鹽度脅迫對河蜆攝食率及鰓ATP酶活力變化研究
2021-12-22 11:01:40劉一萌來琦芳么宗利高鵬程
海洋漁業 2021年6期
關鍵詞:實驗
包 錟,劉一萌,來琦芳,么宗利,周 凱,高鵬程
(1.上海海洋大學水產與生命學院,上海 201306;2.中國水產科學研究院東海水產研究所,鹽堿水域漁業工程技術研究中心,上海 200090)
我國約有4.6×105km2的低洼鹽堿水域和約占全國湖泊面積55%的咸水湖泊[1],近年來利用鹽堿水開展水產養殖逐漸增多。貝類以攝食水中的藻類和有機碎屑為主[2-5],在鹽堿水域混養貝類對鹽堿水的利用具有積極意義。鹽度是鹽堿生境中最主要的脅迫因子,也是水生動物面臨最常見最主要的環境因子之一,在鹽堿水環境生活的貝類會受到鹽度帶來的脅迫。
雙殼貝類在受到脅迫時會將殼緊閉以盡量減少機體與外界環境中的離子交換,這種保護機制在砂海螂(Mya arenaria)、淺溝蛤(Scrobicularia plana)、紫貽貝(Mytilus edulis)、鳥蛤(Cardium edule)、硬殼蛤(Mercenaria mercenaria)、太平洋牡蠣(Crassostrea gigas)、泥蚶(Tegillarca granosa)、魁蚶(Anadara broughtonii)、毛 蚶(Scapharca subcrenata)等雙殼貝類中均有發現[6-9]。由于雙殼貝類閉殼可以存活很長時間,且很難觀察其生存情況,因此急性毒性實驗無法快速判定雙殼貝類對脅迫的耐受能力,如何選擇快速有效的實驗方法科學評價其對脅迫的耐受性能,需要進行相關探究。
攝食率(ingestion rate)可以反映濾食性貝類生理活動情況,常作為貝類受到脅迫時生物能量方面的重要參數[10];鹽度變化伴隨著Na+、K+、Mg2+和Ca2+等陽離子和Cl-、、和等陰離子的濃度變化,鰓作為主要離子和滲透調節器官,鰓ATP酶在受到鹽度脅迫時發揮著重要作用。Na+/K+-ATP酶,又稱鈉鉀泵(sodium-potassium pump),正常狀態下與質膜內側Na+、Mg2+結合后被激活,使ATP分解,每分解1個ATP分子運進3個Na+和2個K+[11-12]。有研究表明,鹽度發生變化時巖扇貝(Crassadoma gigantea)、泥蚶、魁蚶、文蛤(Meretrix meretrix)、青蛤(Cyclina sinensis)等雙殼貝類鰓Na+/K+-ATP酶均有顯著變化[8-9,13-15]。Ca2+-ATP酶,又稱鈣泵(calcium pump),受到鹽度脅迫時能將Ca2+從細胞質基質中逆濃度梯度運送到細胞外或者內質網腔,使細胞質基質中Ca2+濃度保持在低水平,這一過程與肌肉放松有關[12]。EKA等[16]認為,Mg2+-ATP酶的底物是MgATP,游離ATP和Mg2+是具有雙重作用的酶修飾劑,Mg2+-ATP酶每分解1個ATP轉運1個Mg2+。許多研究證明,鹽度變化與水生動物Na+/K+-ATP酶、Ca2+-ATP酶以及Mg2+-ATP酶的活力變化具有相關性[17-20]。
河蜆(Corbicula fluminea)是一種常見的淡水廣鹽性雙殼貝類,隸屬于軟體動物門(Mollusca),瓣 鰓 綱(Lamellibranchia),真 鰓 瓣 目(Eulamellibranchia),蜆科(Corbiculidae),蜆屬,在淡水、湖泊、河口潮間帶等水域有廣泛分布,對鹽度變化有較強的適應能力。本文以河蜆為實驗對象,模擬不同鹽度脅迫,觀察河蜆攝食率、鰓Na+/K+-ATP酶、Ca2+-ATP酶和Mg2+-ATP酶的活力變化,評估河蜆的耐鹽性能,篩選適宜評價貝類耐鹽性能的指示指標,探索雙殼貝類脅迫耐受性能評價方法。
1 材料與方法
1.1 實驗動物及其暫養
實驗用河蜆采自江蘇東臺河道(120°33′E、33°26′N),使用500 L聚乙烯塑料桶進行暫養1個月以上,持續充氣,每天更換經曝氣24 h的過濾自來水,換水量為三分之一,水溫(20.5±0.2)℃,pH(8.06±0.07),溶氧(6.65±0.34)mg·L-1,每天上午和下午各投喂一次球等鞭金藻(Isochrysis galbana)、叉鞭金藻(Dicrateriasp.)和角毛藻(Chaetocerossp.)按1∶1∶1富集后的混合物。挑選殼長(3.44±0.35)cm、殼寬(2.75±0.13)cm、殼高(1.80±0.12)cm的個體用于后續實驗。
1.2 攝食率實驗
1.2.1 鹽度設置
實驗設置8個鹽度梯度,分別為鹽度3、5、8、10、13、15、20和25,以淡水為對照組,另外本實驗還設置了無貝對照組,以消除藻類繁殖和沉降對藻類計數的影響。實驗用淡水為經凈水器(開能,AC/KDF-150 C型)過濾的自來水,鹽度實驗用水由海水素(青島海科通用海水素有限公司)和過濾自來水配制而成。每個處理組設置3個平行。
1.2.2 實驗管理
實驗在聚乙烯塑料箱(37.5 cm×27.5 cm×14.0 cm)中進行,實驗水體為10 L,每個處理組投放河蜆3個。河蜆在實驗水體中穩定3 h后開始實驗。每個處理組投喂混合藻類(球等鞭金藻、叉鞭金藻和角毛藻),水體藻類濃度為(3.5±0.3)×104個·mL-1,實驗持續24 h,每小時用吸管隨機取樣5 mL后用魯戈氏液固定,光鏡下用血球計數板對藻類計數,重復3次。實驗結束后將河蜆軟體部取出并在60℃烘箱中烘干至恒重(精確到0.001 g)(表1)。
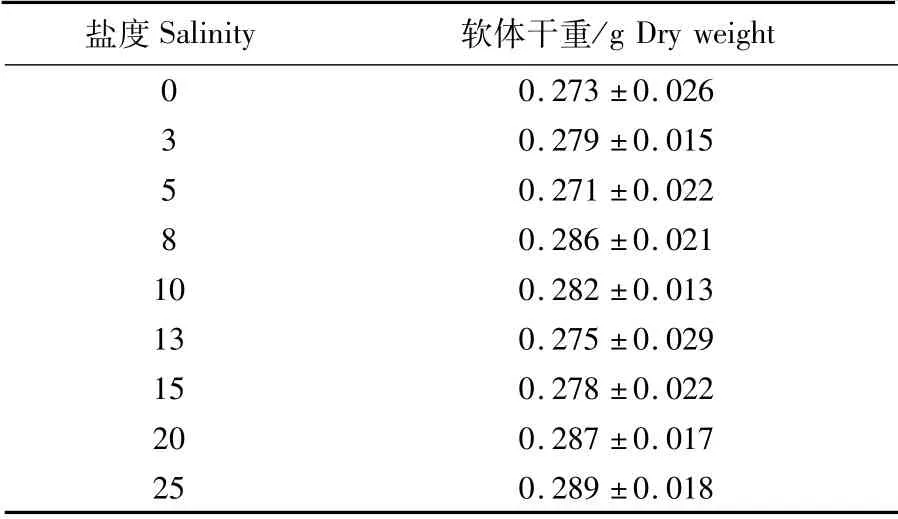
表1 攝食率各組河蜆軟體平均干重Tab.1 Average dry weight of Corbicula fluminea in each ingestion rate test group
1.2.3 攝食率計算
藻類濃度變化系數(S)根據下式計算:

式(1)中:t為實驗時間(h),Cc0為無貝組水體中藻類的初始濃度(個·mL-1),Cct為t小時后無貝組水體中藻類濃度(個·mL-1)。
河蜆攝食率(R)根據下式計算:
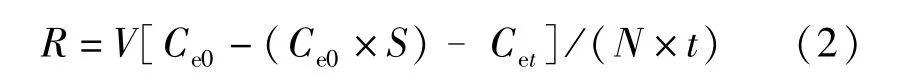
式(2)中:R為攝食率(個·h-1),V為實驗水體量(L),N為實驗貝個數,Ce0為實驗水體中藻類的初始濃度(個·mL-1),Cet為t小時后實驗水體藻類濃度(個·mL-1),S為無貝組藻類濃度變化系數,由式(1)計算所得。
為消除河蜆個體質量對攝食率的影響,將獲得的數據通過下式轉換為1 g標準下的攝食率(Rs):

式(3)中:Rs為1 g標準下的值[個·(g·h)-1];W為軟體干重(g);b為重量指數,參照呂昊澤等[21]的研究,取0.62;R為攝食率(個·h-1),由式(2)計算所得。
1.3 酶活實驗
1.3.1 鹽度設置
參照河蜆攝食率結果,選取3個鹽度梯度,分別為5(S5)、15(S15)和25(S25)。設置淡水為對照組(control),實驗用水同1.2.1。每個處理組設置3個平行。
1.3.2 實驗管理
實驗在聚乙烯塑料箱中進行,實驗水體為10 L,每個梯度設置3個重復,每個處理組投放河蜆10個。實驗期間每天投喂混合藻類2次,水體藻類濃度為(3.5±0.3)×104個·mL-1,并在投喂后3 h進行換水,換水量為三分之一,持續脅迫7 d。每天隨機選取各鹽度組中3個河蜆個體,參照林崗等[22]實驗方法采集新鮮鰓組織樣品制成2%組織勻漿,-80℃保存,待測。ATP酶活力單位定義:每小時每毫克蛋白分解ATP產生1μmol無機磷的量為一個ATP酶活力單位,即μmolPi/mgprot/hour(U·mgprot-1)。具體操作參照南京建成生物工程研究所的考馬斯亮藍試劑盒(A045-2)和ATP酶試劑盒(A016-1)說明書。
1.4 數據統計與分析
運用統計軟件SPSS 20進行統計分析,用one-way ANOVA進行單因素方差分析,若差異顯著,用Tukey檢驗進行多重比較,分析顯著性水平為P<0.05,極顯著性水平為P<0.01。所得數據以3個平行組數據的平均值±標準差表示,并使用Origin9制圖。
2 結果與分析
2.1 鹽度脅迫對河蜆攝食率的影響
如圖1所示,河蜆攝食率隨鹽度的升高而下降。多重比較后發現,鹽度3組攝食率與對照組相比無顯著差異(P>0.05),鹽度5、15和25組河蜆攝食率均有極顯著降低(P<0.01)。分別為2.11×108個·(g·h)-1、1.09×108個·(g·h)-1和7.68×107個·(g·h)-1。
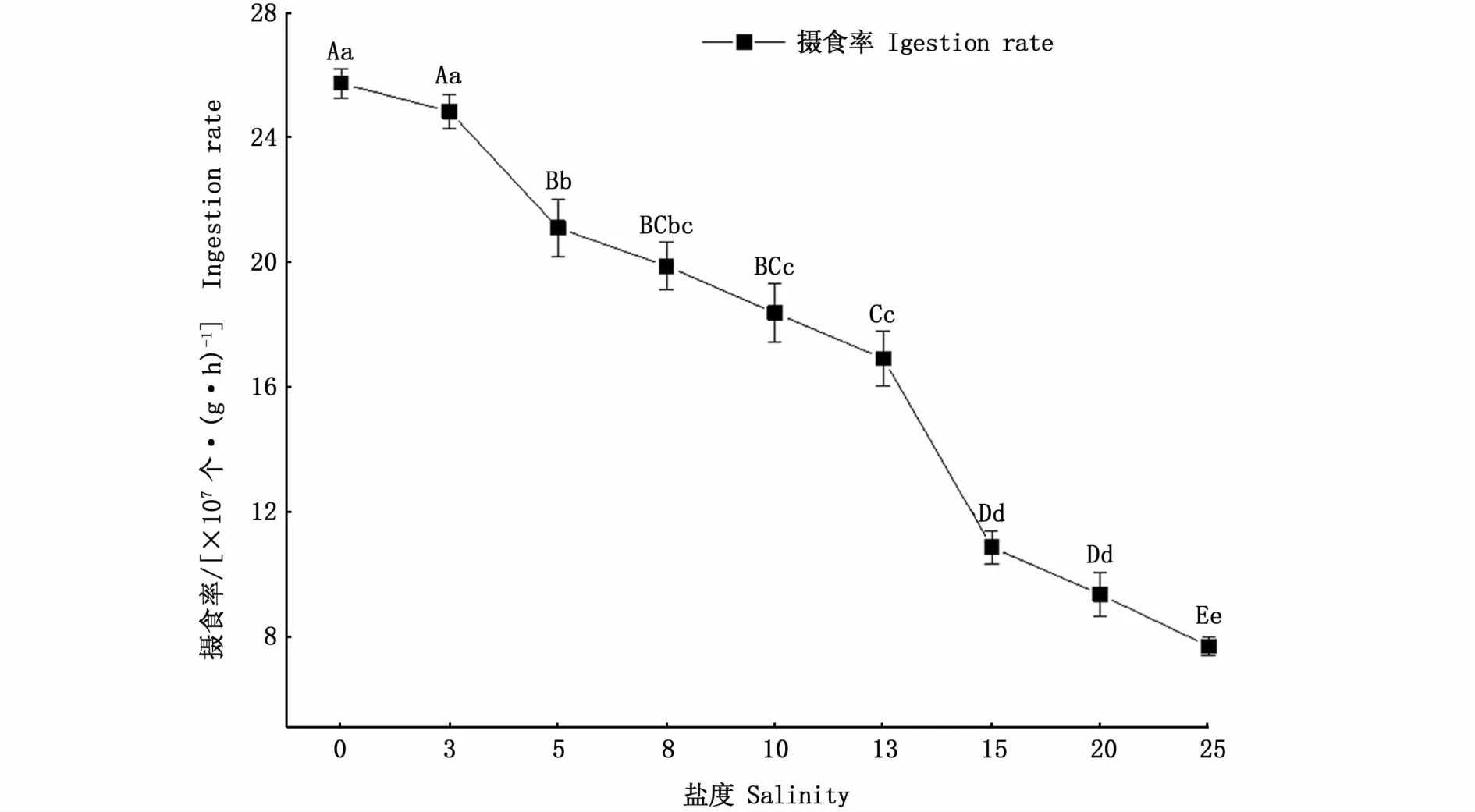
圖1 鹽度脅迫下河蜆攝食率的變化Fig.1 Changes of ingestion rate of Corbicula fluminea under salinity stress
2.2 鹽度脅迫下河蜆鰓Na+/K+-ATP酶活力變化
與對照組相比,不同鹽度脅迫下鹽度5和15組Na+/K+-ATP酶活力變化趨勢相似,1 d時酶活力極顯著降低(P<0.01),之后逐步上升至4 d達到最高(P<0.01),分別為8.53 U·mgprot-1和7.30 U·mgprot-1。從5 d開始,酶活力恢復到初始值附近,并保持穩定(P>0.05);鹽度25組雖然在1 d時酶活力也極顯著降低(P<0.01),之后均與各組相比無顯著差異(P>0.05),7 d時出現大量死亡,存活率僅19%(圖2),存活的河蜆酶活力基本穩定在初始值附近(圖3),其余各組在各時間點內均沒有出現死亡現象。
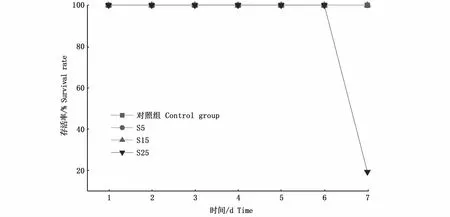
圖2 鹽度脅迫下河蜆的存活率Fig.2 Survival rate of Corbicula fluminea under salinity stress
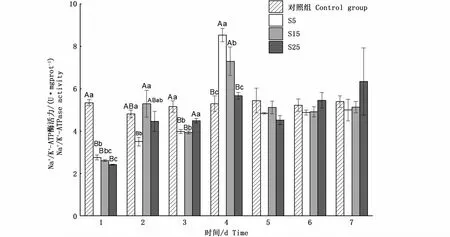
圖3 鹽度脅迫下河蜆鰓Na+/K+-ATP酶的活力變化Fig.3 Changes of Na+/K+-ATPase activity in gill of Corbicula fluminea under salinity stress
2.3 鹽度脅迫下河蜆鰓Ca2+-ATP酶活力變化
與對照組相比,不同鹽度脅迫下鹽度5和15組Ca2+-ATP酶活力變化趨勢相似,2 d時酶活力極顯 著 降 低(P<0.01),分 別 為3.04 U·mgprot-1和3.28 U·mgprot-1,之后逐步上升,4 d時達到峰值,分別為6.26 U·mgprot-1和6.66 U·mgprot-1,7 d時恢復到初始值附近(P>0.05);鹽度25組除1 d、2 d和5 d時極顯著降低(P<0.01),其余時間點酶活力均在初始值附近(P>0.05)(圖4)。
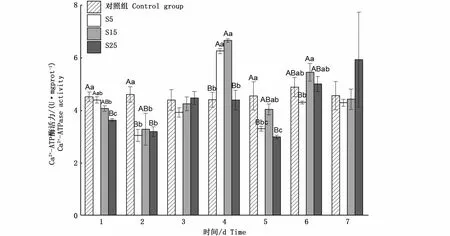
圖4 鹽度脅迫下河蜆鰓Ca2+-ATP酶的活力變化Fig.4 Changes of Ca2+-ATPase activity in gill of Corbicula fluminea under salinity stress
2.4 鹽度脅迫下河蜆鰓Mg2+-ATP酶活力變化
與對照組相比,不同鹽度脅迫下鹽度5和15組Mg2+-ATP酶活力變化趨勢相似,1 d時酶活力極顯 著 降 低(P<0.01),分 別 為5.07 U·mgprot-1和4.55 U·mgprot-1,4 d時極顯著升高并達到峰值(P<0.01),分別為9.91 U·mgprot-1和8.64 U·mgprot-1,從5 d開始,酶活力恢復到初始值附近,并保持穩定(P>0.05);鹽度25組除1 d和3 d時極顯著降低(P<0.01),其余時間點酶活力均穩定在初始值附近(P>0.05)(圖5)。
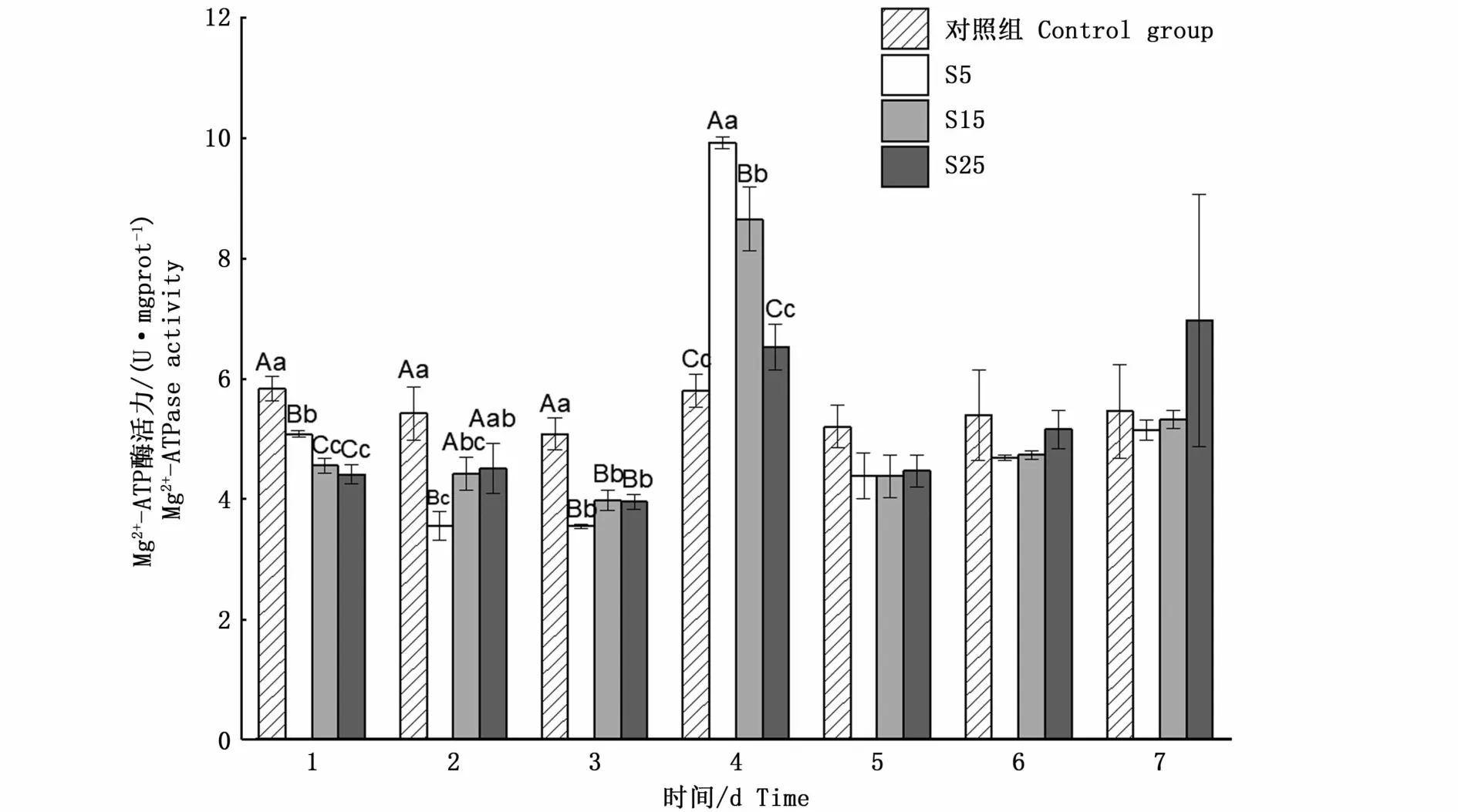
圖5 鹽度脅迫下河蜆鰓Mg2+-ATP酶的活力變化Fig.5 Changes of Mg2+-ATPase activity in gill of Corbicula fluminea under salinity stress
3 討論
3.1 鹽度脅迫對河蜆攝食率的影響
本實驗結果表明,河蜆攝食率與鹽度呈負相關。河蜆在進行鹽度脅迫時,攝食率在鹽度5、15和25出現了相較于上一鹽度極顯著下降的現象。鹽度5組攝食率相較于對照組下降了18%,推測鹽度5對河蜆產生了脅迫;鹽度15組河蜆攝食率相較于對照組下降了58%,背角無齒蚌(Anodonta woodiana)[23]在鹽度10脅迫下攝食率下降約50%,并開始出現死亡現象,雖然河蜆鹽度15脅迫7 d未出現死亡現象,但鹽度25脅迫下河蜆攝食率極低,7 d后出現大量死亡現象,推測鹽度25已超出河蜆耐鹽極限,河蜆耐鹽臨界值在鹽度15附近。鹽度脅迫下由于貝類閉殼,導致攝食率下降,當攝食率下降超過50%時,提示著該鹽度已超出貝類鹽度耐受范圍。另外,海水貝類文蛤和青蛤[24]在經過鹽度5急性脅迫后攝食率相較于鹽度15和20組也嚴重下降,因此攝食率是可以作為比較快速地體現雙殼貝類耐鹽性能的生理指標,攝食率下降比例與耐鹽范圍有一定相關性。
3.2 鹽度脅迫對河蜆鰓ATP酶的影響
本實驗中,鰓Na+/K+-ATP酶活力升高后恢復至正常水平,推測部分水生動物在高鹽脅迫后,可以通過Na+/K+-ATP酶活力在4 d左右時升高,積極進行離子調節,以此建立一種勢能儲備,供細胞其他耗能過程利用[12]。泥蚶[9]在鹽度4脅迫、魁蚶稚貝[8]在鹽度15脅迫時同樣出現4 d升高、5 d恢復的現象;青鳉(Oryzias latipes)和凡納濱對蝦(Litopenaeus vannamei)[25-26]在鹽度脅迫下鰓Na+/K+-ATP酶活力也出現相似的現象。推測水生生物鹽度脅迫后可能有4 d的適應調節期,貝類鰓Na+/K+-ATP酶活力在4 d內無顯著上調趨勢時,預示著該鹽度已超出貝類鹽度耐受范圍。
鰓Ca2+-ATP酶與Mg2+-ATP酶活力變化趨勢與Na+/K+-ATP酶活力變化趨勢類似。推測4 d左右時其將大量細胞質基質中Ca2+轉運到肌質網腔中儲存起來,抑制肌細胞收縮[12],這可能與貝類在受脅迫時的閉殼有關,Ca2+-ATP酶活力的上升促進開殼正常濾水攝食。由于貝類的閉殼行為受肌肉收縮這一耗能過程控制,且貝類行為可能與水中Mg2+濃度有關[27-28],推測Ca2+-ATP酶與Mg2+-ATP酶活力的變化與閉殼之間可能存在相關性。
受到鹽度脅迫時,河蜆首先閉殼應對脅迫,導致攝食率隨之下降[29]。在鹽度脅迫耐受范圍內,通過Na+/K+-ATP酶活升高進行離子調節,同時激活Ca2+-ATP酶與Mg2+-ATP酶來解除閉殼狀態,促進貝類的正常生理活動;如果超出鹽度耐受范圍,ATP酶活力在4 d內不會上升進行適應調節,河蜆始終處于閉殼狀態,攝食率處于極低水平,最終導致死亡。因此鰓Na+/K+-ATP酶亦可作為雙殼貝類耐鹽性能的指示指標。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
