎn)取樣法,采集距表層7~10 cm的連作淮山土壤,粉碎過10目(2 mm)篩,風(fēng)干,將150 g連作土裝入500 mL錐形瓶中,共30瓶,采用蒸汽蒸壓法,在121 ℃下連續(xù)3 d分3次對(duì)風(fēng)干土壤進(jìn)行蒸壓,獲得無菌土壤,待用。
殼聚糖(CTS,滬試,脫乙酰度80.0%~95.0%)、鏈霉菌30702(LM,薯蕷課題組提供)、ISP2培養(yǎng)基(酵母浸粉4 g、麥芽糖10 g、葡萄糖4 g、瓊脂20 g、蒸餾水1 L、pH 7.3)。將菌株30702接種在ISP2培養(yǎng)基中,28 ℃培養(yǎng)箱中培養(yǎng)14 d。將培養(yǎng)好的菌株接種于裝有150 mL無菌水的錐形瓶(500 mL)中,采用平板稀釋計(jì)數(shù)法制備濃度為4.2×106 CFU/mL的紫黑鏈霉菌30702菌懸液,待用。
1.2? 試驗(yàn)設(shè)計(jì)
試驗(yàn)土壤分成滅菌和不滅菌2組,殼聚糖濃度設(shè)為0、0.25、1.00、2.50、10.00 g/kg(干土)5個(gè)梯度,鏈霉菌濃度設(shè)0和1.0×106 CFU/g(干土)2個(gè)梯度,以清水處理為對(duì)照(CK),完全設(shè)計(jì)共20個(gè)處理,每個(gè)處理3個(gè)重復(fù)共60瓶(表1)。采用稱重法控制土壤含水量為20%,處理后置于智能光照培養(yǎng)箱中于30 ℃下16 h和20 ℃下8 h孵育[18]。
1.3? 土壤微生物群落結(jié)構(gòu)的測(cè)定
于處理后的第21天,取自然土試驗(yàn)組中清水(CK)、鏈霉菌30702(LM)和鏈霉菌與不同濃度殼聚糖復(fù)合處理(LM+CTS 0.25簡稱F1,LM+CTS 1簡稱F2,LM+CTS 2.5簡稱F3,LM+CTS 10簡稱F4)的15~20 g新鮮土樣,進(jìn)行高通量測(cè)序。高通量測(cè)序由北京百邁客生物科技有限公司代為完成。
1.3.1? 土壤總DNA提取? 使用MN NucleoSpin 96 Soil試劑盒/PowerSoil DNA Isolation kit強(qiáng)力土壤DNA提取試劑盒。
1.3.2? PCR擴(kuò)增? 采用AB9902 96 well PCR儀,對(duì)真菌ITS1區(qū)進(jìn)行PCR擴(kuò)增,引物為ITS1F(5-CTTGGTCATTTAGAGGAAGTAA-3)和ITS1R(5-GCTGCGTTCTTCATCGATGC-3);對(duì)細(xì)菌16S rRNA V3-V4區(qū)進(jìn)行PCR擴(kuò)增,引物序列采用341F(5-ACTCCTACGGGAGG?CA?GCA- 3)和806R(5-GGACTACHVGGGTWTCTAAT- 3)。PCR反應(yīng)體系:基因組DNA 50 ng ± 20%,*Vn F(10 μmol/L)0.3 μL,*Vn R(10 μmol/L)0.3 μL,KOD FX Neo Buffer 5 μL,dNTP(2 mmol/L each)2 μL,KOD FX Neo 0.2 μL,用ddH2O補(bǔ)至10 μL。PCR反應(yīng)條件:95 ℃ 5 min;95 ℃ 30 s;50 ℃ 30 s;72 ℃ 40 s;72 ℃ 7 min,20個(gè)循環(huán);72 ℃最后延伸5 min。Soleax PCR反應(yīng)體系:目的區(qū)域PCR純化產(chǎn)物2 μL,濃度為2 μmol/L的★MPPI-a 2.5 μL,濃度為2 μmol/L的★MPPI-b 2.5 μL,2×Q5 HF MM 10 μL(★根據(jù)上機(jī)安排進(jìn)行選擇index)。Soleax PCR反應(yīng)條件:98 ℃ 30 s;98 ℃ 10 s;65 ℃ 30 s;72 ℃ 30 s,20個(gè)循環(huán);72 ℃最后延伸5 min,1.8%的瓊脂糖凝膠,電壓120 V,40 min。
1.3.3? PCR產(chǎn)物的混樣和純化? 將Soleax PCR產(chǎn)物根據(jù)電泳定量(ImageJ軟件)結(jié)果,按照質(zhì)量比1∶1進(jìn)行混樣。混樣后,采用OMEGA DNA純化柱進(jìn)行過柱純化。電泳后,切目的片段,并回收。
1.3.4? 上機(jī)測(cè)序? 土壤DNA提取、PCR擴(kuò)增及測(cè)序均由北京百邁客生物科技有限公司協(xié)助完成。
1.4? 土壤酶活性的測(cè)定
于處理后的第7天、21天和140天取30 g土,風(fēng)干,進(jìn)行土壤脲酶、過氧化物酶活性測(cè)定。脲酶使用比色法,過氧化氫酶使用高錳酸鉀滴定法進(jìn)行測(cè)定[19]。
1.5? 數(shù)據(jù)處理
采用Trimmomatic軟件對(duì)原始測(cè)序序列進(jìn)行過濾、使用FLASH進(jìn)行雙端拼接,得到優(yōu)化序列(Tags);將優(yōu)化序列進(jìn)行聚類,利用CD-HIT方法劃分OTU,并根據(jù)OTU的序列組成得到其物種分類;細(xì)菌和真菌分別運(yùn)用Silva和UNITE數(shù)據(jù)庫進(jìn)行比對(duì),對(duì)樣品在各個(gè)分類水平上進(jìn)行分類學(xué)分析,獲得各樣品在門、綱、目、科、屬、種分類學(xué)水平上的物種聚類熱圖;運(yùn)用QIIME軟件分析單個(gè)樣品內(nèi)部的物種多樣性,統(tǒng)計(jì)各樣品在97%相似度水平下的Ace指數(shù)、Chao1指數(shù)、Shannon指數(shù)及Simpson指數(shù);通過Beta多樣性分析比較不同樣品在物種多樣性方面(群落組成及結(jié)構(gòu))存在的差異。運(yùn)用R軟件基于Uni Frac距離算法獲得相應(yīng)距離下的PCoA圖(有分組信息);通過16S功能基因預(yù)測(cè)分析,對(duì)樣品進(jìn)行基因功能預(yù)測(cè)并計(jì)算功能基因豐度,高通量數(shù)據(jù)分析由北京百邁客生物科技有限公司協(xié)助完成。不同處理土壤酶活性使用SAS 9.4統(tǒng)計(jì)軟件進(jìn)行多因素方差分析,采用鄧肯氏新復(fù)極差法進(jìn)行顯著性檢驗(yàn)。
2? 結(jié)果與分析
2.1? 鏈霉菌與殼聚糖對(duì)淮山土壤酶活性的影響
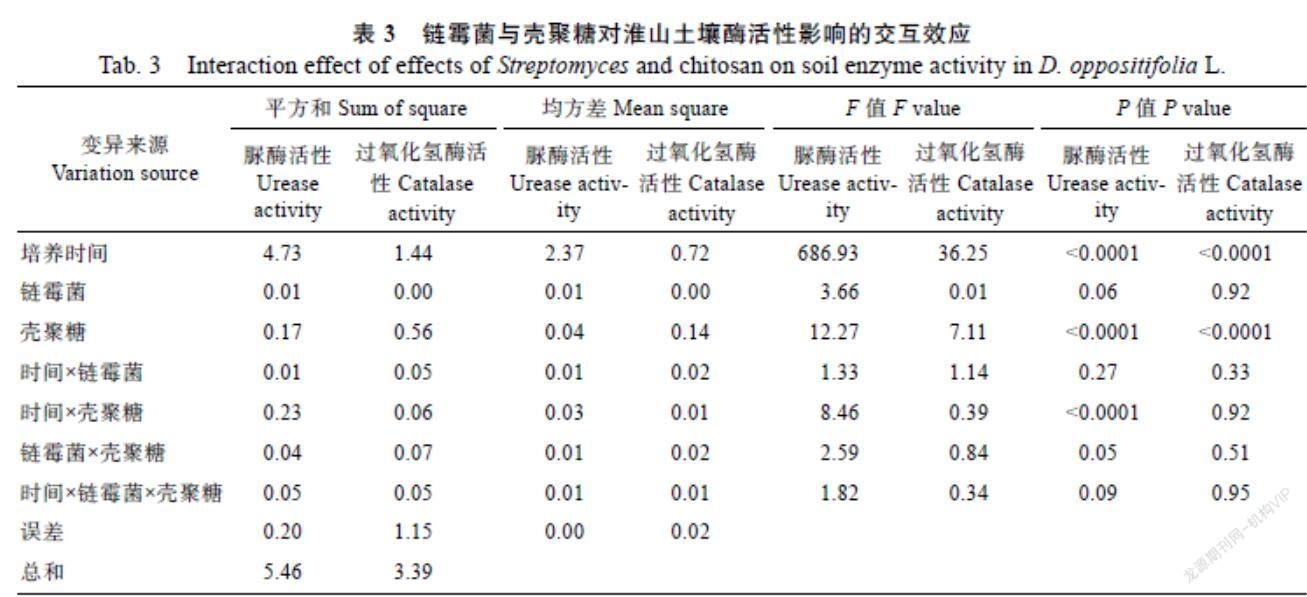
由表2、表3可以看出,土壤微生物、殼聚糖、鏈霉菌和培養(yǎng)時(shí)間均對(duì)土壤脲酶及過氧化氫酶活性有較大影響。其中,土壤微生物和培養(yǎng)時(shí)間對(duì)土壤酶活性的影響最大,淮山土壤中原始微生物和培養(yǎng)第21天時(shí)土壤酶活性最高,不同水平之間的差異極顯著;脲酶及過氧化氫酶活性均隨殼聚糖濃度的增大而升高;鏈霉菌對(duì)土壤脲酶活性有極顯著影響,而對(duì)過氧化氫酶活性的影響差異不顯著;殼聚糖與鏈霉菌之間有正交互效應(yīng),說明這2個(gè)因素之間存在互相促進(jìn)的關(guān)系,復(fù)合處理可增強(qiáng)生物酶活性的提升效果。
2.2? 殼聚糖與鏈霉菌對(duì)淮山土壤微生物群落的影響
2.2.1? 基于土壤ITS高通量測(cè)序的真菌多樣性分析? Alpha多樣性反映的是單個(gè)樣品物種豐度及
物種多樣性,有多種衡量指標(biāo):Chao1指數(shù)、Ace指數(shù)、Shannon指數(shù)、Simpson指數(shù)。Chao1指數(shù)和Ace指數(shù)可以衡量物種豐度即物種數(shù)量的多少,Shannon指數(shù)和Simpson指數(shù)用于衡量物種多樣性,受樣品群落中物種豐度和物種均勻度的影響。相同物種豐度的情況下,群落中各物種具有越大的均勻度,則認(rèn)為群落具有越大的多樣性,Shannon指數(shù)越大,Simpson指數(shù)越小,說明樣品的物種多樣性越高。由表4可知,鏈霉菌和殼聚糖對(duì)淮山土壤真菌的物種豐度和多樣性均無影響,與CK之間的差異均不顯著。
根據(jù)物種注釋結(jié)果,選取每組樣本在屬上最大豐度排名前10的物種,生成物種相對(duì)豐度柱形圖(圖1)。鏈霉菌和殼聚糖對(duì)土壤真菌的種類及其相對(duì)豐度均有一定影響。CK處理中,腐質(zhì)霉屬(Humicola)相對(duì)豐度較大,達(dá)21.55%,枝孢屬(Cladosporium)占11.4%,鐮刀菌屬(Fusarium)占2.48%,頭束霉屬(Cephalotrichum)占8.72%;鏈霉菌處理后與清水處理相比,腐質(zhì)霉屬下降了6.48%,而枝孢屬相對(duì)豐度上升了2.11%,頭束霉屬下降明顯,占1.3%,而曲霉菌屬(Aspergillus)有所增加;鏈霉菌與殼聚糖復(fù)合處理后,與鏈霉菌處理相比,枝孢屬相對(duì)豐度下降了4.46%~ 6.01%,鐮刀菌屬相對(duì)豐度上升0.94%~6.99%,均隨著殼聚糖濃度的增加而減少,而腐質(zhì)霉屬相對(duì)豐度隨殼聚糖濃度的增加而先增加后減少,其中鏈霉菌+1.00 g/kg殼聚糖處理中的腐質(zhì)霉屬相對(duì)豐度最高,鏈霉菌+0.25 g/kg殼聚糖處理的鏈格孢屬(Alternaria)最多,達(dá)8.55%。初步說明,鏈霉菌和殼聚糖處理改變了土壤真菌群落結(jié)構(gòu),但優(yōu)勢(shì)菌屬仍為腐質(zhì)霉屬(Humicola)、枝孢屬(Cladosporium)、Tausonia等。
CK為清水,LM為鏈霉菌菌懸液,F(xiàn)1為鏈霉菌+0.25 g/kg殼聚糖,F(xiàn)2為鏈霉菌+1.00 g/kg殼聚糖,F(xiàn)3為鏈霉菌+2.50 g/kg殼聚糖,F(xiàn)4為鏈霉菌+10.00 g/kg殼聚糖。
網(wǎng)絡(luò)圖是相關(guān)性分析的一種表現(xiàn)形式,可以直觀地體現(xiàn)各個(gè)分類水平(門、綱、目、科、屬、種)的物種在所選擇樣品之間含有的相關(guān)性關(guān)系。根據(jù)屬分類水平各個(gè)物種在各個(gè)樣品中的豐度以及變化情況進(jìn)行相關(guān)分析,計(jì)算物種間的相關(guān)性,并進(jìn)行統(tǒng)計(jì)檢驗(yàn),基于Python繪制共表達(dá)分析網(wǎng)絡(luò)圖(圖2)。根據(jù)圖2可得,枝孢屬與被孢霉屬(Mortierella)正相關(guān)且相關(guān)性最強(qiáng),Occultifur與角菌屬(Angulomyces)、Pyrenochaetopsis與梭孢殼屬(Thielavia)、輪枝孢屬(Verticillium)與Vishniacozyma正相關(guān)且相關(guān)性較強(qiáng);嗜熱真菌屬(Thermomyces)與淡紫紫孢菌(Purpureocillium)負(fù)相關(guān)且相關(guān)性最強(qiáng),元胞菌(Metacordyceps)與Cladorrhhinum、佐菲拉菌屬(Zopfiella)與絲孢菌屬(Scedosporium)、黑團(tuán)孢屬(Periconia)與莖點(diǎn)霉屬(Phoma)負(fù)相關(guān)且相關(guān)性較強(qiáng)。其中,腐質(zhì)霉屬豐度最高,其次為枝孢屬、Tausonia、頭束霉屬。
2.2.2? 基于土壤16S高通量測(cè)序的細(xì)菌多樣性分析? 根據(jù)Alpha多樣性分析結(jié)果(表5)可得,鏈霉菌和殼聚糖處理后,土壤中的細(xì)菌物種豐度大于清水處理,以F3處理最高。清水處理的土壤細(xì)菌多樣性大于鏈霉菌和殼聚糖處理,即鏈霉菌和殼聚糖處理使土壤細(xì)菌的物種多樣性減少,F(xiàn)1、F2、F3和F4均顯著少于CK和LM處理,而LM與CK之間差異不顯著。結(jié)果表明,鏈霉菌對(duì)土壤細(xì)菌的物種豐度和多樣性的影響不大,鏈霉菌與殼聚糖復(fù)合處理,可增加土壤細(xì)菌的物種豐度,而減少細(xì)菌的多樣性。
圖3為各處理以屬為分類級(jí)別的相對(duì)豐度圖,由圖3可以看出,鏈霉菌和殼聚糖對(duì)土壤的細(xì)菌及其相對(duì)豐度有一定影響。CK清水處理中,uncultured_bacterium_c_Subgroup_6相對(duì)豐度較高,達(dá)10.44%,鞘氨醇單胞菌屬(Sphingomonas)占8.72%;LM處理與CK相比,Jatropha_curcas相對(duì)豐度較清水處理有上升,占1.27%,其他與CK處理相似;鏈霉菌與殼聚糖復(fù)合處理后,uncultured_bacterium_c_Subgroup_6和鞘氨醇單胞菌屬(Sphingomonas)相對(duì)豐度分別下降1.46%~ 4.26%和2.36%~4.23%;艾德昂菌屬(Ideonella)的相對(duì)豐度整體上升3.55%~24.14%,隨著殼聚糖濃度的增加,先升高后下降,其中,F(xiàn)3最高,占24.49%;纖維弧菌屬(Cellvibrio)的相對(duì)豐度升高,其中,F(xiàn)2最高,占19.4%。Jatropha_curcas相對(duì)豐度在F1處理中最高,占15.68%。
由圖4可知,JGI_00010001-H03與別生根瘤菌-新生根瘤菌-寄生根瘤菌-根瘤菌(Allorhiz?ob-ium-Neorhizobium-Pararhizobium-Rhizobium)正相關(guān)且相關(guān)性最強(qiáng),uncultured_bac?te?rium_c_ Alphaproteobacteria與Pelomonas和Ellin6067、uncultured_bacterium_o_Elsterales與嗜紅桿菌(Altererythrobacter)、uncultured_bacterium_o_ IMCC26256與uncultured_bacterium_o_Rokuba-cteriales正相關(guān)且相關(guān)性較強(qiáng);Gaiella與鏈霉菌屬(Streptomyces)負(fù)相關(guān)且相關(guān)性最強(qiáng),Jatropha_curcas與uncultured_bacterium_o_Rok-uba?cteriales、uncultured_bacterium_c_Alph?aprot?eo-bacteria與uncultured_bacterium_c_Acidim?icro?biia、滋養(yǎng)層桿菌(Stenotrophobacter)與諾卡氏菌屬(Nocardioides)、硝化螺菌屬(Nitrospira)與uncultured_bacterium_o_Saccharimonadales負(fù)相關(guān)且相關(guān)性較強(qiáng)。其中,艾德昂菌屬(Ideonella)豐度最高,其次為纖維弧菌屬(Cellvibrio)、鞘氨醇單胞菌屬(Sphingomonas)和Jatropha_curcas。
3? 討論
本研究結(jié)果表明,鏈霉菌、殼聚糖、土壤原始微生物和培養(yǎng)時(shí)間等4個(gè)因素對(duì)淮山土壤的脲酶和過氧化氫酶活性均有顯著影響,殼聚糖濃度在0~10.0 g/kg范圍內(nèi),土壤酶活性隨殼聚糖濃度的增大而提高;隨培養(yǎng)時(shí)間的延長,土壤酶活性先升再降;殼聚糖與鏈霉菌之間存在交互效應(yīng)。鏈霉菌和殼聚糖處理增加淮山土壤細(xì)菌的物種豐度,而減少細(xì)菌的多樣性,增加了艾德昂菌屬(Ideonella)和纖維弧菌(Cellvibrio)等有益菌屬的相對(duì)豐度,而對(duì)真菌的物種豐度和多樣性的影響較小,土壤優(yōu)勢(shì)菌屬為腐質(zhì)霉屬(Humicola)、枝孢屬(Cladosporium)、鞘氨醇單胞菌屬(Sphingomonas)和uncultured_bacte?rium_c_Su-bgroup_6。其中,鏈霉菌與2.5 g/kg殼聚糖復(fù)合處理的細(xì)菌物種豐度最大,細(xì)菌的多樣性最少。
土壤酶是重要的土壤微生態(tài)環(huán)境指示因子,土壤脲酶可以水解土壤中尿素生成氨、二氧化碳和水[20],能部分反映土壤的生產(chǎn)力,土壤過氧化氫酶可以反映有機(jī)質(zhì)的轉(zhuǎn)化速度和土壤腐殖質(zhì)化的強(qiáng)度大小[21]。尹淑麗[22]研究發(fā)現(xiàn),單一生防菌株對(duì)土壤脲酶及過氧化氫酶活性有促進(jìn)作用,與本研究結(jié)果一致。王艷芳等[23]使用1.0 g/kg甲殼素提高了土壤脲酶及過氧化氫酶活性,分別提高了40.5%和18.7%,與本研究結(jié)果相似。
抑制植物土傳病害在一定程度上是土壤微生物的群體作用,土壤微生物群落結(jié)構(gòu)越豐富,物種越均勻,抑制病原菌的綜合能力增強(qiáng)[24]。研究表明,土壤微生物多樣性與土壤肥力、土傳病害密切相關(guān)[25]。在設(shè)施蔬菜種植過程中,土壤微生物群落結(jié)構(gòu)與土壤質(zhì)量同步變化,進(jìn)而反映土壤質(zhì)量受損程度[26],可以作為土壤質(zhì)量變化的重要預(yù)警指標(biāo)之一[27]。Tripathi等[28]研究表明,在脅迫條件下,腐質(zhì)霉屬(Humicola)對(duì)植物生長有促進(jìn)作用。枝孢屬(Cladosporium)是一種可用于抗植物源的真菌,對(duì)由種子傳播的病原引起的水稻病害有較高的防效,可作為生防菌劑、酶和生物活性肽的潛在來源已被廣泛研究[29]。鞘氨醇單胞菌屬(Sphingomonas)能增強(qiáng)擬南芥的抗旱性,通過釋放揮發(fā)性有機(jī)化合物促進(jìn)植物生長[30]。枝孢屬(Cladosporium)與被孢霉屬(Mortierella)正相關(guān)且相關(guān)性最強(qiáng),枝孢屬可引起牡丹葉斑病[31],同時(shí)也是番茄葉霉的病原菌。本研究發(fā)現(xiàn),鏈霉菌和殼聚糖處理后,淮山土壤中腐質(zhì)霉屬(Humicola)、枝孢屬(Cladosporium)、鞘氨醇單胞菌屬(Sphingomonas)和uncult?ured_bac?-terium_c_Subgroup_6均為優(yōu)勢(shì)菌屬,這些真菌或細(xì)菌可能在淮山根際土壤中發(fā)揮重要的作用。
本研究表明,鏈霉菌30702與殼聚糖復(fù)合處理與清水處理對(duì)比,土壤中鐮刀菌屬(Fusarium)和艾德昂菌屬(Ideonella)、纖維弧菌(Cellvibrio)相對(duì)豐度均有不同程度的增加。艾德昂菌屬中的一種細(xì)菌使用聚對(duì)苯二甲酸乙二醇酯(PET)作為能量源,能夠在30 ℃溫度條件下經(jīng)過6周完全降解PET薄膜[32];革蘭氏陰性腐生植物纖維弧菌對(duì)纖維素、木聚糖和果膠底物具有顯著降解能力,纖維弧菌作為纖維素降解細(xì)菌,具有強(qiáng)降解多糖的能力[33-34],也是廣東博羅普通野生水稻主要內(nèi)生固氮菌屬之一[35],因此纖維弧菌豐度的增高有助于更有效地分解有機(jī)物,使作物更有效地利用物質(zhì)和能量。推測(cè)鏈霉菌30702與殼聚糖的交互作用可通過豐富土壤微生物多樣性實(shí)現(xiàn)抑制病原菌的作用。
本研究發(fā)現(xiàn),鏈霉菌和殼聚糖處理后,淮山土壤的細(xì)菌物種豐度增加,多樣性減少,真菌物種豐度和多樣性影響較小,土壤微生物區(qū)系從低肥的“真菌型”土壤向高肥的“細(xì)菌型”土壤轉(zhuǎn)化。Phillips等[36]的研究結(jié)果表明,施用微生物菌劑后,增加了土壤微生物的多樣性。馬海濱等[37]、陳紅麗等[38]研究得出,殼聚糖可以豐富連作煙草土壤的微生物多樣性,與本研究結(jié)果相似。
4? 結(jié)論
研究表明,鏈霉菌30702和殼聚糖能促進(jìn)淮山土壤酶活性,增加土壤細(xì)菌的物種豐度,降低細(xì)菌的多樣性,對(duì)真菌的物種豐度和多樣性影響小,土壤優(yōu)勢(shì)菌屬為腐質(zhì)霉屬(Humicola)、枝孢屬(Cladosporium)、鞘氨醇單胞菌屬(Sphi?ngo-monas)和uncultured_bac?terium_c_Sub?g?roup_6,增加了艾德昂菌屬(Ideonella)和纖維弧菌(Cellvibrio)等有益菌屬的相對(duì)豐度。本研究結(jié)果為鏈霉菌和殼聚糖防治淮山病害和調(diào)節(jié)土壤的研究奠定了理論基礎(chǔ)。
參考文獻(xiàn)
[1] 湯? 潔, 戴興臨, 涂玉琴, 等. 淮山藥高效栽培技術(shù)研究[J]. 安徽農(nóng)業(yè)科學(xué), 2017, 45(4): 122-127.
[2] 李月仙, 黃東益, 黃小龍, 等. 山藥的研究進(jìn)展[J]. 中國農(nóng)學(xué)通報(bào), 2009, 25(9): 91-96.
[3] 岑家蘭, 王雪芳, 黎祖文, 等. 廣西八步區(qū)淮山生產(chǎn)連作障礙的影響及解決措施[J]. 長江蔬菜, 2016(7): 3-4.
[4] 陳偉益, 黃東益, 吳文嬙, 等. 殼聚糖及其衍生物對(duì)連作淮山炭疽病的影響[J]. 海南大學(xué)學(xué)報(bào)(自然科學(xué)版), 2020, 38(2): 141-146.
[5] 鄒? 莉, 袁曉穎, 李? 玲, 等. 連作對(duì)大豆根部土壤微生物的影響研究[J]. 微生物學(xué)雜志, 2005(2): 27-30.
[6] 黃? 敏. 大田雙孢蘑菇連作障礙的土壤微生物學(xué)特性研究[D]. 雅安: 四川農(nóng)業(yè)大學(xué), 2006.
[7] 黃小龍, 陳吉良, 李建平, 等. 熱帶藥用植物根際放線菌的分離、鑒定及生物活性分析[J]. 生物技術(shù)通報(bào), 2012(2): 121-127.
[8] 王? 玉. 放線菌劑對(duì)連作番茄生理生態(tài)及土壤微環(huán)境的影響[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2012.
[9] 馬云艷, 徐萬里, 唐光木, 等. 生防鏈霉菌配施棉稈炭對(duì)連作棉田土壤微生物區(qū)系的影響[J]. 中國生態(tài)農(nóng)業(yè)學(xué)報(bào), 2017, 25(3): 400-409.
[10] 張炳欣, 張? 平, 陳曉斌. 影響引入微生物根部定殖的因素[J]. 應(yīng)用生態(tài)學(xué)報(bào), 2000(6): 951-953.
[11] 叢子文, 焦敬華, 周雙清, 等. 鏈霉菌30702的鑒定及其生防特性[J]. 生物技術(shù)通報(bào), 2018, 34(6): 190-198.
[12] 焦敬華. 山藥炭疽病生防菌30702的生防特性及分類鑒定[D]. 海口: 海南大學(xué), 2016.
[13] 焦敬華, 黃東益, 吳文嬙, 等. 山藥炭疽病菌拮抗放線菌30702菌株的初步鑒定及發(fā)酵培養(yǎng)基優(yōu)化[J]. 熱帶作物學(xué)報(bào), 2016, 37(4): 775-783.
[14] Chien P J, Chou C C. Antifungal activity of chitosan and its application to control post-harvest quality and fungal rotting of Tankan citrus fruit (Citrus tankan Hayata)[J]. Journal of the Science of Food and Agriculture, 2006, 86: 1964-1969.
[15] 劉清瑋, 宋宇鵬. 有機(jī)肥對(duì)三年生農(nóng)田栽培人參根際微生態(tài)及產(chǎn)量的影響[J]. 吉林農(nóng)業(yè)大學(xué)學(xué)報(bào), 2020, 42(4): 409-414.
[16] Bol J F, Linthorst H, Cornelissen B. Plant pathogenesis-related proteins induced by virus infection[J]. Annual Review of Phytopathology, 1990, 28(1): 113-138.
[17] 李寶英. 殼聚糖制劑控制蔬菜土傳病害的研究[J]. 中國農(nóng)學(xué)通報(bào), 2005(1): 275-277.
[18] Yuan W M, Crawford D L. Characterization of streptomyces lydicus WYEC108 as a potential biocontrol agent against fungal root and seed rots[J]. Applied and Environmental Microbiology, 1995, 61(8): 3119-3128.
[19] 關(guān)松蔭. 土壤酶及其研究法[M]. 北京: 農(nóng)業(yè)出版社, 1986.
[20] 劉宇彤, 霍璐陽, 李志國, 等. 不同處理方式對(duì)土壤酶活性的影響[J]. 森林工程, 2019, 35(2): 21-26.
[21] 曾憲軍, 劉登魁, 朱世民, 等. 不同濃度阿特拉津?qū)θN肥力條件土壤過氧化氫酶的影響[J]. 湖南農(nóng)業(yè)科學(xué), 2005(6): 33-35.
[22] 尹淑麗, 麻耀華, 張麗萍, 等. 不同生防菌對(duì)黃瓜根際土壤微生物數(shù)量及土壤酶活性的影響[J]. 北方園藝, 2012(1): 10-14.
[23] 王艷芳, 付風(fēng)云, 李家家, 等. 甲殼素對(duì)連作條件下平邑甜茶幼苗生長及土壤環(huán)境的影響[J]. 生態(tài)學(xué)報(bào), 2016, 36(19): 6218-6225.
[24] 蔡燕飛, 廖宗文, 章家恩, 等. 生態(tài)有機(jī)肥對(duì)番茄青枯病及土壤微生物多樣性的影響[J]. 應(yīng)用生態(tài)學(xué)報(bào), 2003(3): 349-353.
[25] 章家恩, 廖宗文. 試論土壤的生態(tài)肥力及其培育[J]. 生態(tài)環(huán)境, 2004, 9(3): 253-256.
[26] 楊? 紅, 徐唱唱, 曹麗花, 等. 米林縣不同種植年限蔬菜大棚土壤pH和無機(jī)氮變化特征研究[J]. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào), 2016, 35(12): 2397-2404.
[27] Zhang J, Wang P C, Tian H M, et al. Pyrosequencing-based assessment of soil microbial community structure and analysis of soil properties with vegetable planted at different years under greenhouse conditions[J]. Soil and Tillage Research, 2019, 187: 1-10.
[28] Tripathi P, Khare P, Barnawal D, et al. Bioremediation of arsenic by soil methylating fungi: Role of Humicola sp. strain 2WS1 in amelioration of arsenic phytotoxicity in Bacopa monnieri L.[J]. Science of the Total Environment, 2020, 716: 136758.
[29] Ren J, Xue C, Tian L, et al. Asperelines A-F, peptaibols from the marine-derived fungus Trichoderma asperellum[J]. Journal of Natural Products, 2009, 72(6): 1036-1044.
[30] Luo Y, Zhou M, Zhao Q, et al. Complete genome sequence of Sphingomonas sp. Cra20, a drought resistant and plant growth promoting rhizobacteria[J]. Genomics, 2020, 112(5): 3648-3657.
[31] 段亞冰. 牡丹葉斑病病原真菌鑒定及生物學(xué)特性研究[D]. 洛陽: 河南科技大學(xué), 2009.
[32] Voets J P, Van Hove J K. Purification and cellulolytic activity of Cellvibrio[J]. Nature, 1953, 171(4363): 1073-1074.
[33] Gardner J G. Polysaccharide degradation systems of the saprophytic bacterium Cellvibrio japonicus[J]. World Journal of Microbiology and Biotechnology, 2016, 32(7): 1-12.
[34] Voets J P, Van Hove J K. Purification and cellulolytic activity of Cellvibrio.[J]. Nature, 1953, 171(4363): 1073-1074.
[35] 譚志遠(yuǎn), 彭桂香, 徐培智, 等. 普通野生稻(Oryza rufipogon)內(nèi)生固氮菌多樣性及高固氮酶活性[J]. 科學(xué)通報(bào), 2009, 54(13): 1885-1893.
[36] Phillips R P, Fahey T J. Tree species and mycorrhizal associations influence the magnitude of rhizosphere effects[J]. Ecology, 2006, 87(5): 1302-1313.
[37] 馬海賓, 翟嬋嬋, 王勝坤, 等. 不同分子量殼聚糖抑菌作用的研究[J]. 中國農(nóng)學(xué)通報(bào), 2014, 30(31): 267-271.
[38] 陳紅麗. 腐熟麥秸對(duì)植煙土壤的營養(yǎng)效應(yīng)及其機(jī)理研究[D]. 鄭州: 河南農(nóng)業(yè)大學(xué), 2013.
責(zé)任編輯:謝龍蓮
猜你喜歡
河北科技師范學(xué)院學(xué)報(bào)(2022年2期)2022-08-26 08:55:40
河北科技師范學(xué)院學(xué)報(bào)(2021年1期)2021-05-10 03:34:20
中成藥(2017年12期)2018-01-19 02:06:57
電源技術(shù)(2017年1期)2017-03-20 13:37:59
廣西科技大學(xué)學(xué)報(bào)(2016年1期)2016-06-22 13:10:38
天然產(chǎn)物研究與開發(fā)(2016年1期)2016-06-05 10:29:25
食品界(2016年4期)2016-02-27 07:36:46
中國果菜(2015年2期)2015-03-11 20:01:01
應(yīng)用化工(2014年7期)2014-08-09 09:20:21
應(yīng)用技術(shù)學(xué)報(bào)(2014年4期)2014-02-28 14:52:40
