中華鱉組蛋白H2A變體克隆及其在卵母細胞中的表達分析
2021-12-24 05:18:40祝駿賢劉曉莉王亞坤洪孝友于凌云徐紅艷朱新平
水生生物學報 2021年6期
關鍵詞:信號
祝駿賢 陳 辰 劉曉莉 王亞坤 雷 駱 洪孝友 于凌云 徐紅艷 李 偉* 朱新平 *
(1. 南京農業大學無錫漁業學院, 無錫 214081; 2. 中國水產科學研究院珠江水產研究所農業農村部熱帶亞熱帶水產資源利用與養殖重點實驗室, 廣州 510380)
中華鱉(Pelodiscus sinensis)是我國常見的水產養殖品種, 年產量達30余萬噸[1]。一直以來, 水產養殖業選取效益較高的性別進行單性養殖, 是很多學者研究的目標, 中華鱉養殖業也具有高效全雄繁育的需求。近些年, 雖然國內外學者圍繞中華鱉的性別控制及單性繁育已開展了一系列的探索性研究,但仍未取得實質性進展[2]。開展中華鱉性控育種,首先需要了解其性別差異的理論機制, 特別是在分子生物學層面上, 為此我們做了一些研究嘗試。在之前的工作中, 我們通過對中華鱉發育初期雌雄性腺轉錄組進行差異分析, 篩選到許多與性別相關的基因, 組蛋白H2A變體就是其中之一, 其相對表達量與中華鱉雌性性別顯著相關(數據未發表)。因此, 我們以組蛋白H2A變體為目標基因, 對其進行初步研究。
組蛋白H2A是染色質核小體的核心組蛋白之一(H3、H4、H2A和H2B), 是維持染色體結構和功能所必需的[3,4], 組蛋白H2A廣泛參與了轉錄調控、DNA修復和染色質濃縮等生物學過程[5—7]。此外,組蛋白H2A還擁有眾多變體, 其中包括H2A.X、H2A.Z、H2A.BbD和macroH2A等[8—10]。它們的差異主要在于C端的長度和序列的不同[11]。MacroH2A和H2A-Bbd僅限于脊椎動物或哺乳動物[12], MacroH2A主要富集在哺乳動物雌性中的非活性X染色體上[13], H2A Bbd則定位于活性X染色體和常染色體[12], 而H2A.X和H2A.Z則在整個基因組中組成性表達和定位[14]。在小鼠(Mus musculus)中, H2A.Z幾乎不存在于正在生長的卵母細胞核中(第5天卵母細胞除外), 但是在囊胚期的細胞核中可以檢測到熒光信號[11]。在裂殖酵母(Schizosaccharomyces pombe)中, H2A.Z還可以通過調節染色體結構, 以促進減數分裂中DNA雙鏈斷裂(DSB)的形成, 從而促進配子的發生[15]。H2A.X在小鼠整個卵子發生過程中和植入前的胚胎的核中都可檢測到其表達[11],同時敲除H2A.X可以導致雄性小鼠不育[16]。H2A.Bbd在小鼠胚胎原核期和2細胞期的細胞核中均顯示較弱的表達信號, 但從4細胞期到胚泡期的細胞核中可以檢測到強信號[11]。同時, 也有研究表明H2A.Bbd也存在于哺乳動物睪丸的高級生精組分和人類成熟精子的染色質中[17]。然而, 目前在龜鱉類動物中有關組蛋白H2A變體在性別分化中的功能尚未得到很好的解析。在這里, 我們克隆了中華鱉PsH2A的同源物, 分析其表達模式, 并對其轉錄本在卵巢組織中進行卵母細胞定位, 以研究其在卵母細胞發育成熟過程中的作用。
1 材料與方法
1.1 實驗動物的收集和處理
實驗于2019年1、4、7和10月份選取1、2和3冬齡健康的中華鱉, 雌雄各6只, 共144只, 全部來自于廣東惠州財興實業有限公司。將鱉麻醉后放血, 解剖, 掀開背甲, 然后采集精巢、卵巢、心臟、肝臟、脾臟、腎臟、腦和肌肉等組織, 速凍后,-80℃保存, 用于總RNA提取。另外, 再取一部分卵巢組織用4%多聚甲醛(Solarbio, China)固定, 4℃冰箱過夜。然后用甲醇/1×PBS(DEPC水配制)梯度脫水, 并置于100%甲醇中, -20℃保存, 用于制備切片。
1.2 RNA的提取與cDNA的合成
采集的組織使用Eastep?Super試劑盒(Promega, China)提取總RNA, 濃度和純度由NanoQTM分光光度計測定, RNA的完整性由1%瓊脂糖凝膠電泳檢測, 使用M-MLV反轉錄試劑盒(Invitrogen,USA)進行cDNA第一鏈的合成。
1.3 中華鱉PsH2A cDNA的克隆
根據測序所得的中華鱉PsH2AcDNA序列, 設計上下游特異性引物Ps-H2A-F和Ps-H2A-R (表 1)。以中華鱉卵巢cDNA為模板進行PCR擴增, 反應條件為: 94℃ 2min; 98℃ 30s, 55℃ 30s, 72℃ 90s (共35個循環); 72℃延伸10min。PCR產物使用Gel Extraction Kit (Omega, China)試劑盒回收。然后將目的片段連接到pMD19-T (TaKaRa, China)載體, 轉化至DH5α感受態細胞(TaKaRa, China)中, 克隆篩選,并將陽性克隆送廣州天一輝遠基因科技有限公司測序。DNAMAN軟件預測氨基酸序列, Clustal W(BioEdit)軟件進行氨基酸序列對比, 按MEGA 7軟件的鄰近法(Neighbour-Joining, NJ)構建系統進化樹。

表1 用于中華鱉PsH2A基因克隆和表達分析的引物Tab. 1 Primers used for cDNA cloning and expression analysis of PsH2A in P. sinensis
1.4 組織表達
根據測序所得的中華鱉PsH2AcDNA序列, 設計一對特異的定量引物PsDL-H2A-F和PsDL-H2AR, 并以中華鱉EF1α為內參基因(表 1)。以成體卵巢、精巢、心臟、肝臟、脾臟、腎臟、腦和肌肉的cDNA為模板, 進行RT-qPCR和RT-PCR擴增。RT-qPCR擴增反應在StepOnePlus real-time PCR System (Applied Biosystems)中完成, 每組3個平行重復, 采用2-ΔΔCt法來計算目的基因的相對表達量,反應程序如下: 95℃預變性10min; 95℃ 15s, 60℃15s, 72℃ 15s (共40個循環); 95℃ 15s, 60℃ 30s,95℃ 15s。RT-PCR使用與RT-qPCR相同的特異性引物(PsDL-H2A-F/PsDL-H2A-R, 表 1)進行擴增, 反應程序如下: 94℃ 2min; 98℃ 30s, 58℃ 30s, 72℃40s (共30個循環); 72℃延伸10min。作為對照, 從相同的cDNA樣本中擴增出內參基因(EF1α)。RTPCR產物由1%的瓊脂糖凝膠進行分離, 并拍照(Alpha Innotech, USA)。
1.5 探針的制備
根據測序所得的中華鱉PsH2AcDNA序列設計一對特異性引物PsCISH-H2A-F和PsCISH-H2AR(表 1), PCR擴增產物通過1%的瓊脂糖凝膠分離, 并用Gel Extraction Kit試劑盒(Omega, China)進行膠回收。將純化后的產物連接到pMD19-T(TaKaRa, China)載體中, 然后轉入DH5α感受態細胞(TaKaRa, China)中, 篩選陽性克隆, 提取質粒,送至天一輝遠生物技術有限公司進行測序。使用SP6/T7 Enzyme Mix (Roche, Germany)和Digoxigenin (DIG) RNA Labeling試劑盒(Roche, Germany)體外合成正反義探針。
1.6 冰凍切片的制備
將現取的卵巢組織, 置于4%的多聚甲醛(Solarbio, China)中, 4℃冰箱固定過夜。使用甲醇/1×PBS(DEPC水配制)梯度脫水, -20℃保存。待切片時, 使用甲醇/1×PBS(DEPC水配制)梯度復水, 并置于30%蔗糖溶液中, 4℃冰箱過夜, 具體步驟按張飄逸等人的方法進行[18]。待組織塊完全沉入管底后,取出, 用OCT(SAKURA, USA)包埋, Leica冷凍切片機(CM1950)切成4—6 μm厚的切片貼于防脫載玻片上。
1.7 化學原位雜交
化學原位雜交操作過程按照Xu等[19,20]的方法進行。采用BCIP/NBT(Roche, Germany)顯色液進行化學顯色, 50%甘油(PBS配置)封片, 使用尼康Ri2顯微鏡觀察并用NIS Elements成像系統拍照。根據中華鱉卵子發生過程中濾泡細胞及透明帶等結構特征, 對其進行分期和鑒定[21,22]。
1.8 數據分析
每個實驗獨立重復至少3次。所有實驗數據均以平均值±標準誤表示, 并使用SPSS軟件進行方差分析和Duncan氏多重比較,P<0.05表示差異顯著。
2 結果
2.1 中華鱉PsH2A cDNA序列特征
克隆獲得的中華鱉PsH2AcDNA (GenBank登錄號: MZ546415)序列全長為575 bp, 5′端非編碼區(5′-UTR) 68 bp, 3′端非編碼區(3′-UTR) 108 bp, 開放閱讀框(Open reading frame, ORF)為399 bp, 共編碼133個氨基酸, 具有H2A超家族特有的結構功能域,我們將其命名為PsH2A。氨基酸序列同源性比對結果顯示: 其與三趾箱龜(Terrapene carolina triunguis)和綠海龜(Chelonia mydas)的同源性最高, 分別為82.71%和80.45%; 與白喉帶鹀(Zonotrichia albicollis)和朱紅蜂鳥(Calypte anna)的同源性分別為56.92%和51.11%; 與非洲爪蛙(Xenopus tropicalis)和蚓螈(Microcaecilia unicolor)的同源性分別為52.38%和49.59%; 與斑馬魚(Danio rerio)和虹鱒(Oncorhynchus mykiss)的同源性分別為53.49%和54.26%; 與人(Homo sapiens)和褐家鼠(Rattus norvegicus)的同源性分別為52.71%和54.55%。NJ系統進化樹結果表明,PsH2A在進化關系上與龜類動物較近, 其次是鳥類、兩棲類和魚類, 與哺乳動物較遠(圖 1)。
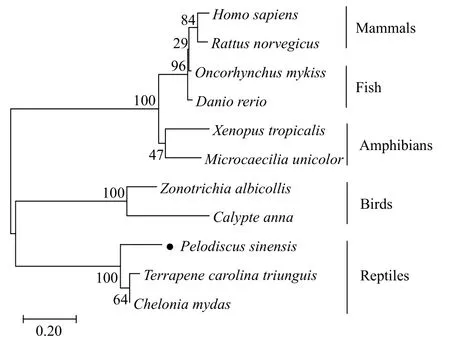
圖1 采用NJ法構建中華鱉PsH2A系統進化樹Fig. 1 Phylogenetic tree using the neighbor-joining method
2.2 中華鱉PsH2A mRNA組織表達
以EF1α為內參基因, 利用RT-qPCR和RTPCR檢測PsH2A轉錄本在4月份, 1冬、2冬和3冬齡中華鱉各組織中的相對表達。結果顯示, 在1冬、2冬和3冬齡中華鱉個體中,PsH2AmRNA在卵巢中具有較高水平的表達(P<0.01), 而在精巢、肌肉、腦、腎臟、脾臟、肝臟和心臟等體組織中幾乎檢測不到, 提示中華鱉PsH2AmRNA的表達具有組織特異性和性別二態性。PsH2AmRNA在1冬齡到3冬齡中華鱉雌雄性腺的表達見圖 2。PsH2AmRNA在3冬齡中華鱉體組織中的表達見圖 3。
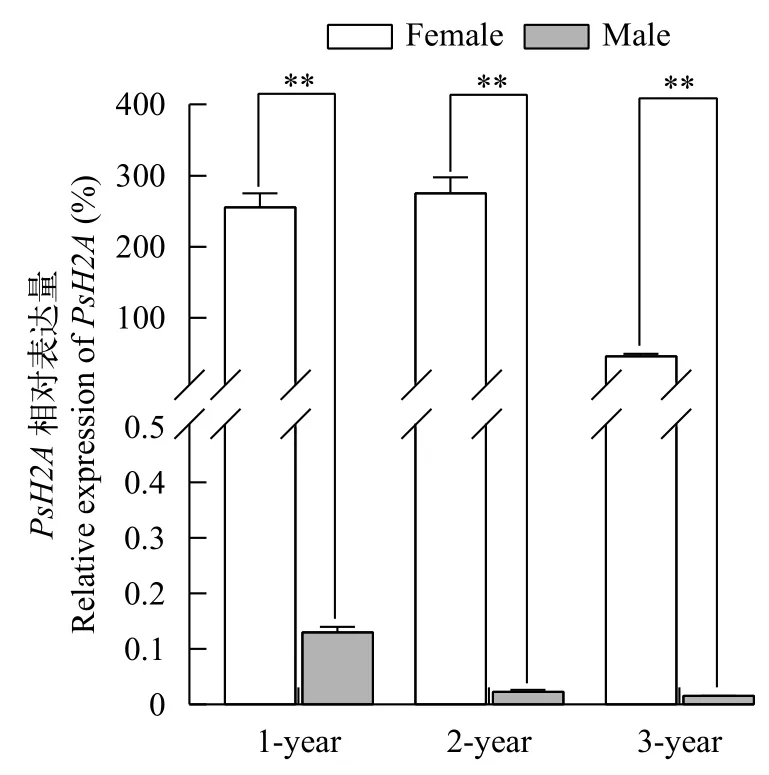
圖2 中華鱉PsH2A mRNA在4月份不同冬齡雌雄性腺中的相對表達Fig. 2 The relative expression of PsH2A mRNA in female and male gonads of different ages in April
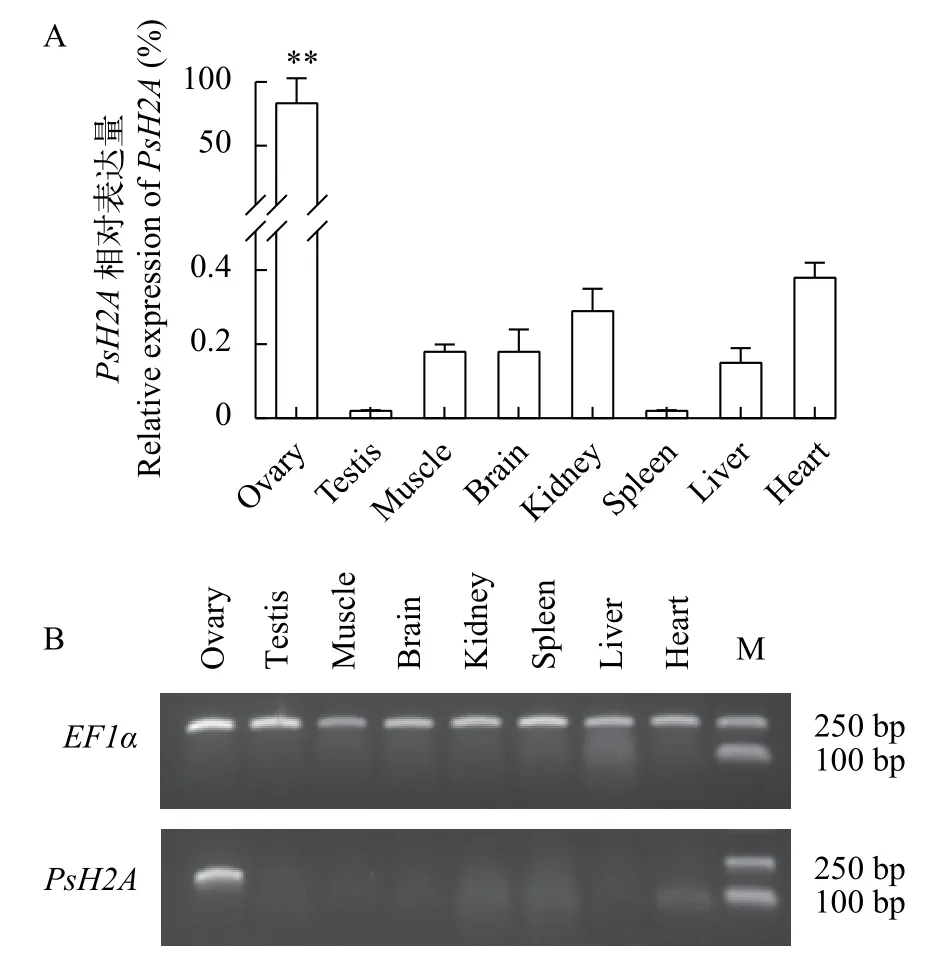
圖3 中華鱉PsH2A mRNA在3冬齡成體組織中的特異性表達Fig. 3 Specific expression analysis of PsH2A mRNA in 3-yearold adult tissues
2.3 中華鱉PsH2A mRNA在卵母細胞發生過程中的表達分布
1冬齡中華鱉卵巢發育至初級卵泡期, 初級卵母細胞核位于細胞中央, 部分細胞核由于染色質空泡的出現被擠壓到細胞邊緣; 2冬齡中華鱉卵巢進入生長期, 卵巢內具有各個時期的卵泡, 少部分卵泡進入成熟期, 卵黃顆粒在卵的周圍開始積累; 3冬齡中華鱉卵巢已經進入成熟期, 輸卵管附近可見石灰質殼的卵[23,24]。我們選取2冬齡中華鱉卵巢切片,對PsH2A進行細胞定位, 化學原位雜交結果顯示:PsH2A轉錄本通過反義探針顯示化學信號(圖 4A和4B), 而正義探針檢測不到信號(圖 4C和4D)。PsH2AmRNA在卵母細胞中特異性表達, 并且在初期卵母細胞發育至第Ⅲ期的時候, 表達信號最強,且均勻地分布在卵母細胞質中。在初級卵母細胞發育至第Ⅴ期的時候, 目的信號減弱, 信號從細胞核周圍至細胞質外圍逐漸遞減。隨著發育進程, 卵母細胞逐漸增大, 進入生長期和成熟期, 目的信號逐漸減弱。在生長期(第Ⅶ期)卵母細胞中, 目的信號呈片狀分布在細胞核周圍區域, 信號強度較第Ⅴ期的卵母細胞更弱, 細胞質外圍幾乎很難檢測目的信號(圖 4A)。在成熟期(第Ⅸ期)卵母細胞中, 目的信號在細胞核周圍呈斑點狀分布, 而在細胞質外圍已經檢測不到目的信號(圖 4B)。PsH2A的這種特殊表達模式, 提示其在中華鱉卵母細胞發育過程中的重要作用。
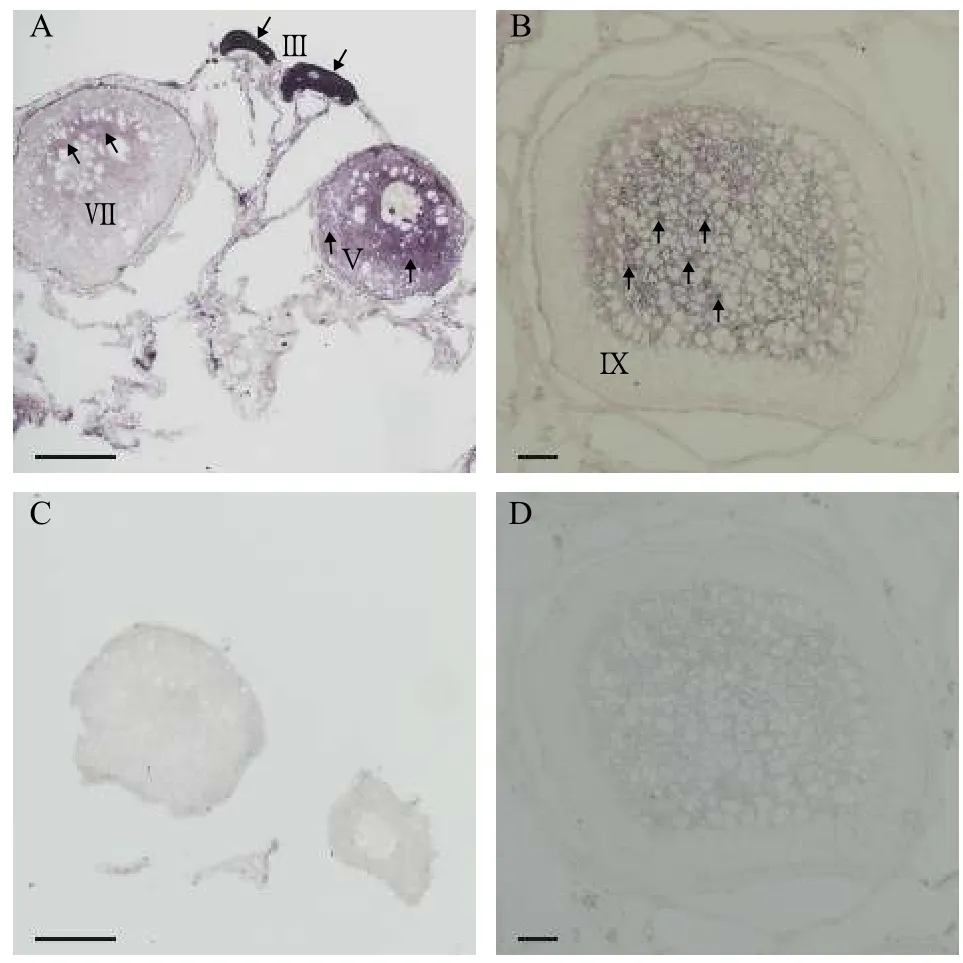
圖4 通過化學原位雜交分析中華鱉PsH2A mRNA在卵巢中的表達Fig. 4 The PsH2A mRNA expression in ovary by chemical in situ hybridization
2.4 中華鱉PsH2A mRNA季節表達模式
中華鱉屬于季節性繁殖動物, 我國南方養殖的中華鱉一般4—8月為產卵季節, 5—7月為產卵高峰期, 11月開始冬眠, 并將交配后的精子儲存在輸卵管中, 供來年受精使用[25,26]。用RT-qPCR檢測中華鱉卵巢組織中PsH2A不同季節的表達水平變化, 結果顯示: 1冬齡個體卵巢PsH2A轉錄本的相對表達量先上升后下降再上升, 10月的表達量最高; 2冬齡個體PsH2A轉錄本的相對表達量先上升后下降, 在4月達到峰值; 3冬齡個體PsH2A轉錄本的相對表達量隨著月份整體呈上升趨勢。此外, 在相同月份中,隨著中華鱉年齡的增長,PsH2A轉錄本的相對表達量整體呈現下降趨勢, 即PsH2A表達量與年齡增長呈負相關(圖 5)。
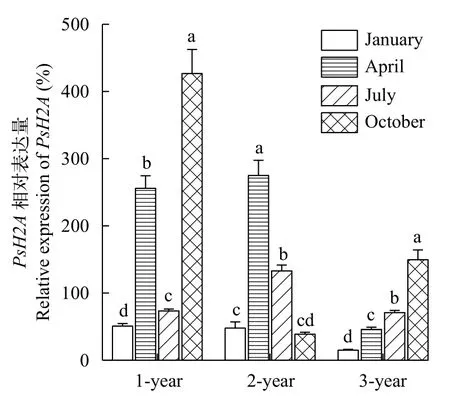
圖5 中華鱉PsH2A mRNA在不同季度和不同冬齡卵巢中的相對表達Fig. 5 The Relative expression of P. sinensis PsH2A mRNA in ovary of different seasons and ages
3 討論
卵母細胞發育過程中的重要事件之一是染色質重塑, 其中就包括組蛋白的修飾和交換[27]。本研究克隆的中華鱉PsH2AcDNA序列全長為575 bp,編碼133個氨基酸, 具有H2A超家族特有的結構功能域。在進化上, 與龜類親緣關系最近。使用RT-qPCR和RT-PCR技術, 對中華鱉PsH2A組織差異性表達情況進行分析, 發現PsH2AmRNA在卵巢組織中高表達, 而精巢和其他組織幾乎不表達, 表明PsH2A的表達在中華鱉中具有性別二態性和組織特異性。這與魚類中發現的表達模式類似, 在異育銀鯽(Carassius auratus gibelio)中,h2af1o轉錄本僅在成體卵巢中被檢測到。在蛋白水平上, 也僅在彩鯽(Carassius auratus)和斑馬魚的卵巢中檢測到類似的蛋白帶[28]。
為了進一步確定PsH2A在中華鱉卵巢組織中的細胞定位, 我們對其轉錄本在卵巢切片中進行了原位雜交實驗。結果發現PsH2AmRNA在卵母細胞中特異性表達, 在初級卵母細胞中表達信號最強,且均勻的分布在卵母細胞質中, 此時卵母細胞發育到第Ⅲ期, 卵母細胞中燈刷染色體明顯[21], 卵母細胞進行第一次減數分裂并停留在雙線期, 染色體加倍。隨著卵母細胞發育成熟, 細胞增大, 目的信號逐漸減弱, 主要分布在核周區域, 此時卵母細胞已經進入生長期(第Ⅶ期)和成熟期(第Ⅸ期), 已觀察不到燈刷染色體的存在[21], 完成第一次減數分裂。中華鱉PsH2AmRNA在卵母細胞不同發育階段中的表達模式似乎與減數分裂中染色體的動態變化相契合。在哺乳動物中, 組蛋白H2A的翻譯后修飾和組蛋白H2A變體與染色質的結合參與了卵母細胞染色質的重塑, 對雌性配子的減數分裂和發育能力是至關重要的[29]。PsH2A這種特殊表達模式提示其在中華鱉卵母細胞發育和成熟中可能發揮著重要作用。中華鱉PsH2AmRNA在卵母細胞發育過程中的表達模式與魚類相似[28], 而與小鼠[11]和牛(Bos indicus)[30]有所不同。在異育銀鯽中,H2A變體h2af1o在整個卵子發生期間特異性表達, 在卵原細胞和初級卵母細胞中,h2af1o轉錄本信號均勻地分布在細胞質中, 從皮質肺泡卵母細胞到成熟卵母細胞, 轉錄信號主要集中在細胞質外周, 呈斑點式分布, 而在體細胞中檢測不到信號。在牛的卵母細胞中,H2A轉錄本的表達隨著卵泡的增大而逐漸增加, 暗示其對于卵母細胞的發育至關重要。Osakabe等[11]認為, 組蛋白H2A與其變異體在C端上存在差異, C端的異質性決定了其功能的不同, 解釋了組蛋白H2A及其變異體在不同物種的差異表達的可能原因。
不同年齡中華鱉卵巢組織中PsH2AmRNA的相對表達量, 隨著年齡增長, 呈現下降趨勢。此外,同一年齡中華鱉在不同季節中, 卵巢組織中PsH2AmRNA的水平也存在顯著差異。這可能與中華鱉性成熟過程中卵巢的發育程度有關。1冬齡的中華鱉沒有性成熟, 卵巢組織中主要是初級卵母細胞[24],PsH2AmRNA的相對表達量整體呈上升趨勢; 2冬齡中華鱉開始性成熟, 卵巢進入生長期, 卵巢內具有各個時期的卵泡, 少部分卵泡進入成熟期[23],PsH2AmRNA在4月和7月高表達, 可能與氣溫逐漸升高, 卵子發生比較活躍有關; 3冬齡中華鱉完全性成熟, 其卵巢已經進入成熟期, 輸卵管附近可見石灰質殼的卵[23],PsH2A的轉錄水平隨月份逐漸升高,可能因為卵巢已進入成熟期, 卵巢一直經歷著由卵原細胞到成熟卵子的周期性過程。這也與PsH2AmRNA在卵母細胞發育過程中的原位雜交結果相一致。中華鱉PsH2AmRNA主要在初級卵母細胞中表達, 隨著卵母細胞發育成熟, 表達信號逐漸減弱。中華鱉PsH2AmRNA的相對表達與卵子發育的成熟程度呈負相關, 因此可以在生產實踐中將其作為評估中華鱉卵巢成熟程度和高繁殖力群體的一個潛在指標。總之, 本研究提示PsH2A在卵母細胞發育成熟過程中起著重要作用, 同時也為今后研究龜鱉類動物生殖細胞發育調控機制拓展了新思路。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
