碧香早雜交F1代生化成分多樣性分析及特異優株篩選
2021-12-18 04:55:02段繼華黃飛毅康彥凱陳瑩玉李賽君
茶葉通訊 2021年4期
羅 意,雷 雨,段繼華,黃飛毅,康彥凱,丁 玎,陳瑩玉,李賽君
湖南省農業科學院 茶葉研究所/國家茶樹改良中心 湖南分中心/農業部 茶樹及茶葉加工科學觀測實驗站/湖南省茶樹品種與種苗工程技術研究中心,湖南 長沙 410125
我國歷來重視茶樹品種的選育與應用,品種資源的豐富度和多樣性也為世界之最[1-2]。雜交育種是茶樹創造遺傳變異的主要方式,也是獲得新品種的重要途徑[3]。茶葉中的生化成分特別是影響茶葉品質的主要生化成分,則是茶類適制性與品質優劣的物質基礎[4-5]。
碧香早為福鼎大白茶×云南大葉種的人工雜交后代,內含物豐富,制紅、綠茶品質兼優[6-8],且雜交親和力強[9]。本試驗研究以從碧香早為親本的7個雜交組合中篩選出的62個F1代優株為材料,探討不同雜交組合間及各優株間生化成分的差異,以期篩選出優異新品系,為碧香早的創新利用及茶樹新品種選育提供基礎。
1 材料與方法
1.1 試驗材料
以碧香早、保靖黃金茶1號、金萱、龍井長葉、菊花春、鐵觀音、云抗10號等7個國家級或省級茶樹品種為雜交親本,根據親本選配的一般原則[3],選配雜交組合7個,獲得631個F1后代,通過生長勢、物候期、抗性等調查從中篩選出62個優株(表1)。育種試驗地設在湖南省長沙縣高橋鎮湖南省茶葉研究所品種試驗基地。
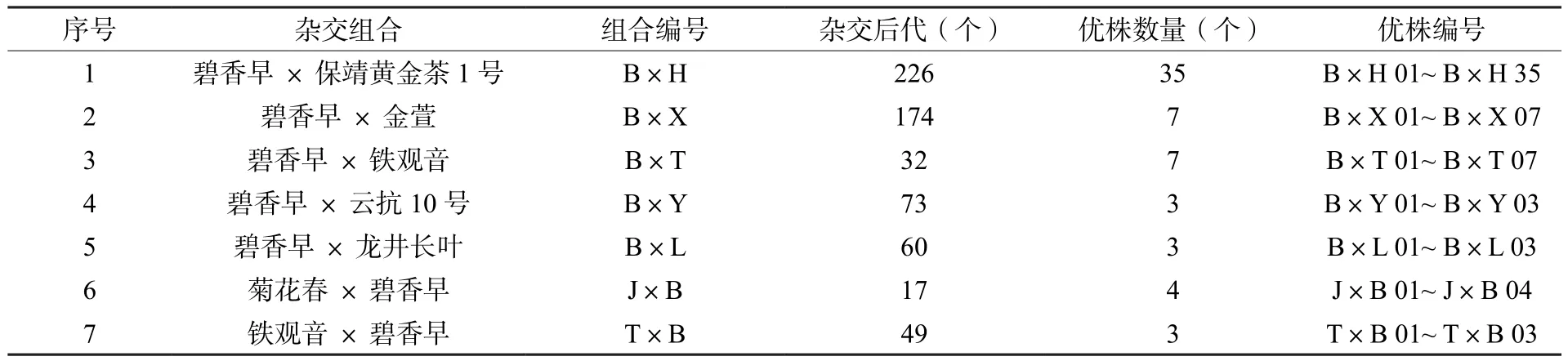
表1 參試材料基本信息Table 1 Basic information of test materials
1.2 試驗方法
1.2.1 樣品制作
采摘春季第1輪新梢的一芽二葉,采用微波殺青制蒸青樣,用于生化成分測定。
1.2.2 生化成分測定
水浸出物、咖啡堿、茶多酚和氨基酸含量分別采用國標GB/T 8305、GB/T 8312、GB/T 8313和GB/T 8314進行測定,各生化成分的測定均為3次重復的平均值。
1.3 統計分析
采用Excel及Spss 20.0進行數據統計分析。計算遺傳多樣性指數(H')前先對數量性狀進行質量化處理,以每個性狀級差的 1/10 為間距將各性狀分為 10 個等級,表型性狀的遺傳多樣性指數采用Shannon-Weave 指數(H') ,計算公式為:H' = -∑PjlnP j,其中Pj為某性狀第j個代碼出現的頻率[4]。
2 結果與分析
2.1 F1代主要生化成分含量及遺傳多樣性
由表2可知, 62個F1代生化成分差異較大。茶多酚含量的變化幅度為15.65% ~ 29.20%,均值為22.36%;氨基酸含量的變化幅度為3.05%~ 6.77%,均值為4.45%;咖啡堿含量的變化幅度為2.35% ~ 4.91%,均值為3.66%;水浸出物含量的變化幅度為34.00% ~ 41.61%,均值為37.04%;酚氨比的變化幅度為2.51 ~ 8.73,均值為5.38。F1代主要生化成分的變異系數最大的是酚氨比(31.11%),其次為氨基酸(23.37%),最小的是水浸出物(4.54%),說明在生化成分的改良上,氨基酸的改良潛力最大,水浸出物的改良潛力最小。
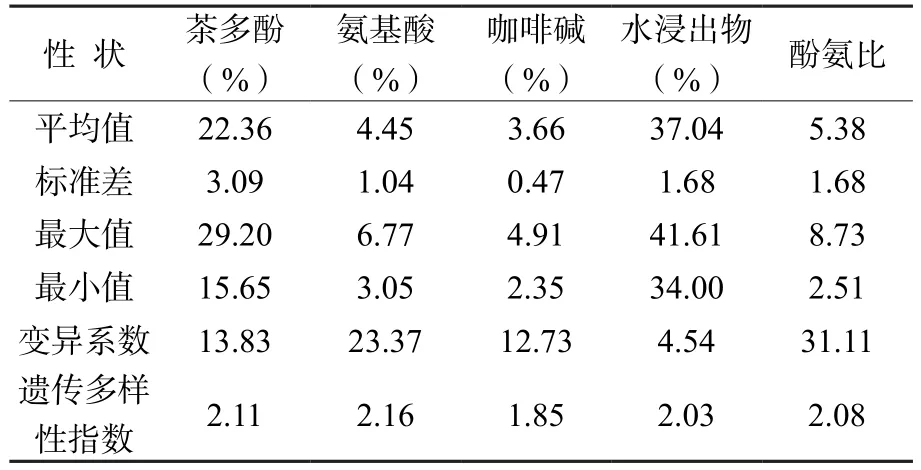
表2 F1代主要生化成分指標Table 2 Main biochemical components of F1 generation
62個F1代優株品質成分平均多樣性指數為2.05%,變化范圍為1.85% ~ 2.16%,具有豐富的遺傳多樣性。其中,氨基酸遺傳多樣性指數最大,為2.16,說明氨基酸含量在62個F1代中多樣性廣泛,包含的信息量較大;其次是茶多酚、酚氨比和水浸出物,分別為2.11、2.08和2.03;最小的是咖啡堿,為1.85。
2.2 F1代主成分分析
由7個雜交組合F1代的生化成分主成分分析可知(表3),共提取2個主成分,累計貢獻率達87.050%,包含了所有性狀的大部分信息。第1主成分特征值為2.942,貢獻率為58.848%,對應的特征向量以酚氨比、氨基酸影響最大,為高度相關因子,其次為茶多酚,其中氨基酸的影響為負,反映的主要是酚氨比的信息;第2主成分特征值為1.410,貢獻率為28.201%,對應的特征向量以水浸出物影響最大。
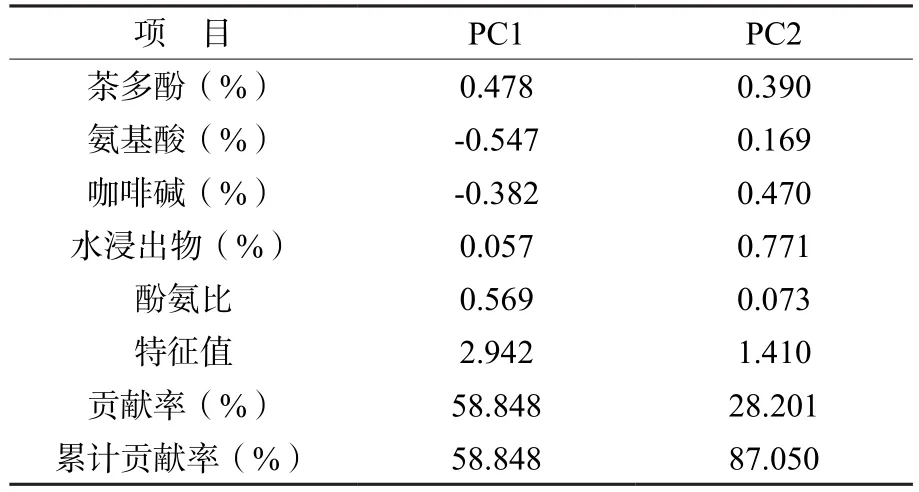
表3 F1代生化成分含量主成分分析Table 3 Principal component analysis of biochemical component content of F1 generation
2.3 F1代聚類分析
對以碧香早為雜交親本F1代62個優株生化成分數據進行聚類(圖1),根據聚類結果將優株分為3大類群(圖1),并對各類群生化成分進行比較分析(表4)。3大類群間各生化成分差異顯著,茶多酚、氨基酸、酚氨比等3大類群間均存在極顯著差異;水浸出物第Ⅱ類群與第Ⅰ和第Ⅲ類群存在極顯著差異;咖啡堿第Ⅲ類群與第Ⅰ和第Ⅱ類群存在極顯著差異。第Ⅰ類群包括30個優株,分別來自B×H組合20個、B×X組合4個、B×Y組合2個、B×L組合1個和J×B組合3個;第Ⅱ類群包括16個優株,分別來自B×H組合11個、B×X組合2個、B×Y組合1個、J×B組合1個和T×B組合1個;第Ⅲ類群包括16個優株,分別來自B×H組合4個、B×X組合1個、B×T組合7個、B×L組合2個和T×B組合2個。
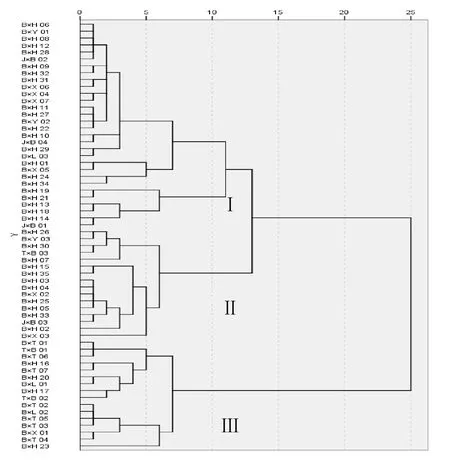
圖1 62個F1代優株聚類圖Figure 1 Clustering diagram of 62 F1 excellent plants
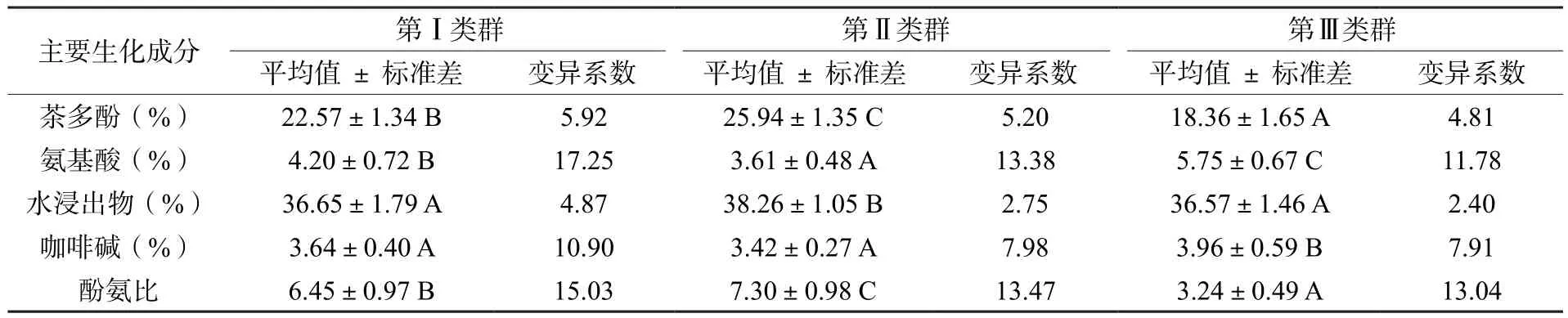
表4 不同類群的主要生化成分比較Table 4 Comparison of main biochemical components of different groups
總體比較,第Ⅱ類群生化成分含量豐富,水浸出物(38.26% ± 1.05%)、茶多酚(25.94%± 1.35%)和酚氨比(7.30 ± 0.98)平均含量最高,適制紅茶;第Ⅲ類群氨基酸平均含量(5.75%± 0.67%)最高,茶多酚含量(18.36% ± 1.65%)及酚氨比(3.24 ± 0.49)平均最低,適制綠茶;第Ⅰ類群優株生化成分含量中等,酚氨比適中(6.45 ± 0.97),屬于紅綠茶兼制。
2.4 不同雜交組合生化成分含量
以碧香早為親本的7個雜交組合間生化成分差異明顯,部分組合在茶多酚、氨基酸、酚氨比上有顯著差異,所有組合的水浸出物、咖啡堿含量無顯著差異(圖2)。雜交組合B×T與B×L間茶多酚含量差異不顯著,但兩者均與其余組合差異顯著;雜交組合B×Y與B×T、B×L的氨基酸含量、酚氨比均差異顯著,B×T與B×Y、B×X的氨基酸含量差異顯著,B×T與B×Y、B×X、B×H的酚氨比差異顯著。
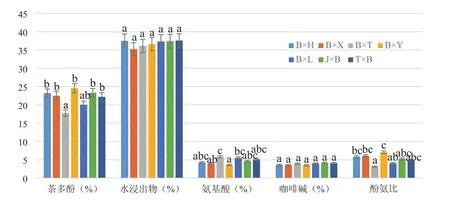
圖2 7個雜交組合生化成分比較Figure 2 Comparison of biochemical components of 7 hybrid combinations
2.5 特異優株篩選
參照《農作物優異種質資源評價規范——茶樹》(NY/T 2031—2011)相關指標,從7個雜交組合F1代的62個優株中初步篩選出高氨基酸(≥5%)特異優株20個,占優株總數的32.26%;這20個優株分別來自除B×Y以外的6個組合,其中組合B×T和B×H優株數最多,各7個,分別占其組合優株數的100%和20%。高茶多酚(≥25%)特異優株13個,占比20.97%;這13個優株分別來自B×H、B×X、B×Y和T×B 等4個組合。高水浸出物(≥40%)特異優株2個,占優株總數的5.71%;這2個優株均來自組合B×H。優株編號為B×H19和B×H21的2個優株氨基酸和水浸出物含量均達特異資源標準。這些特異種質資源可作為選育茶樹特色新品種和雜交親本的材料(表5)。
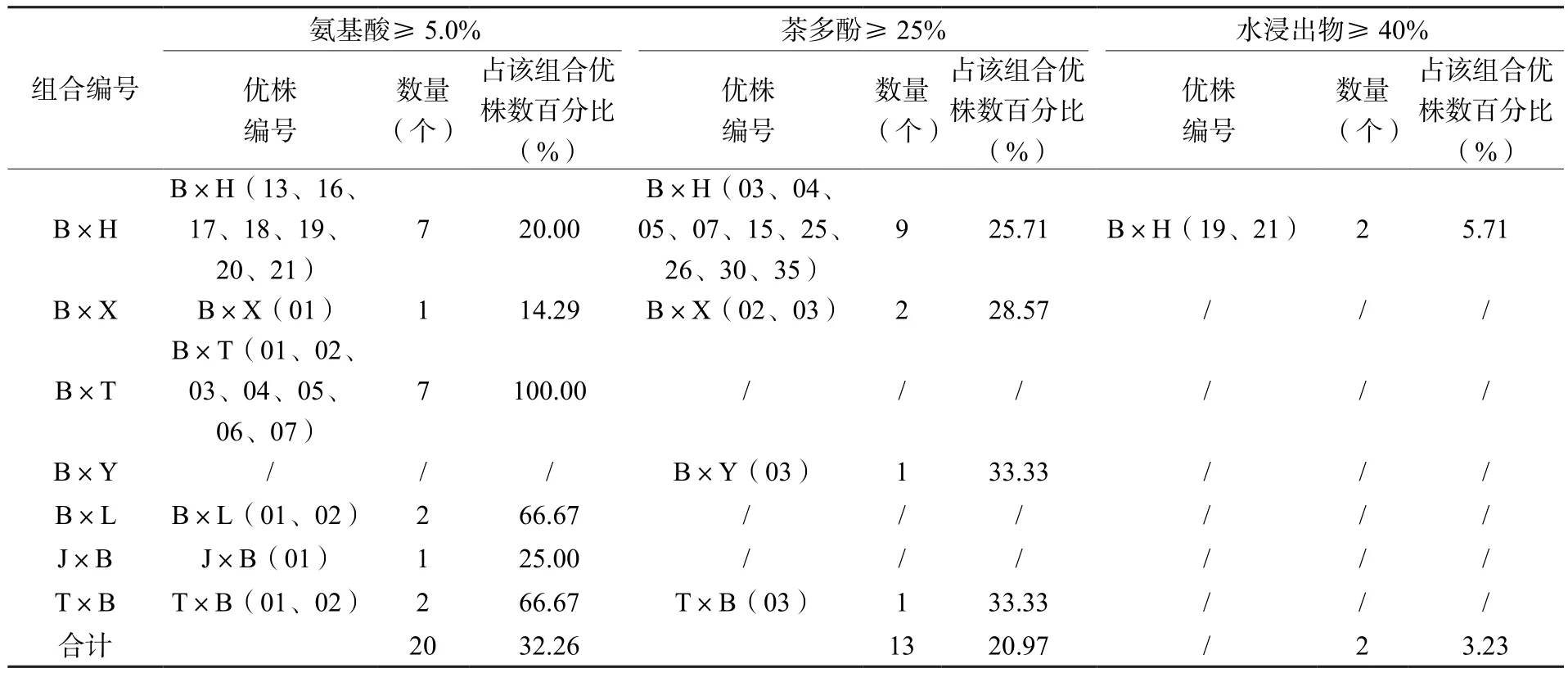
表5 生化成分含量特異優株Table 5 Excellent plants with specific biochemical component content
3 結論與討論
遺傳多樣性是新品種選育的基礎,多樣性指數越高,改良或選育新品種的潛力就越大。本試驗通過對以碧香早為雜交親本的62個F1代優株生化成分進行系統鑒定和評價,發現變異類型較豐富,在酚氨比和氨基酸含量上變異系數較高,說明在酚氨比和氨基酸含量上有很大的選擇潛力,主要生化成分變異系數的大小順序與廣西[4]、貴州[11]等自然群體資源類似;同時,也存在較高的遺傳多樣性,平均指數達到2.05,除咖啡堿外,其余成分遺傳多樣性指數均大于2.0,說明這些生化成分的多樣性指數豐富,具有較好的改良潛力。
對62個F1代優株生化成分進行主成分分析,共提取2個主成分,累計貢獻率達到87.050%,能反應所提供的絕大部分信息,各主成分包含的信息具有一定的相關性,不同的成分在2個主成分中具有不同的載荷值,氨基酸與咖啡堿含量對主成分具有負載荷。以歐氏距離12為閾值將62個F1代優株分為3大類群,3大類群間茶多酚、氨基酸、酚氨比兩兩存在極顯著差異;水浸出物第Ⅱ類群與第Ⅰ和第Ⅲ類群存在極顯著差異;咖啡堿第Ⅲ類群與第Ⅰ和第Ⅱ類群存在極顯著差異。第Ⅰ類群包含30個優株,各優株生化成分含量中等,酚氨比適中,屬紅綠茶兼制類型;第Ⅱ類群包括16個優株,各優株生化成分含量豐富,水浸出物、茶多酚和酚氨比高,多數適制紅茶;第Ⅲ類群包括16個優株,氨基酸含量高,茶多酚含量及酚氨比低,適制綠茶。
組合碧香早×保靖黃金茶1號57.14%(20個)的F1代優株屬于紅綠茶兼制,31.43%(11個)適制紅茶,11.43%(4個)適制綠茶;組合碧香早×鐵觀音的F1代優株全部聚于Ⅲ類,適制綠茶;組合碧香早×龍井長葉及鐵觀音×碧香早各66.67%(2個)的F1代優株聚于Ⅲ類,適制綠茶;組合碧香早×金萱及菊花春×碧香早的F1代優株多數聚于Ⅰ類,屬于紅綠茶兼制。由此可見,以碧香早為母本,鐵觀音和龍井長葉為父本的雜交F1代多適制綠茶,而以保靖黃金茶1號和金萱為父本的F1代多為紅綠茶兼制,這可能與其親本本身特性有關,與李賽君[10]、楊素娟[12]、黃華林[13]和陳宇宏[14]等關于鐵觀音、龍井長葉、金萱、保靖黃金茶1號的適制性相符。
本試驗通過系統鑒定評價,初步篩選出一批生化成分特異的組合及優株。其中以碧香早和鐵觀音為親本的正反交后代均為特異優株(100%);其次為組合碧香早×保靖黃金茶1號(45.71%),且其后代中有2個優株(B×H19和B×H21)在氨基酸和水浸出物含量上均達特異資源標準。這些特異資源的篩選可為特色茶樹新品種選育及茶葉精深加工提供材料。由于茶樹結實率低,親本親和力差異大,導致本試驗研究中有幾個組合后代數量偏少,不能完全評價雜交組合及其優株情況。因此,今后將進一步擴大這幾個雜交組合后代數量,對以碧香早為親本的雜交后代及其組合開展更深入研究,以探明主要生化成分的遺傳規律,篩選出更多優勢組合。
