制動應激對孕豬海馬結構神經元可塑性的影響
2021-12-31 01:39:50陳思彥孫維婧王子旭陳耀星董玉蘭
畜牧獸醫學報 2021年12期
關鍵詞:海馬
陳思彥,孫維婧,王子旭,陳耀星,曹 靜,董玉蘭
(中國農業大學動物醫學院,北京 100193)
在我國的生豬規模化養殖中,妊娠期母豬常使用限位欄養殖。雖然這種養殖方法能夠節省養殖空間、提高養殖效率、方便孕期管理、疫苗免疫和精準飼喂,但也使孕豬長期處于制動應激狀態,這種應激不僅不符合動物福利的要求,還會造成母豬生理和心理上的雙重損傷,影響母豬的激素分泌、神經營養和生育繁殖等機能,甚至對子代健康產生負面作用,最終導致母豬的使用年限下降,淘汰率升高,給豬場造成經濟損失。因此,如何正確評估限位欄養殖對母豬造成的影響,降低養殖風險,提高后備母豬的繁殖力也逐漸引起學者和養殖界的關注。同時,妊娠模型動物豬應激反應的研究對人類妊娠期應激導致疾病的機理研究和防治同樣具有啟發意義。下丘腦-垂體-腎上腺軸(HPA軸)是機體神經內分泌系統的重要組成部分。當應激源刺激機體時,信號從大腦皮層傳入邊緣系統,作用于下丘腦,在HPA軸的調控下,最終釋放大量糖皮質激素(glucocorticoid,GC)。應激時HPA軸的功能是由海馬、下丘腦室旁核(paraventricular nucleus of hypothalamus,PVN)和杏仁核共同調節的,GC又可反饋作用于海馬的糖皮質激素受體(GR)和鹽皮質激素受體(MR)引起一系列反應。應激能夠引起海馬形態和功能的改變。大量證據表明[1-2],海馬是應激損傷的敏感區[3],應激會對海馬突觸可塑性、神經元存活、海馬神經發生、海馬神經元細胞代謝和機體記憶力等各個方面造成影響。可塑性被認為是神經系統應對內、外部刺激時產生的適應性結構重組以改變功能的能力[4-10]。大量資料顯示,在應激改變海馬形態和功能的過程中,腦源性神經營養因子(brain-derived neurotrophic factor,BDNF)可能充當了重要的角色。海馬中BDNF的表達水平在急性或慢性的應激中均有敏感的變化[11-13],而BDNF的減少也是影響海馬神經元的發生[14-15]、導致海馬神經元的退行性變化和突觸的可塑性變化的一個因素[16-17]。而且,海馬神經元中BDNF/TrkB由5-HT2A受體介導、雌激素參與的信號傳導的調控[18],其下突觸級聯反應涉及突觸可塑性[19],提示BDNF是妊娠母豬應激反應時影響突觸可塑性且可監測的重要指標蛋白。本研究選用8~9月齡的巴馬小型母豬,將配種成功的母豬隨機分為應激組(n=3)和對照組(n=3),應激組從妊娠第1天開始飼養于定制的母豬限位欄中,模擬實際生產中孕豬所承受的制動應激環境,對照組在寬敞自由的環境中飼養,其他飼養條件相同。妊娠第18天處死動物,采集孕豬血液,進行相關激素水平的檢測;分離孕豬的海馬組織,檢測海馬BDNF表達水平,并通過尼氏染色和鍍銀染色檢測海馬的神經元丟失率、樹突復雜度以及樹突棘數量等多個指標,來研究心理應激在胚胎著床期對孕豬造成的類固醇激素合成功能的變化及對海馬神經元可塑性的影響。
1 材料與方法
1.1 材料
1.1.1 實驗動物及處理 于北京實創世紀小型豬養殖基地購入6頭雌性巴馬小型豬作為實驗動物,所有母豬月齡相近(8~9月齡)、體重相似(20~30 kg)且健康無病。將其均飼養于標準環境下(溫度為22 ℃;12 h/12 h光照制度;每日上午8:00開燈,下午8:00關燈)并自由采食飲水。將實驗母豬成功配種后隨機分為兩組,每組3頭:1)對照組:將每頭妊娠巴馬小型豬單獨飼養于干凈、寬闊的環境下,可自由活動;2)制動應激組:將每頭妊娠巴馬小型豬單獨飼養于50 cm×60 cm×100 cm的金屬籠中,孕豬只可前后小范圍運動,無法左右轉頭。
在持續制動應激處理的第18天上午對實驗動物麻醉后心臟放血處死。用開顱器取出完整豬腦,于延髓后切斷,從正中矢狀面將大腦一分為二,左半球分離出各腦部結構并置于液氮凍存,右半球完整置于4%多聚甲醛固定。
血漿獲取:制動應激處理前1 d記為D0(發情期),試驗期間分別于D0和D18采集孕豬頸靜脈血樣。
1.1.2 主要試劑和設備 兔單克隆抗體BDNF(ab108319,abcam,美國),RIPA裂解液、蛋白酶抑制劑和HRP標記山羊抗兔IgG(CW0211)均購自江蘇康為世紀生物科技有限公司。豬促腎上腺皮質激素釋放激素(CRH)ELISA 試劑盒(CEA835Po)和豬5-羥色胺(5-HT)ELISA檢測試劑盒(CEA808Ge)均購自武漢云克隆科技股份有限公司。YP1200電子天平購自上海第二天平儀器廠,生物組織包埋機購自浙江科迪儀器設備有限公司,高速臺式離心機購自長沙湘儀離心儀器有限公司,酶標儀購自美國BLO-RAD,多管放射免疫計數器購自合肥眾成機電技術公司,轉膜電泳槽購自北京六一儀器廠,電泳儀購自上海伯樂生命醫學產品有限公司。
1.2 方法
1.2.1 血液激素含量的測定 檢測血漿中促腎上腺皮質激素釋放激素(corticotropin-releasing hormone,CRH)的濃度:按照ELISA試劑盒說明書操作檢測血漿中CRH的濃度。
檢測血漿中的促腎上腺皮質激素(adrenocorticotropic hormone,ACTH)和皮質醇(cortisol,COR)濃度:使用放射免疫法。將血漿樣品解凍,分別加樣,混勻后室溫放置15 min,3 500 g轉速離心15 min。棄上清,檢測各管沉淀的放射性計數并采用“相對結合率-對數計量的分數對數”的數學模型進行擬合。
1.2.2 尼氏染色 海馬在4%多聚甲醛中固定,經過修塊、梯度脫水、透明浸蠟,切片每張厚度15 μm。將石蠟切片脫蠟并在酒精下行水化,進行尼氏染色,將切片置于尼氏染液中染色10 min,在蒸餾水中輕洗5 s,然后放入95%酒精中分色,分色程序視染色效果而定,一般為95%酒精Ⅰ 1 min、95%酒精Ⅱ 1 min、95%酒精Ⅲ 1 min,再酒精上行脫水,入二甲苯,最后中性樹膠封片,置于光學顯微鏡下觀察。視野可見尼氏體呈深藍色,細胞核呈淺藍色,背景為無色或者淡藍色。
神經元密度評價方法:根據神經元形態和神經纖維排列的不同,海馬本部被分為CA1、CA2、CA3和CA4 4個區。從海馬切片中隨機選取6張相同定位切片,在放大400倍條件下,從CA1區和CA3區含有完整結構的神經元區域開始,連續采集3個不重疊的視野,計數每個視野內結構完整的神經元數量,求均值以代表正常形態神經元密度[20]。
1.2.3 鍍銀染色 將新鮮取材的豬腦半球放入4%多聚甲醛中固定一周以上,然后使用1.5%重鉻酸鉀鉻化,3%的硝酸銀溶液浸泡鍍銀48 h,脫水、包埋、切片、封片后即可進行觀察。
樹突測量標準。在孕豬海馬CA3區選取具有代表性的錐體細胞,在齒狀回(dentate gyrus,DG)選擇獨立的與周圍神經元交叉較少的顆粒細胞進行圖像采集、圖像處理和分析,神經元采集標準:1)選取位于切片厚度中部的錐體細胞進行統計,盡量避免樹突的缺失;2)盡量選取相對獨立的,與周圍的神經元樹突有較少交叉的神經元進行分析。照片使用Olympus成像系統在20倍物鏡下連續采集圖像,通過均勻微調細準焦螺旋在切片不同焦距采集連續的神經元的圖像,每個視野采集6張圖像。神經元的二維重建采用Neuronstudio軟件,圖像的測量分析使用Image-J win64軟件,數據統計分析使用SPSS軟件。數據描述為“平均值±標準差”,數據比較P≤0.05 視為有顯著差異,Graphpad軟件作圖[21]。
Sholl分析法是一種對神經元樹突復雜度進行定量分析的經典方法,可用于確定樹突形態。從神經元胞體開始,每隔相同距離畫一個圈覆蓋樹突,由同心圓與樹突交叉點的數量來評估神經元樹突的復雜性,神經元樹突追蹤可獲得神經元樹突分支點的數量以及神經元樹突的總長度。
樹突棘測量標準。樹突棘是神經元樹突上的微小突起,是和其他神經元的軸突末梢形成突觸的結構。樹突棘的主要形狀包括瘦長型、短粗型、蘑菇形和絲狀偽足。每張切片選擇3個處于切片正中、樹突盡量完整的神經元參與統計,每個神經元選取5根只有一個分支點的二級樹突參與統計,在100倍油鏡下選取和背景無交叉的、總體處于同一焦距的、平行于鏡頭的樹突,通過均勻微調細準焦螺旋在切片不同焦距采集連續的樹突圖像,使用Photoshop合成一個較為清晰的視野,每個視野選取相對直的長20 μm的樹突節段進行計數,樹突棘密度即為視野內總樹突棘數目除以樹突分支長度。
1.2.4 Western blot 新鮮組織提取蛋白。用RIPA倍比稀釋蛋白樣品并使用酶標儀測量吸光光度值,做出標準曲線得到蛋白樣品濃度后用RIPA將蛋白樣品濃度調平。將未變性蛋白與Loading buffer混合煮樣使之變性后,進行電泳,然后冰浴轉膜,轉膜完成后將PVDF膜放入脫脂奶中封閉,用一抗(BDNF)孵育過夜,用TBST洗滌后再放入HRP標記的羊抗兔二抗中,室溫搖床孵育2 h,之后進行熒光顯色,最后使用全自動數碼凝膠圖像分析系統進行圖像采集。目的蛋白和內參蛋白的面積和光密度使用ImageJ軟件統計。目的蛋白表達量=(目的蛋白面積×光密度)/(內參蛋白面積×光密度)。
1.3 數據統計
使用軟件 SPSS 23.0(SPSS Sciences,Chicago,USA)進行數據分析。采用獨立樣本T檢驗分析對照組與應激組的差異顯著性。試驗數據采用“平均值± 標準誤”,P≤0.05為差異顯著。
2 結 果
2.1 孕豬血漿中應激相關激素的水平
在制動應激處理18 d后,應激組血漿CRH水平較應激前上升221.65%(t=2.71,P=0.046),較同時間的對照組顯著上升296.36%(t=3.026,P=0.038,圖1A)。由放射免疫法測得,在第18天時應激組血漿ACTH水平較應激前上升64.97%(t=5.151,P=0.007),較同時間的對照組高出62.98%(t=4.549,P=0.010,圖1B);而COR水平較應激前上升97.76%(t=5.131,P=0.007),較同期對照組顯著高出 109.86%(t=5.602,P=0.005,圖1C)。制動應激處理后,孕豬血漿中的各級應激激素水平均顯著上升,表明成功建立孕豬制動應激模型。
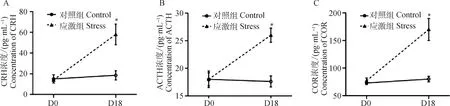
D0 表示制動應激開始前1 d,D18 表示制動應激第18天。*表示制動應激組與相同日齡的對照組具有顯著性差異(P≤0.05)D0 represents the day before the restraint stress treatment,and D18 represents the 18th day of restraint stress treatment.* indicates that the difference is significant between the control group and the stress group (P≤0.05)圖1 制動應激對孕豬血漿CRH、ACTH和COR水平的影響Fig.1 The effect of restraint stress on plasma CRH,ACTH and COR level in pregnant sows
2.2 制動應激對孕豬海馬組織結構的影響
尼氏染色法是一種通過堿性染料染色神經組織的方法,本研究使用尼氏染色來對神經元進行計數,觀察是否有神經元丟失的情況發生。將孕豬海馬石蠟切片進行尼氏染色后,在DG的顆粒細胞層中,對照組的顆粒細胞整體染色均勻,胞核大且呈卵形,細胞排列緊密;應激組中,顆粒細胞胞核排列整體松散,胞核染色不均且整體呈錐形,顆粒細胞內側的膠質細胞和多形細胞的細胞形態基本一致,無明顯可見差異(圖2A)。而在海馬CA1區和CA3區中,對照組錐體細胞層的錐體細胞核大而圓,頂樹突長而直,投向分子層,其中CA3區整體排列整齊緊密,CA1區較為松散,尼氏體染色均勻、無凝集、無分散;應激組海馬的CA3區部分錐體細胞出現空泡狀,排列松散,肉眼可見部分神經元位置空缺,而在應激組CA1區的錐體細胞中可見尼氏小體分布減少,染色較淺。
通過神經元計數分析發現,應激組CA1區中的錐體細胞數量和對照組無顯著差異(P=0.221 9,圖2B),應激組海馬CA3區的錐體細胞層顯著低于對照組11.3%(t=2.318,P=0.034,圖2C),應激組齒狀回中顆粒細胞數量較對照組也下降31.7%(t=3.407,P=0.003,圖2D),故在應激組的CA3區和DG均發生了神經元丟失的情況。

A.海馬組織學結構,圖中黑色箭頭指示齒狀回中的顆粒細胞層,紅色箭頭指示 CA1 區和 CA3 區中的錐形細胞層,圖中比例尺=100 μm;B.CA1 區錐體細胞層;C.CA3 區錐體細胞層;D.齒狀回顆粒細胞層,*表示對照組和應激組差異顯著(P≤0.05)A.Hippocampal stracture,the black arrow in the figure indicates the granulosa cell layer in the dentate gyrus,and the red arrow indicates the pyramidal cell layer in the CA1 and CA3 regions,scale bar =100 μm;B.Pyramidal cell layer in CA1 regions;C.Pyramidal cell layer in CA1 regions;D.Granule cell layer in DG,* indicates that the difference is significant between the control group and the stress group (P≤0.05)圖2 制動應激對孕豬海馬結構的影響及孕豬海馬尼氏染色的結果統計Fig.2 The effect of restraint stress on Nissl staining of hippocampus in pregnant sow and statistics of Nissl staining results in pregnant sows
2.3 制動應激對孕豬海馬神經可塑性的影響
2.3.1 制動應激對孕豬海馬CA3區神經元復雜度的影響 鍍銀染色可顯示出神經元的胞體、樹突、軸突以及更微小的樹突棘結構。為了評估制動應激對妊娠母豬大腦海馬CA3區(圖3A)神經元可塑性的影響,對孕豬海馬切片進行鍍銀染色(圖3B),利用ImageJ軟件對神經元的樹突追蹤后采取Sholl分析,結果表明:在孕豬海馬CA3區中,應激組中海馬錐體細胞神經元與同心圓交點的數量較對照組下降(圖3C)。應激組和對照組CA3區中的錐體細胞頂樹突和基樹突的樹突平均長度均無顯著差異(P=0.847 6和P=0.938 5,圖3D)。應激組CA3區錐體細胞的頂樹突終末點的數量較對照組減少61.3%(t=3.245,P=0.005,圖3E),基樹突終末點的數量較對照組減少46.1%(t=2.542,P=0.02,圖3E)。故制動應激后,孕豬海馬CA3區的錐體神經元樹突復雜度較對照組顯著下降。
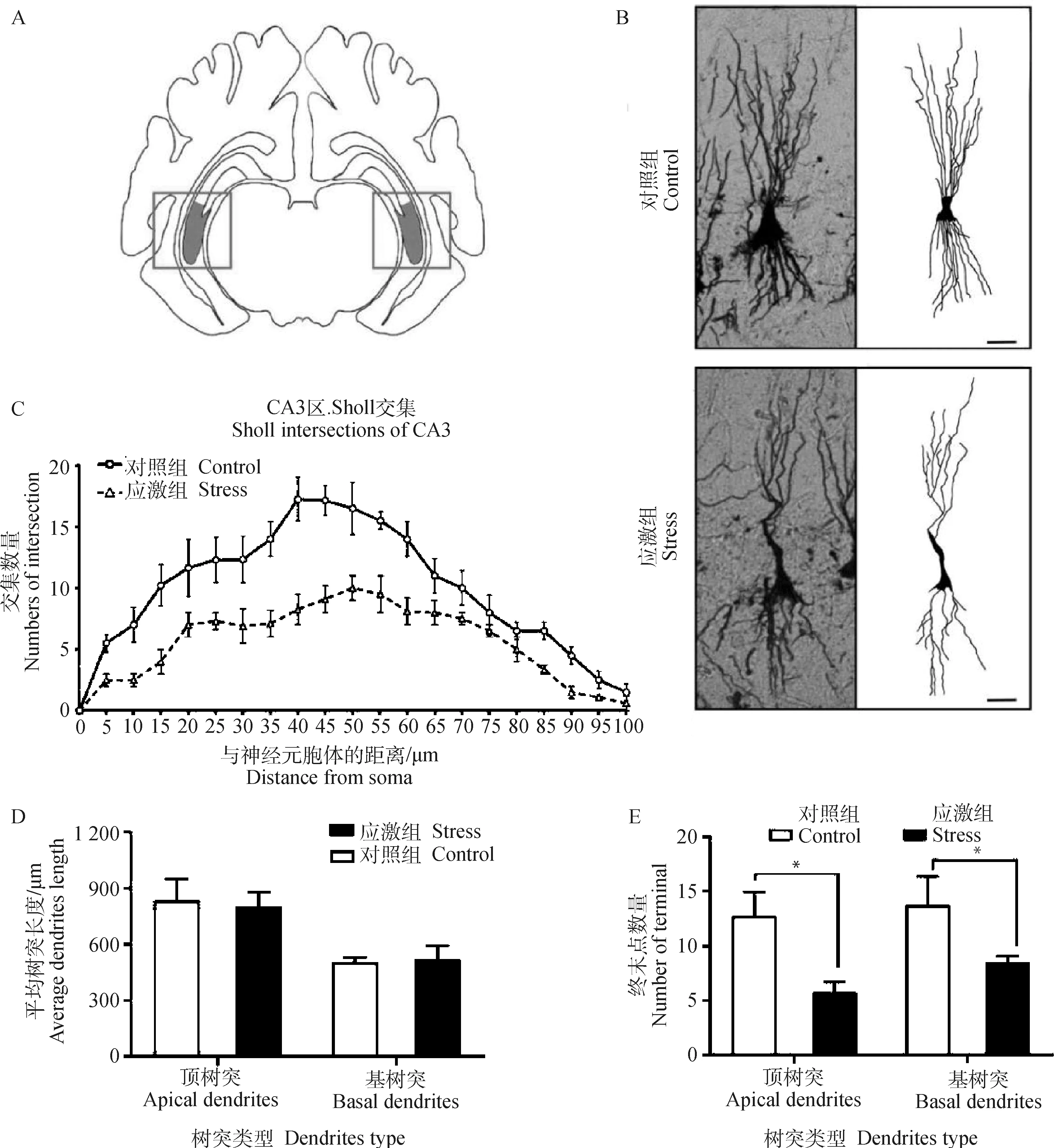
A.成年豬海馬 CA3 區示意圖;B.CA3 區錐體細胞的鍍銀染色及樹突追蹤結果,圖中比例尺為 20 μm;C.CA3 區樹突 Sholl 交叉點分析;D.CA3 區平均樹突長度;E.CA3 區樹突終末點數量。*表示對照組和應激組差異顯著(P≤0.05)A.Schematic diagram of hippocampal CA3 area of grown-up pig;B.Golgi’s staining and dendrite tracing results of pyramidal cells in CA3 region,scale bar=20 μm;C.Sholl intersection of CA3 region;D.Average dendritic length in CA3 region;E.Dendrite terminal number in CA3 region.* indicates that the difference is significant between the control group and the stress group (P≤0.05)圖3 制動應激對孕豬海馬 CA3 區神經元復雜度的影響Fig.3 The effect of restraint stress on neuronal dendrites complexity in hippocampus CA3 region of pregnant sow
2.3.2 制動應激對孕豬DG神經元復雜度的影響 海馬中新生神經元發生在DG(圖4A)的亞顆粒區,這些神經元在發育成熟后會整合到現有的神經網絡中。錐體細胞的鍍銀染色及樹突追蹤結果見圖4B。Sholl分析結果顯示,應激組妊娠母豬海馬DG中顆粒細胞的神經元與同心圓交點的數量較對照組無顯著變化(圖4C)。應激組DG的樹突平均長度(P=0.644)和顆粒細胞樹突終末點的數量(P=0.786)也較對照組無顯著性差異(圖4D和E)。故在18 d制動應激后,孕豬海馬DG中顆粒細胞的樹突復雜度無明顯變化。
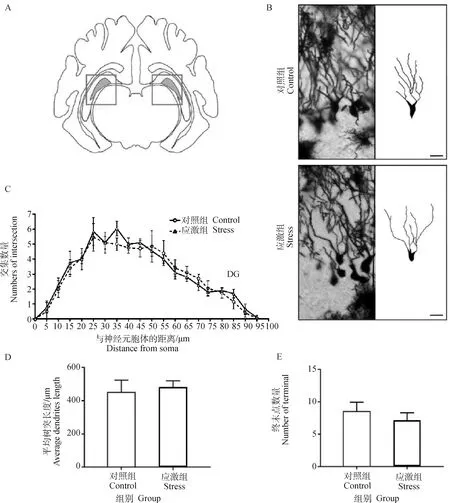
A.成年豬海馬 DG 示意圖;B.DG 錐體細胞的鍍銀染色及樹突追蹤結果,圖中比例尺為 20 μm;C.DG樹突 Sholl 交叉點分析;D.DG 平均樹突長度;E.DG 樹突終末點數量。*表示對照組和應激組差異顯著(P≤0.05)A.Schematic diagram of hippocampal DG area of grown-up pig;B.Golgi’s staining and dendrite tracing results of pyramidal cells in DG region,scale bar=20 μm;C.Sholl intersection of DG region;D.Average dendritic length in DG region;E.Dendrite terminal number in DG region.* indicates that the difference is significant between the control group and the stress group (P≤0.05)圖4 制動應激對孕豬海馬DG神經元復雜度的影響Fig.4 The effect of restraint stress on neuronal dendrites complexity in hippocampus CA3 region of pregnant sow
2.3.3 制動應激對孕豬海馬CA3區和DG神經元樹突棘的影響 鍍銀染色能顯示出神經元樹突上的細小突起,這種突起就是承載著哺乳動物中樞神經系統中大多數興奮性突觸功能的樹突棘,樹突棘根據其形態通常被分為4類:蘑菇型樹突棘(mushroom spines)、細樹突棘(thin spines)、短粗樹突棘(stubby spines)和絲狀偽足(filopodia),見圖5A。樹突、樹突棘形態等見圖5B、C。一般絲狀偽足和細樹突棘無法十分準確地區分計數,故將絲狀偽足也計入未成熟樹突棘中。樹突棘統計結果表明(圖5D~G):在18 d制動應激后,孕豬海馬CA3區的錐體細胞樹突上的樹突棘總數較對照組降低17.8%(t=2.115,P=0.046,圖5E)。DG中顆粒細胞樹突上的樹突棘總體數量較對照組降低19.3%(t=2.701,P=0.013,圖5G)。而將未成熟樹突棘和成熟樹突棘分開統計比較后發現,應激組CA3區未成熟樹突棘數量下降17.9%(t=3.099,P=0.01,圖5D),成熟樹突棘數量下降31.28%(t=2.255,P=0.024,圖5D)。DG的成熟樹突棘數量下降26.8%(t=2.463,P=0.022,圖5F),未成熟樹突棘數量變化不顯著(P=0.50,圖5F)。綜上所述,18 d的制動應激降低孕豬海馬CA3區和DG中的新生樹突棘的形成,同時增加CA3區中成熟樹突棘的消除,共同造成應激組海馬CA3區和DG中神經元樹突棘密度的減少。
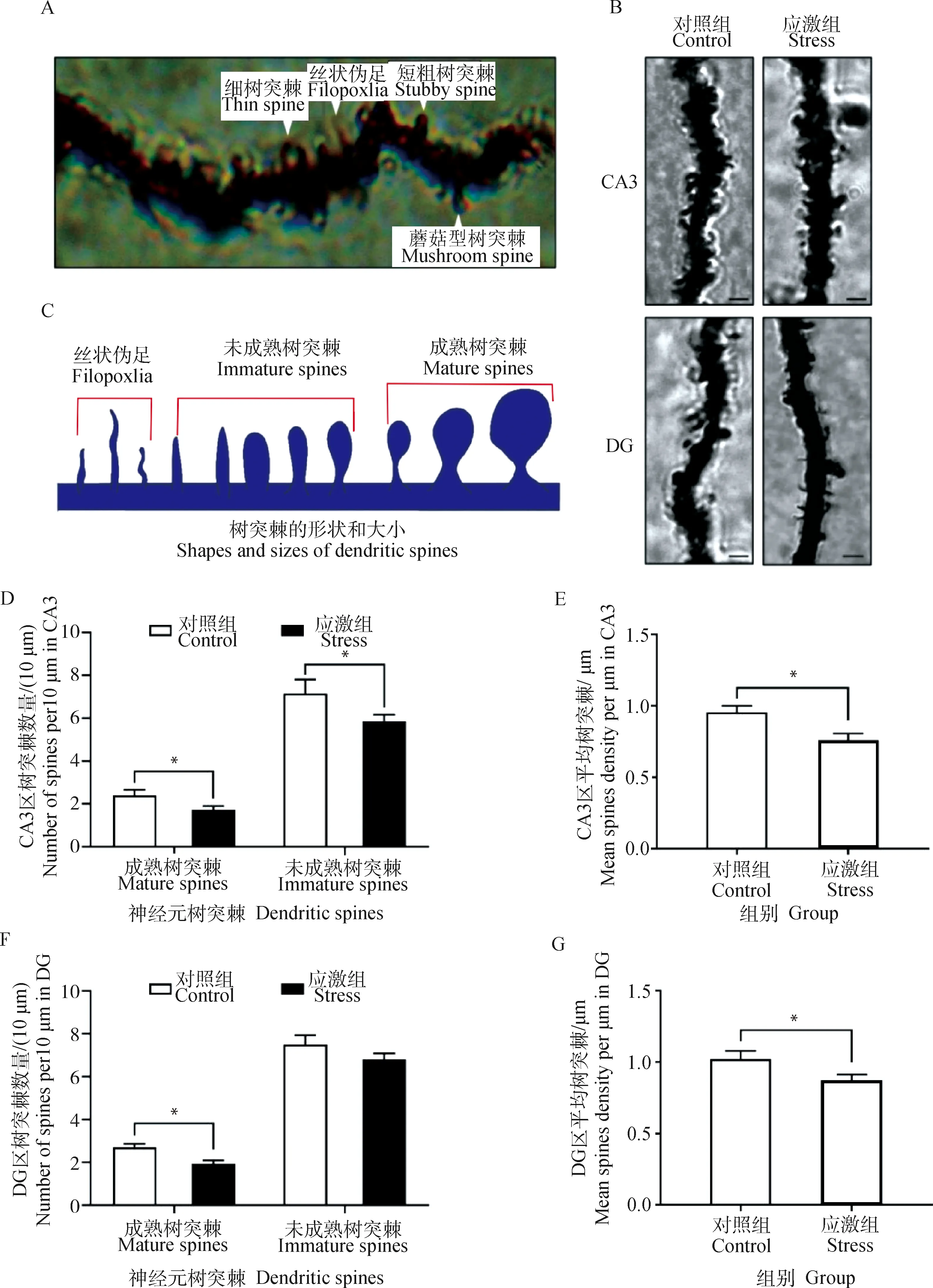
A.樹突棘的典型形態,包括細樹突棘、絲狀偽足、短粗樹突棘和蘑菇型樹突棘;B.應激組和對照組的樹突形態;C.樹突棘的形狀和大小,圖中包括絲狀偽足、未成熟樹突和成熟樹突;D~G.樹突棘密度(即視野內總樹突棘數除以樹突分支長度)A.Four typical dendritic spines morphology,including thin spines,filopodia,stubby spines and mushroom spines;B.Dendrite morphology in the stress group and the control group;C.shapes and sizes of dendritic spines,including filopodia,immature spines and mature spines;D-G.Dendrite spine density(Number of dendritic spines in the visual field divided by the length of dendritic branches)圖5 制動應激對孕豬CA3區和DG神經元樹突棘的影響Fig.5 The effect of restraint stress on neuronal dendrites spines in hippocampus CA3 region and DG of pregnant sow
2.4 制動應激對孕豬海馬BDNF蛋白表達水平的影響
BDNF是神經元和神經膠質細胞的中央神經營養因子(central neurotrophic factor,NTF)[22],在海馬中高表達[23-24],與海馬相關的眾多神經損傷性疾病有關[25]。Western blot結果顯示,應激組中孕豬海馬內的BDNF蛋白表達量較對照組降低70.6%(t=6.642,P=0.00,圖6)。
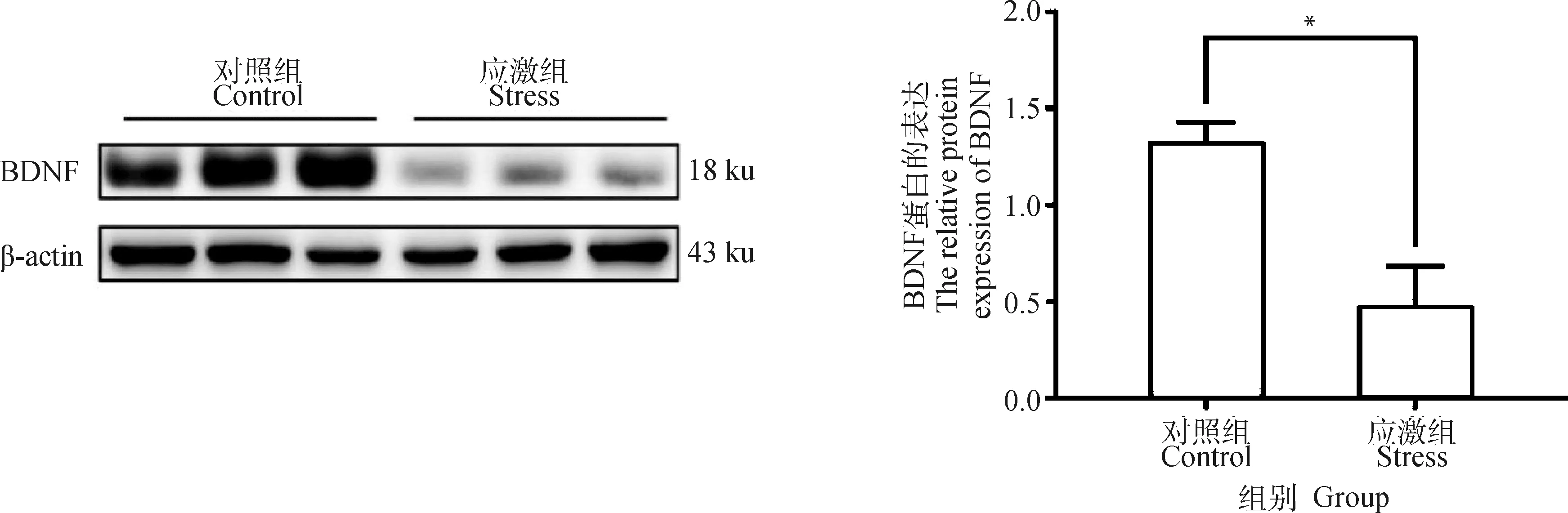
*表示對照組和應激組差異顯著(P≤0.05)* indicates that the difference is significant between the control group and the stress group (P≤0.05)圖6 制動應激對孕豬海馬BDNF蛋白表達水平的影響Fig.6 The effect of restraint stress on BDNF level of hippocampus in pregnant sows
3 討 論
關于慢性應激對中樞神經系統產生的影響已經有了眾多進展,如慢性輕度應激(chronic mildstress,CMS)可降低小鼠海馬內分子層的突觸囊泡密度[26],慢性隔離應激(social instabilitystress)的雄性小鼠海馬中新生神經元中存活率降低,且新生比例降低[27]。考慮到豬和人在應激激素類型和海馬結構上的相似性,因此我們認為用豬作為妊娠期心理應激的模型動物更具有臨床意義。同時,探索限位欄養殖模式對于妊娠母豬生理、心理的影響也是學者和養殖界關心的重要課題。因此,作者通過構建著床期孕豬制動應激模型,探索了著床期慢性心理應激對孕豬海馬神經可塑性的影響。
突觸可塑性(synaptic plasticity)是指突觸的形態會朝著有利于某些神經元通路的方向改變,同時減少與其他神經元通路的聯系,這種可塑性被認為是神經系統應對內、外部刺激時產生的適應性結構重組以改變功能的能力。而神經可塑性不僅反映在海馬神經元構建新的神經通路的能力中,同時也反映在海馬神經元的組織學特性里,包括樹突棘的大小和數量、突觸的數量以及新神經元形成的可塑性。本研究的結果顯示,慢性應激使孕豬海馬的部分細胞形態結構發生了變化,應激組孕豬海馬DG顆粒細胞胞核排列松散,染色不均且整體呈錐形,并發生了神經元丟失的情況;CA3區部分錐體細胞出現空泡狀,整體排列松散,肉眼可見部分神經元位置空缺;而在應激組CA1區的錐體細胞中可見尼氏小體分布減少,染色較淺。CA3區的樹突復雜度也顯著下降,樹突長度則無顯著變化,但本研究鍍銀染色切片的厚度較薄,樹突并不一定能展現其全長并納入統計,這可能會影響樹突長度結果的可信度。樹突棘統計結果顯示,在CA3區,未成熟和成熟樹突棘數量較對照組均有顯著下降,在DG中成熟樹突棘也有顯著下降,這表明制動應激可減少海馬神經元新生樹突棘的形成或影響新生樹突棘的存活率,同時加速成熟樹突棘的消除,共同造成海馬神經元樹突棘密度的降低。細胞形態的變化、樹突復雜度和樹突棘的數量的變化共同表明,慢性心理應激對于海馬的神經元可塑性功能造成了負面影響。
慢性應激通常會引起大腦中BDNF表達水平的降低,降低的程度和應激的性質和程度以及不同腦區都有直接關聯。大量研究表明,糖皮質激素和BDNF表現出對海馬結構和細胞可塑性的相反作用[34],糖皮質激素對海馬神經元可塑性的慢性負面效果可能是通過降低BDNF的表達并下調與其相關的信號通路來達成的[35]。在Radecki等人的研究中,BDNF的緩慢灌注逆轉了慢性應激誘導的海馬長時程增強(LTP)[36-39]和記憶功能的損傷,證明慢性應激造成大腦中神經元可塑性負面變化可能是通過降低神經元營養供給這一方式達成[40]。在本研究中,制動應激后孕豬海馬內的BDNF蛋白表達的顯著下降也可能參與了海馬神經可塑性的負面轉歸。
限位欄養殖對于神經可塑性的影響或許不僅僅與制動應激有關,還可能與運動量的減少有關。有研究顯示[41],運動量對于大腦血管系統的發生也有影響,在有氧運動參數條件下,運動皮層中的血管結構可塑性主要表現為新生血管生成,運動大鼠大腦區域毛細血管直徑增加,內皮細胞核直徑增大。因此,作者推測,限位欄養殖所致的母豬運動量減少,對大腦供血造成影響,可能也是限位欄養殖模式影響神經細胞可塑性的一個因素。
4 結 論
制動應激使應激組孕豬海馬的DG和CA3區出現了神經元丟失現象,CA3區錐形細胞樹突復雜度下降,神經元成熟樹突棘和未成熟樹突棘數量均顯著降低,DG的成熟樹突棘顯著降低。故制動應激減弱了孕豬大腦海馬中CA3區和DG的神經元可塑性。
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
娃娃樂園·綜合智能(2020年8期)2020-08-28 00:32:14
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
作文周刊·小學二年級版(2018年9期)2018-04-18 10:01:40
小學生導刊(2018年1期)2018-03-15 08:02:37
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
科技知識動漫(2016年6期)2016-06-24 21:04:53
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
汽車觀察(2009年1期)2009-02-18 09:11:50
