兩頭烏豬JAZF1基因的克隆及生物信息學分析
2022-01-13 10:44:06徐興莉饒榮海
豬業科學 2021年12期
楊 虎 ,徐興莉 ,饒榮海
(1.宜春學院生命科學與資源環境學院,江西 宜春 336000;2.宜黃縣桃陂鎮獸醫站,江西 撫州 344400)
JAZF1(Ju xtaposed with another zinc finger gene1)基因又名Tip27,ZNF802,最早由全基因組關聯研究發現的與人Ⅱ型糖尿病相關的基因,JAZF1基因對動物糖和脂肪代謝有重要調控作用。盧春敏等將JAZF1基因轉染到小鼠肝細胞,發現JAZF1基因過表達可以使SREBP1基因表達下調,而SREBP1基因表達產物是脂肪代謝調控的重要轉錄因子,這些轉錄因子的調控直接影響脂肪的合成。通過對小鼠研究發現,JAZF1基因過表達能夠顯著抑制肝臟的糖異生作用。這些研究表明JAZF1是體內脂代謝和糖代謝的重要調控因子。JAZF1基因在動物體內對脂肪代謝分子調控機制還不清楚,而脂肪含量特別是肌內脂肪和肌間脂肪含量是評價動物肉質的重要指標。文章以肌內脂肪含量適中的贛西兩頭烏豬為研究對象,克隆JAZF1基因的cDNA全長,并對該序列的組成、編碼蛋白質的特點進行了生物信息學分析,為探索JAZF1基因調控脂肪代謝機制提供基礎。
1 材料與方法
1.1 實驗材料
組織樣品取自贛西兩頭烏豬的背最長肌,且保存于-80 ℃,5'RACE和3'RACE克隆試劑盒分別購自Invitrogen公司和 Clontech 公司。
1.2 實驗方法
1.2.1 總RNA提取及cDNA 合成
用反轉錄酶和特異性引物先合成cDNA第1鏈,然 后 用RNase Mix 對cDNA處理,最后再純化cDNA。
1.2.2 PCR擴增
根據GenBank提供的人JAZF1基因(登錄號:NM_175061)序列設計PCR引物(表1)擴增并克隆測序,即可獲得JAZF1基因部分CDS(coding domain sequence)序列。

表1 引物序列
1.2.3 RACE擴增
1)5'RACE擴增。首先對純化后的cDNA末端加上多聚C,再用試劑盒特異性引物對已經加尾的cDNA進行第一輪擴增;再用特異性引進行第二輪擴增,電泳檢測擴增產物進行并回收,最后將PCR產物與pMD18-T連接,并進行測序,得到長度為684 bp的目的序列。2)3'RACE擴增。 使用試劑盒特異引物及合成的cDNA進PCR擴增,電泳檢測并回收,純化后的PCR產物與載體連接并克隆測序,最終得到420 bp的序列。
1.3 統計分析
1.3.1 JAZF1基因的序列分析
根據克隆測序結果,利用軟件DNAstar7.1 預測JAZF1基因的ORF(Open Reading Frame)并預測氨基酸序列。
1.3.2 豬JAZF1基因的蛋白結構與功能預測
分別預測JAZF1氨基酸的磷酸化位點( http://www.cbs.dtu.dk/services/NetPhos/)、潛在的糖基化位點(http://www.cbs.dtu.dk/services/NetOGlyc/)及JAZF1蛋白的親水性(http://web.expasy.org/protscale/),利用Smart(http://smart.embl-heidelberg.de/)分析氨基酸序列的結構域。
2 結果與分析
本實驗成功克隆了兩頭烏豬JAZF1基因部分CDS序列,利用RACE技術成功獲得了該基因的5'末端序列和3'末端序列。電泳檢測5'RACE和3'RACE的兩個產物片段,結果如圖1所示,兩頭烏豬部分CDS序列產物長為 620 bp(圖1A),5'RACE 產物長 684 bp(圖1B),3'RACE 產物長420 bp(圖1C), 回收條帶并克隆測序。 最后得到了JAZF1基因的cDNA全長序列,共1 098 bp,包括ORF 732 bp,編碼243個氨基酸,氨基酸序列見圖2(DNASTAR)。

圖1 豬JAZF1基因 1%瓊脂糖凝膠電泳圖

圖2 豬JAZF1核苷酸序列及其推導的氨基酸序列圖
2.1 豬JAZF1蛋白結構分析
2.1.1 蛋白理化性質分析
豬JAZF1蛋白分子量為27 079.4 Da,等電點為8.63;JAZF1蛋白的氨基酸中,絲氨酸(Ser)含量為11.9%,所占比例最高,含量最低為蛋氨酸(Met),比例為2.1%。
2.1.2 JAZF1蛋白跨膜螺旋結構
TMPRED分析蛋白跨膜結構,顯示豬JAZF1蛋白有1個跨膜結構,且由內向外,跨膜區的氨基酸殘基位置在8-25處,見圖3。

圖3 TMPRED軟件預測JAZF1蛋白跨膜結構
2.1.3 JAZF1潛在磷酸化位點和糖基化位點分析
在線分析JAZF1蛋白的磷酸化位點,發現2個蘇氨酸(Thr)、3個酪氨酸(Tyr)及18個絲氨酸(Ser)屬于蛋白激酶磷酸化位點(圖4); 糖基化位點有19個(圖5)。
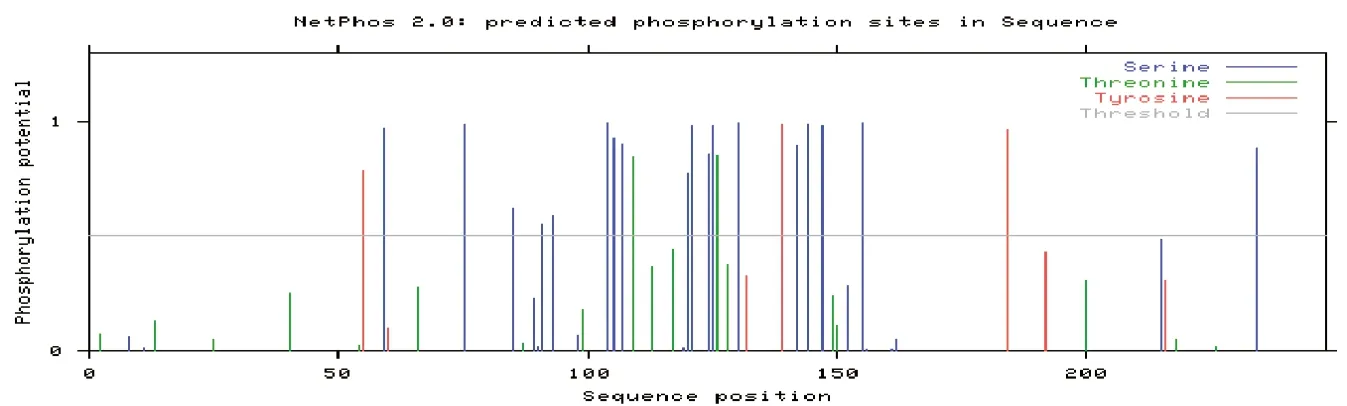
圖4 JAZF1蛋白磷酸化位點
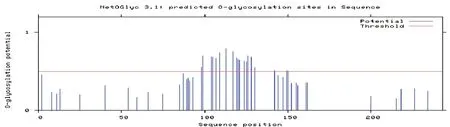
圖5 JAZF1糖基化位點
2.1.4 JAZF1蛋白親疏水性分析
蛋白疏水性分析顯示,豬JAZF1基因編碼蛋白為水溶性蛋白,如圖6所示。JAZF1蛋白疏水性最小值為-2.700,,最大值為1.511,氨基酸序列內大部分屬于親水性氨基酸。
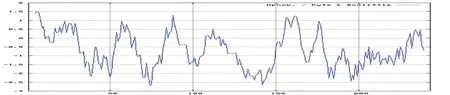
圖6 JAZF1氨基酸親水性分析
2.1.5 蛋白結構域分析及功能預測
結構域顯示,豬JAZF1氨基酸殘基分別有3個 ZnF-C2H2 的結構域;JAZF1蛋白功能分析結果如表2所示,發現該蛋白在能量代謝、脂肪酸代謝的作用概率明顯高于其他功能,蛋白運載結合概率最高為2.030,說明JAZF1基因在脂肪酸代謝和轉運中發揮重要作用,可能對調節脂肪的沉積有重要影響。
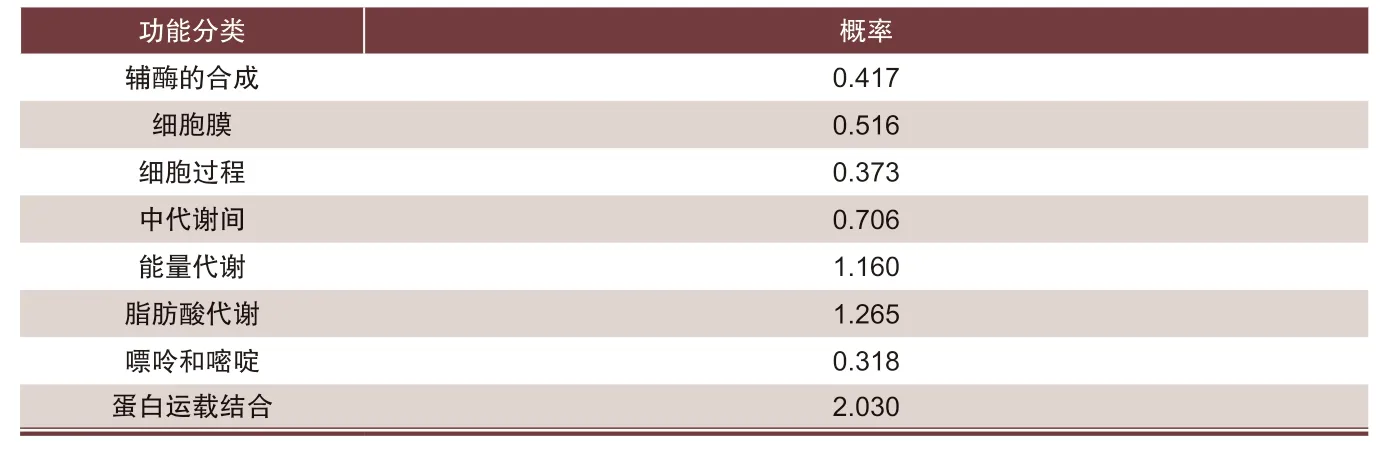
表2 JZAF1蛋白功能預測結果
3 討論與結論
JAZF1作為一個新發現的基因,其對糖代謝和脂肪代謝的重要調控作用已經得到證實。糖代謝方面,JAZF1的表達與人的二型糖尿病密切相關,其詳細調控機制目前尚不明晰。脂肪代謝方面主要是通過抑制脂肪形成和促進其分解來調控,有研究證實,JAZF1主要是通過抑制細胞內的脂肪酸和甘油三酯的沉積,來提高肝臟脂肪酸的β氧化作用,進而促進脂肪分解。梁艷等研究發現,隨著日齡的增加,JAZF1基因在豬肌肉中的表達呈現下降趨勢,研究結果顯示JAZF1基因在肌肉發育和肌內脂肪沉積過程中發揮著重要調控作用;朱才業對不同品種羊全基因組關聯分析,證實JAZF1基因與脂肪形成密切相關。JAZF1的結構域顯示該蛋白含有鋅指家族中最為普遍的C2H2結構域類型,C2H2結構域根據含鋅指結構數量的多少可分為三類鋅指蛋白,含3個鋅指結構的C2H2型鋅指蛋白(tC2H2),含多個C2H2結構的鋅指蛋白(maC2H2);鋅指成對間隔排列的 spC2H2 型。豬的JAZF1蛋白結構含有3個鋅指結構,屬于tC2H2型鋅指蛋白,每個鋅指由一個螺旋和2個折疊組成,鋅指間由接頭連接,接頭內氨基酸具有保守性,3個鋅指在蛋白內串聯排列均結合到DNA大溝內。此類結構域通常作為重要的轉錄調控因子參與表達調控,所以JAZF1蛋白極有可能是參與肌內脂肪沉積的重要調控因子。
蛋白質磷酸化糖基化是最常見、最重要的蛋白質翻譯后修飾方式之一,也是一種普遍的生命活動調節方式,在信號傳遞等過程中發揮重要作用。利用生物信息學軟件分析,發現JAZF1蛋白存在18個絲氨酸磷酸化位點,3個酪氨酸磷酸化位點,2個蘇氨酸磷酸化位點及19個糖基化位點, 這些位點可能在細胞的信號轉導、分化及細胞生長等多個生理過程中發揮著重要作用。JAZF1蛋白功能的預測表明該蛋白對脂肪酸代謝及能量代謝有重要的調節作用,JAZF1基因對脂肪的合成可能有重要影響。
