短發(fā)夾RNA沉默HIPK2對(duì)口腔鱗癌SCC-25細(xì)胞株增殖、自噬和裸鼠移植瘤生長(zhǎng)的影響*
2021-12-20 05:36:34李森森楊再波童國(guó)勇張宏謐
廣西醫(yī)科大學(xué)學(xué)報(bào) 2021年11期
關(guān)鍵詞:檢測(cè)
李森森,楊再波△,朱 彬,童國(guó)勇,張宏謐
(1.湖北恩施土家族苗族自治州中心醫(yī)院口腔診療中心,恩施 445000;2.湖北民族大學(xué)附屬民大醫(yī)院口腔科,恩施 445000)
口腔鱗狀細(xì)胞癌(oral squamous cell carcinoma,OSCC)是常見(jiàn)的惡性腫瘤,占所有頭頸鱗狀細(xì)胞癌(HNSCC)病例的40%,通常表現(xiàn)為侵略性行為,經(jīng)常導(dǎo)致局部浸潤(rùn)和早期淋巴結(jié)轉(zhuǎn)移[1]。大多數(shù)OSCC 患者在診斷時(shí)處于晚期臨床階段,由于手術(shù)難以根治并且預(yù)后較差,OSCC 的5 年生存率<50%[2]。約25%~50%的患者在治療后會(huì)出現(xiàn)局部復(fù)發(fā)和遠(yuǎn)處轉(zhuǎn)移[3]。因此,尋找可靠的生物標(biāo)志物和新的分子靶點(diǎn)對(duì)于開(kāi)發(fā)和實(shí)施OSCC患者的精準(zhǔn)個(gè)性化治療至關(guān)重要。
同源結(jié)構(gòu)域相互作用蛋白激酶2(homeodomain-interacting protein kinase 2,HIPK2)屬于絲/蘇氨酸蛋白激酶家族成員,通過(guò)參與調(diào)控基因表達(dá)對(duì)細(xì)胞周期、凋亡及衰老具有調(diào)控作用[4]。相關(guān)研究表明,HIPK2可作為抑癌基因,抑制乳腺癌、卵巢癌和胰腺癌等腫瘤的發(fā)生和發(fā)展,或表達(dá)上調(diào)發(fā)揮促癌作用[5-7]。HIPK2 在OSCC 中表達(dá)升高,且以?xún)H胞質(zhì)陽(yáng)性表達(dá)為主,與OSCC 的密預(yù)后密切相關(guān)[8]。但HIPK2 對(duì)OSCC 的具體作用機(jī)制尚未見(jiàn)相關(guān)報(bào)道,因此,本文通過(guò)構(gòu)建HIPK2 短發(fā)夾RNA 沉默HIPK2 表達(dá),探究其對(duì)OSCC 細(xì)胞SCC-25 生長(zhǎng)、自噬及裸鼠移植瘤生長(zhǎng)的影響。
1 材料和方法
1.1 實(shí)驗(yàn)試劑
DMEM 培養(yǎng)基、胎牛血清、青—鏈霉素和胰蛋白酶購(gòu)自美國(guó)Gibco公司;人舌磷癌細(xì)胞SCC-25購(gòu)自中國(guó)科學(xué)院典型培養(yǎng)物保藏委員會(huì)細(xì)胞庫(kù);BCA蛋白濃度測(cè)定試劑盒、辣根過(guò)氧化物酶標(biāo)記山羊抗兔IgG 購(gòu) 自Thermo Fisher;一 抗HIPK2、c-Myc、Bax、Bcl-2、Caspase-3、Bcelin1、ATG7、LC3Ⅰ/LC3Ⅱ和Actin購(gòu)自英國(guó)Abcam公司。
1.2 細(xì)胞培養(yǎng)
SCC-25 細(xì)胞置于37 ℃、5%CO2恒溫培養(yǎng)箱中用高糖DMEM培養(yǎng)基(含10%胎牛血清和1%青—鏈霉素)培養(yǎng),每3 d更換1次培養(yǎng)基,收集細(xì)胞時(shí)用0.25%胰蛋白酶消化,試驗(yàn)選用對(duì)數(shù)生長(zhǎng)期細(xì)胞。
1.3 細(xì)胞處理及分組
設(shè)計(jì)兩對(duì)shRNA序列(HIPK2-shRNA1、HIPK2-shRNA2)干擾HIPK2 表達(dá),取對(duì)數(shù)生長(zhǎng)期SCC-25細(xì)胞,將細(xì)胞隨機(jī)分為4組:Control組、shRNA-NC 組、HIPK2-shRNA1 組 和HIPK2-shRNA2組。Control 組不做處理,shRNA-NC 組轉(zhuǎn)染shRNA-NC 陰性對(duì)照載體,HIPK2-shRNA1 組轉(zhuǎn)染HIPK2-shRNA1 慢病毒載體,HIPK2-shRNA2 組轉(zhuǎn)染HIPK2-shRNA2慢病毒載體。
1.4 實(shí)時(shí)熒光定量PCR(RT-qPCR)檢測(cè)HIPK2 mRNA水平
用Trizol法從SCC-25細(xì)胞中提取總RNA,采用PrimeScrip反轉(zhuǎn)錄試劑盒進(jìn)行反轉(zhuǎn)錄成cDNA,通過(guò)SYBR Premix Ex Taq說(shuō)明書(shū)配置PCR反應(yīng)體系,反應(yīng)條件為:95 ℃30 s,95 ℃10 s,60 ℃30 s,35 個(gè)循環(huán)。以Actin為內(nèi)參基因,用2-△△CT計(jì)算各目的基因相對(duì)表達(dá)量。引物序列:HIPK2-shRNA1 上游引物為5'-TGACCACTGTCCACAACCAGCCCTCAG-3',下游引物為5'-TCGCACCGAGTAGCCAGCGTGC-3';HIPK2-shRNA2 上游引物為5'-AGGAAGAGTAAGCAGCACCAG-3',下游引物為5'-TGCTGATGGTGATGACACTGA-3';Actin上游引物為5'-CGATGCCCTGAGGCTCTTT-3',下游引物為5'-TAGTTTCATGGATGCCACAGGAT-3’。
1.5 克隆形成實(shí)驗(yàn)檢測(cè)細(xì)胞增殖
將SCC-25 細(xì)胞接種至6 孔板,約500 個(gè)細(xì)胞每孔,按方法“1.3項(xiàng)”進(jìn)行處理及分組,培養(yǎng)7 d后棄掉上清液,4%多聚甲醛固定20 min,0.1%結(jié)晶紫染色15 min,洗凈干燥后于顯微鏡下觀察克隆形成數(shù)目,計(jì)算克隆形成率。細(xì)胞克隆形成率(%)=細(xì)胞克隆總數(shù)/接種細(xì)胞數(shù)×100%。
1.6 流式細(xì)胞儀檢測(cè)細(xì)胞凋亡
取對(duì)數(shù)生長(zhǎng)期細(xì)胞SCC-25細(xì)胞接種于6孔板,按方法“1.3 項(xiàng)”分組處理,胰酶消化吸收后,PBS 洗滌2 次,加入500 μL 緩沖液懸浮細(xì)胞,再加入5 μL磷脂酰絲氨酸蛋白混勻,最后加入碘化丙啶10 μL室溫避光反應(yīng)20 min 通過(guò)流式細(xì)胞儀檢測(cè)各組細(xì)胞凋亡。
1.7 Western blotting 檢測(cè)HIPK2、c-Myc、Bax、Bcl-2、Caspase-3、Bcelin1、ATG7、LC3Ⅰ和LC3Ⅱ蛋白表達(dá)
各組細(xì)胞加入含蛋白酶抑制劑的細(xì)胞裂解液提取總蛋白,BCA 法測(cè)定蛋白含量。SDS-PAGE 凝膠電泳分離蛋白轉(zhuǎn)移至PVDF 膜。4 ℃下加入HIPK2、c-Myc、Bax、Bcl-2、Caspase-3、Bcelin1、ATG7、LC3Ⅰ和LC3Ⅱ一抗(1∶1 000)孵育過(guò)夜,TBST清洗后加入HRP-IgG(1∶10 000),加入電化學(xué)發(fā)光顯色液顯色。以GAPDH 為內(nèi)參,ImageJ 軟件分析各組細(xì)胞目的蛋白與GAPDH比值。
1.8 裸鼠移植瘤模型的建立
20 只雄性Balb/c 裸鼠購(gòu)自武漢大學(xué)動(dòng)物實(shí)驗(yàn)中心,5 周齡,體重(20±2)g,許可證號(hào):SCXK(鄂)2019—0004,在特定的無(wú)病原體條件下飼養(yǎng)。將裸鼠隨機(jī)分為兩組:Control 組和HIPK2-shRNA2 組,取方法“1.3 項(xiàng)”處理后的Control 組和HIPK2-shRNA2 組處于對(duì)數(shù)生長(zhǎng)期的SCC-25 細(xì)胞,分別注射于裸鼠后背部靠近腋窩處皮下(1×107個(gè)/mL,0.2 mL/只)。達(dá)成瘤標(biāo)準(zhǔn)(瘤徑≥0.5 cm)進(jìn)行后續(xù)實(shí)驗(yàn)。處死所有裸鼠,稱(chēng)取腫瘤重量,TUNEL 染色檢測(cè)細(xì)胞凋亡,免疫組化檢測(cè)Caspase-3 陽(yáng)性表達(dá)率,western blotting檢測(cè)LC3Ⅰ、LC3Ⅱ蛋白表達(dá)水平。
1.9 TUNEL染色
將裸鼠腫瘤組織切片用二甲苯浸洗2 次脫蠟,每次5 min,梯度乙醇(100%、95%、90%、80%、70%)各浸洗1 次,每次3 min。參照TUNEL 凋亡試劑盒說(shuō)明書(shū),用蛋白酶K 工作液在37 ℃下封閉20 min,PBS 清洗2 次。在每個(gè)切片樣本上加入TUNEL 反應(yīng)液50 μL,37 ℃閉光反應(yīng)60 min。用DAPI 復(fù)染10 min后熒光顯微鏡下觀察。
1.10 免疫組化測(cè)定察Caspase-3蛋白的表達(dá)情況
取各組裸鼠腫瘤組織,常規(guī)石蠟包埋制成厚度約4 μm的切片,按免疫組化檢測(cè)試劑盒說(shuō)明書(shū)進(jìn)行免疫組化染色,陽(yáng)性表達(dá)為細(xì)胞質(zhì)出現(xiàn)棕黃色顆粒,采用光學(xué)顯微鏡觀察Caspase-3在腫瘤組織中的表達(dá)情況。
1.11 統(tǒng)計(jì)學(xué)方法
采用GraphPad Prism 9 軟件對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析,計(jì)量資料采用均數(shù)±標(biāo)準(zhǔn)差()表示,組間比較采用t檢驗(yàn),多組間比較采用單因素方差分析(One-Way ANOVA),組間兩兩比較采用LSD-t檢驗(yàn)。以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 各組HIPK2 mRNA和蛋白表達(dá)水平
與Control 組比較,HIPK2-shRNA1 組、HIPK2-shRNA2組HIPK2 mRNA水平和蛋白表達(dá)水平均明顯降低(P<0.05),見(jiàn)圖1。
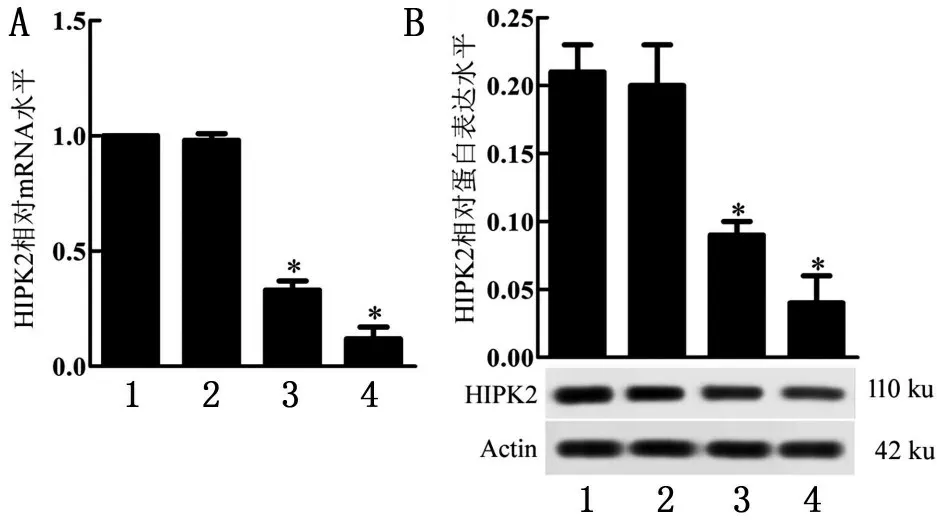
圖1 HIPK2 mRNA水平和蛋白表達(dá)水平
2.2 HIPK2 短發(fā)夾RNA 對(duì)SCC-25 細(xì)胞克隆形成率的影響
與Control 組比較,HIPK2-shRNA1 組、HIPK2-shRNA2 組細(xì)胞克隆形成率明顯降低,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05),見(jiàn)圖2。

圖2 HIPK2短發(fā)夾RNA對(duì)SCC-25細(xì)胞克隆形成率的影響
2.3 HIPK2 短發(fā)夾RNA 對(duì)SCC-25 細(xì)胞凋亡率的影響
與Control組比較,HIPK2-shRNA1、HIPK2-shRNA2 組細(xì)胞凋亡率明顯升高,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05),見(jiàn)圖3。

圖3 HIPK2短發(fā)夾RNA對(duì)SCC-25細(xì)胞凋亡率的影響
2.4 HIPK2 短發(fā)夾RNA 對(duì)SCC-25 細(xì)c-Myc、Bax、Bcl-2、Caspase-3蛋白表達(dá)的影響
與Control組比較,HIPK2-shRNA1、HIPK2-shRNA2 組c-Myc 蛋白表達(dá)明顯降低,cleaved Caspase-3/Caspase-3、Bax/Bcl-2 比值顯著升高(P<0.05),見(jiàn)圖4。

圖4 HIPK2短發(fā)夾RNA對(duì)SCC-25細(xì)c-Myc、Bax、Bcl-2、Caspase-3蛋白表達(dá)的影響
2.5 HIPK2短發(fā)夾RNA 對(duì)SCC-25細(xì)胞Bcelin1、ATG7、LC3Ⅰ、LC3Ⅱ蛋白表達(dá)的影響
與Control 組比較,HIPK2-shRNA1 組、HIPK2-shRNA2 組Bcelin1、ATG7 蛋白表達(dá)水平和LC3Ⅱ/LC3Ⅰ比值顯著升高,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05),見(jiàn)圖5。

圖5 HIPK2短發(fā)夾RNA對(duì)SCC-25細(xì)胞Bcelin1、ATG7、LC3Ⅰ、LC3Ⅱ蛋白表達(dá)的影響
2.6 HIPK2短發(fā)夾RNA 對(duì)口腔鱗狀細(xì)胞癌裸鼠移植瘤模型的影響
與Control 組比較,HIPK2-shRNA2 組裸鼠腫瘤重量降低,Caspase-3陽(yáng)性細(xì)胞數(shù)、細(xì)胞凋亡率、LC3Ⅱ/LC3Ⅰ比值明顯升高(P<0.05),見(jiàn)圖6。
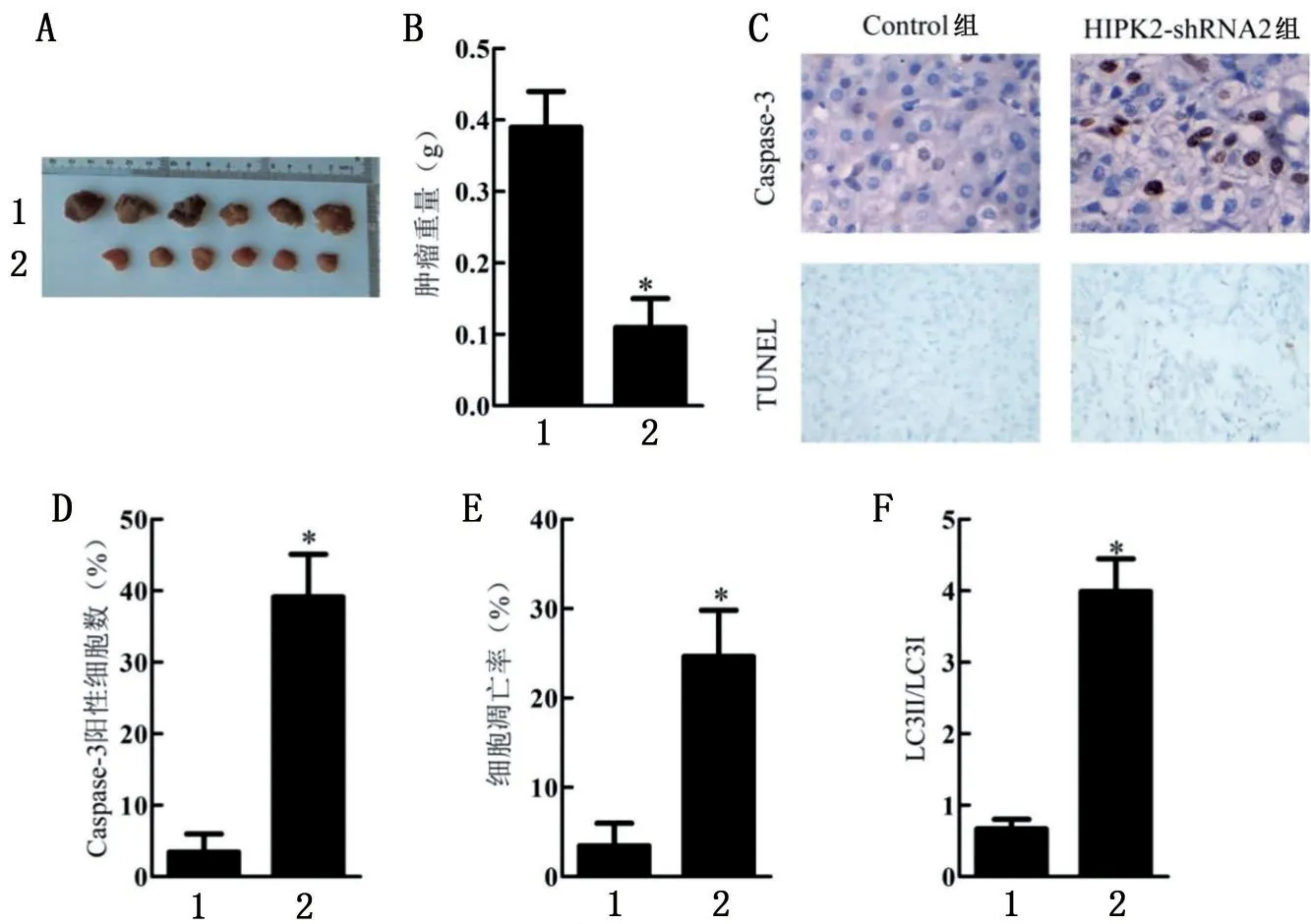
圖6 HIPK2短發(fā)夾RNA對(duì)口腔鱗狀細(xì)胞癌裸鼠移植瘤模型的影響
3 討論
OSCC發(fā)病機(jī)制復(fù)雜,涉及多種遺傳因素、生活方式和代謝風(fēng)險(xiǎn),但其具體分子機(jī)制尚不明確。因此,需要探索新的分子治療靶點(diǎn),并通過(guò)更好地闡明分子機(jī)制和功能意義來(lái)發(fā)現(xiàn)OSCC有效的治療方式。
增殖能力的增強(qiáng)及凋亡的減弱是腫瘤細(xì)胞重要的生物學(xué)特征,也是腫瘤發(fā)生發(fā)展的重要生物學(xué)基礎(chǔ)[9]。抑制腫瘤細(xì)胞增殖,誘導(dǎo)腫瘤細(xì)胞凋亡是治療OSCC 的有效手段。c-Myc 為原癌基因,可介導(dǎo)腫瘤細(xì)胞無(wú)限增殖,促進(jìn)腫瘤細(xì)胞凋亡,相關(guān)研究表明,c-Myc高表達(dá)可促進(jìn)OSCC的發(fā)生[10]。與細(xì)胞凋亡相關(guān)的基因Bax 升高能夠誘導(dǎo)細(xì)胞凋亡,Bcl-2 是細(xì)胞凋亡的抑制劑,Bax/Bcl-2 比值能反應(yīng)細(xì)胞凋亡狀況[11]。細(xì)胞中的Caspase-3一般無(wú)活性,以procaspase-3 形式存在,其活化后剪切下游分子,進(jìn)而誘導(dǎo)細(xì)胞凋亡[12]。Cheng等[13]研究發(fā)現(xiàn),HIPK-2 表達(dá)降低可一起宮頸癌細(xì)胞凋亡增加,增殖減少。Puca 等[14]研究發(fā)現(xiàn),HIPK2 可通過(guò)激活p53 抑癌基因細(xì)胞凋亡活性,從而促進(jìn)乳腺癌細(xì)胞凋亡。本文研究表明,短發(fā)夾RNA沉默HIPK2表達(dá)可升高SCC-25 細(xì)胞凋亡率,可降低SCC-25 細(xì)胞克隆形成率,降低裸鼠移植瘤腫瘤重量,降低c-Myc蛋白表達(dá)水平,升高cleaved Caspase-3/Caspase-3、Bax/Bcl-2比值,升高裸鼠移植瘤腫瘤細(xì)胞Caspase-3陽(yáng)性細(xì)胞數(shù)和細(xì)胞凋亡率。提示HIPK2 短發(fā)夾RNA 可抑制SCC-25細(xì)胞增殖和裸鼠移植瘤腫瘤的生長(zhǎng),并誘導(dǎo)SCC-25細(xì)胞和裸鼠移植瘤腫瘤細(xì)胞凋亡。
自噬在腫瘤的發(fā)生發(fā)展中具有促進(jìn)和抑制的雙重作用[15]。Beclin1基因,位于染色體17q21上,可作為抑癌基因參與大部分惡性腫瘤的發(fā)生發(fā)展,相關(guān)研究表明,Beclin1在口腔鱗癌組織中的陽(yáng)性表達(dá)率顯著低于癌旁組織,與腫瘤分化及淋巴結(jié)轉(zhuǎn)移有關(guān)[16]。ATG7是自噬調(diào)節(jié)的重要蛋白,可通過(guò)調(diào)節(jié)多種酶的活性從而激活細(xì)胞的自噬及凋亡過(guò)程[17]。自噬蛋白LC3 可分為L(zhǎng)C3-Ⅰ和LC3-Ⅱ兩種,LC3-Ⅱ定位在雙層膜和自噬體上,是自噬體的特異性標(biāo)志分子,LC3Ⅱ/LC3Ⅰ的比值與自噬泡的數(shù)量呈正相關(guān),其比值大小是判定自噬水平的高低的依據(jù)[18]。本研究發(fā)現(xiàn),HIPK2 短發(fā)夾RNA 具有升高SCC-25細(xì)胞Bcelin1、ATG7 蛋白表達(dá)水平和LC3Ⅱ/LC3Ⅰ比值,升高裸鼠移植瘤腫瘤細(xì)胞LC3Ⅱ/LC3Ⅰ比值。提示HIPK2短發(fā)夾RNA可誘導(dǎo)并激活SCC-25細(xì)胞和裸鼠移植瘤腫瘤細(xì)胞自噬,促進(jìn)腫瘤細(xì)胞凋亡。
綜上所述,HIPK2 短發(fā)夾RNA 可抑制SCC-25細(xì)胞增殖,誘導(dǎo)細(xì)胞凋亡和自噬,抑制裸鼠移植瘤模型移植瘤的生長(zhǎng)。HIPK2有望成為治療OSCC的新靶點(diǎn)。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
 廣西醫(yī)科大學(xué)學(xué)報(bào)2021年11期
廣西醫(yī)科大學(xué)學(xué)報(bào)2021年11期
- 廣西醫(yī)科大學(xué)學(xué)報(bào)的其它文章
- 多發(fā)性骨髓瘤自體干細(xì)胞移植后中樞神經(jīng)系統(tǒng)復(fù)發(fā)1例并文獻(xiàn)復(fù)習(xí)
- 19 067例無(wú)償獻(xiàn)血志愿者HBV、HIV感染篩查結(jié)果分析*
- 右右美托咪定對(duì)七氟醚在小兒血管畸形介入術(shù)中的麻醉效果觀察*
- 原發(fā)性卵巢癌肉瘤16例臨床分析*
- 肺結(jié)核患者血清促甲狀腺激素和糖化血紅蛋白的相關(guān)性分析*
- 支氣管肺泡灌洗液液基細(xì)胞學(xué)聯(lián)合腫瘤標(biāo)志物鑒別良惡性孤立性肺結(jié)節(jié)的價(jià)值*