草魚褪黑激素受體Mtnr1基因克隆及組織表達分析*
2022-01-14 07:48:44袁漢文郜衛華陳敦學郭利偉許巧情
漁業科學進展 2022年1期
胡 偉 吳 慧 袁漢文 郜衛華 陳敦學 郭利偉 許巧情
(1. 長江大學 濕地生態與農業利用教育部工程研究中心 湖北 荊州 434023;2. 湖南師范大學 省部共建淡水魚類發育生物國家重點實驗室 湖南 長沙 410081;3. 貴州大學動物科學學院 貴州 貴陽 550025)
褪黑激素(melatonin)是一種廣泛分布于動物、植物、細菌和真菌中的天然小分子物質(Xu et al, 2017)。在哺乳動物中,褪黑激素主要由下丘腦中的松果體在夜間分泌,在性腺、骨髓、視網膜、晶狀體、腎臟和肝 臟 等 組 織 中 也 有 發 現 (Carpentieri et al, 2012;Acu?a-Castroviejo et al, 2014)。褪黑激素不僅具有調節晝夜節律、清除自由基的作用(Han et al, 2017),還能通過內分泌、旁分泌和自分泌方式保護細胞抗氧化和炎癥損傷、改善胰島素抵抗(Heo et al, 2018)、調控生殖發育(Maitra et al, 2016)、調節脂質代謝(Yang et al, 2017)和控制血壓(de Berardis et al, 2013)。褪黑激素還能參與維持神經元的內穩態(Hermoso et al,2016)和保護機體免疫系統的功能 (艾克拜爾·熱合曼等, 2018; Zhang et al, 2019)。
褪黑激素的功能主要通過與其高親和力的特異性褪黑激素受體(melatonin receptor, Mtnr1)結合,再進一步啟動信號級聯反應,參與細胞內各種生理過程。Mtnr1屬于G蛋白偶聯受體超級家族(Brydon et al,1999),與其他G蛋白偶聯受體結構相似,包括7個跨膜域、1個G蛋白結合位點、多個磷酸化以及糖基化位點(Klosen et al, 2019)。另外,第 3跨膜域的下游含有NRY結構域和C(C/Y) ICHS結構域,第7跨膜域中存在 NPXXY 結構域(Reppert et al, 1995b; Klosen et al, 2019)。Mtnr1在脊椎動物中存在3種不同亞型,分別為 Mtnr1A (Mel 1a或 MT1)、Mtnr1B (Mel 1b或MT2)和 Mtnr1C (Mel 1c或GPR50) (Reppert et al,1995a),而哺乳動物中僅有前 2種亞型:Mtnr1A和Mtnr1B (Gaildrat et al, 2000),僅在魚類等低等脊椎動物中發現第 3種亞型 Mtnr1C (Dufourny et al, 2008)。此外,魚類特有基因組加倍事件導致Mtnr1A和Mtnr1B存在多個不同亞型,如斑馬魚(Daniorerio)Mtnr1Aa和Mtnr1Ab(Reppert et al, 1995b);虹鱒(Oncorhynchus mykiss) Mtnr1Aa、Mtnr1Ab和 Mtnr1B(Mazurais et al, 1999);星點東方鲀(Takifugu niphobles)Mel1a1.4、Mel1a1.7 和 Mel1b (Ikegami et al, 2009b);大彈涂魚(Boleophthalmus pectinirostris) Mel1a1.4、Mel1a1.7 和Mel1b (朱文博等, 2012; Hong et al,2014);大西洋鮭(Salmo salar) Mtnr1Aaα、Mtnr1Aaβ、Mtnr1Ab、Mtnr1al和 Mtnr1B (Ciani et al, 2019);金魚(Carassiusauratus)Mel1a1.4、Mel1a1.7 和 Mel1b(Ikegami et al, 2009a)。值得注意的是,Mtnr1廣泛參與機體神經調節(Klosenet al, 2019)、節律同步(Ikegami et al, 2009b)、內分泌激素合成(Ciani et al,2019)和生長繁殖(Ikegami et al, 2009b; Hong et al,2014)等生理活動。例如,敲除Mtnr1A基因后的小鼠表現出焦慮、類似抑郁癥,增加了獎勵和上癮傾向,降低了快速眼動睡眠(Comaiet al, 2019),生理節律(包括年生殖節律)也發生改變(Ng et al, 2017)。另外,Mtnr1在機體的細胞和體液免疫反應過程發揮積極作用,如在人(Homosapiens)淋巴細胞(Jurkat)和單核細胞(U937)研究中發現,褪黑激素能通過Mtnr1增強其白細胞介素 IL-2和 IL-6分泌(García-Mauri?oet al,2000),添加MtnrB抑制劑Luzindole后,不僅降低了人淋巴細胞中IL-2和 IL-2受體的分泌(Carrillo-Vico et al, 2004),還增強了前列腺素E2對IL-2的抑制效果(Carrillo-Vico et al, 2005)。此外,褪黑激素能促進野生型和MtnrA敲除型小鼠脾淋巴細胞增殖,而抑制MtnrA 蛋白后增殖現象消失(Drazen et al, 2001)。這些研究證實了Mtnr1參與哺乳動物神經活動和免疫過程,但有關硬骨魚類Mtnr1的研究仍處于起步階段。
草魚(Ctenopharyngodonidella)是我國重要的淡水經濟魚類,其2018年產量達到5.5萬t (農業農村部漁業漁政管理局等, 2019)。草魚疾病的暴發與其免疫調節能力密切相關,褪黑激素在機體免疫過程中具有重要作用,Mtnr1是褪黑激素發揮生理功能的關鍵受體分子,因此,解析Mtnr1基因在草魚免疫調節中的功能對草魚疾病的免疫防治具有重要意義。本研究以草魚為研究對象,克隆得到Mtnr1基因6種異構性的全長cDNA序列,通過生物信息學技術,研究不同亞型的序列和結構特征,同時,采用實時熒光定量PCR (RT-qPCR)檢測6種亞型在不同組織/器官中的分布和表達模式,研究結果可為Mtnr1在脊椎動物中的進化補充數據,同時,可為后續深入研究其基因功能以及揭示Mtnr1在草魚免疫過程中的作用奠定分子基礎。
1 材料與方法
1.1 實驗材料
實驗草魚購自湖北荊州水產市場,大小規格為(1.00±0.25) kg,暫養在實驗室循環養殖系統2周。水溫為(24±2)℃。飽食投喂 2次/d (09:00和 17:00)。暫養結束后,饑餓 24 h 取樣,用 MS-222 (10 mg/L)麻醉魚體,在冰上迅速取出其肝臟、心臟、鰓、腦、肌肉、前腸、中腸、后腸和腎臟9個組織,液氮速凍后,存于-80℃冰箱備用。
1.2 實驗方法
1.2.1 總RNA的提取和cDNA的合成 采用Total RNA kit I提取試劑盒(Omega, 美國)提取總 RNA,使用1%的瓊脂糖凝膠電泳和超微量核酸蛋白測定儀(Thermo Scientific, 美國)檢測 RNA 完整性和濃度。利用MonScript? RTⅢ Super Mix with dsDNase(two-step) (莫納, 蘇州)合成 cDNA,嚴格按照試劑盒說明書操作,PCR產物保存于-20℃。
1.2.2 草魚 Mtnr1基因全長 cDNA 克隆 利用斑馬魚、鯉魚(Cyprinus carpio)、鯽魚(Carassius auratus)以及哺乳動物人類、褐家鼠(Rattus norvegicus)等已公布Mtnr1A、Mtnr1B和Mtnr1C基因序列,比對保守區域后設計引物(表 1),進行擴增實驗。再基于獲得的部分cDNA序列,分別利用RACE技術,按照RACE試劑盒(TaKaRa)對 Mtnr1的5′和3′端進行擴增。擴增產物測序正確后,通過DNAStar軟件拼接獲得完整的cDNA序列。
1.2.3 Mtnr1基因序列分析 利用 NCBI中 Blast (http://blast.ncbi.nlm.nih.gov/)分析獲得的 Mtnr1不同亞型 cDNA序列的正確性。通過 NCBI網站的 ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/)查 找Mtnr1不同亞型的ORF框。使用ExPASy (http://web.expasy.org/translate/)翻譯Mtnr1不同亞型cDNA編碼蛋白序列。利用ProtParam在線軟件預測Mtnr1不同亞型的蛋白基本理化特性(https://web.expasy.org/ protparam/)。通過Tmpred在線軟件對Mtnr1不同亞型的跨膜域進行預測(http://www.cbs.dtu.dk/services/ TMHMM/#opennewwindow)。并采用Singal4.1軟件預測其信號肽。采用Phyre2軟件對 Mtnr1不同亞型蛋白質的三級結構進行預測(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)。利用Cell-PLoc2.0在線軟件(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)預測Mtnr1的亞細胞定位。通過NetPhos和NetNGlyc軟件分別對蛋白磷酸化位點(http://www.cbs.dtu.dk/ services/NetPhos/)和糖基化位點(http://www.cbs.dtu.dk/services/NetNGlyc/)進行分析。采用ClustalW軟件對 Mtnr1不同亞型不同物種間進行序列比對和一致性分析。最后利用MEGA 5.2軟件,以鄰接法(NJ)對Mtnr1構建系統進化樹,設定bootstrap值為1000。
1.2.4 草魚Mtnr1基因的表達模式 隨機選取4條暫養后的草魚,分別提取9個不同組織(肝臟、心臟、鰓、腦、肌肉、前腸、中腸、后腸和腎臟)的總RNA,設置4個生物學重復。以總RNA為模板,按照MonScript? RTⅢ Super Mix with dsDNase (莫納,蘇州)說明書合成cDNA,并將cDNA稀釋 10倍作為RT-qPCR模板,保存于-20℃備用。Mtnr1基因的組織表達模式測定方法參考侯吉倫等(2019):依據克隆獲得的草魚Mtnr1基因序列,利用Primer 5.0軟件,設計Mtnr1基因RT-qPCR定量引物RT-F和RT-R,并以β-actin為內參基因(表1)。根據說明書設置定量反應體系:SYBR?fast qPCR master mixⅡ20 μL,cDNA模板 4 μL,引物各 0.5 μL。RT-qPCR 反應程序:95℃ 5 min;95℃ 15 s,60℃ 30 s,72℃ 20 s,共 40個循環。利用 2-ΔΔCt方法(Livak et al, 2002)檢測 Mtnr1不同亞型在不同組織中的表達模式。
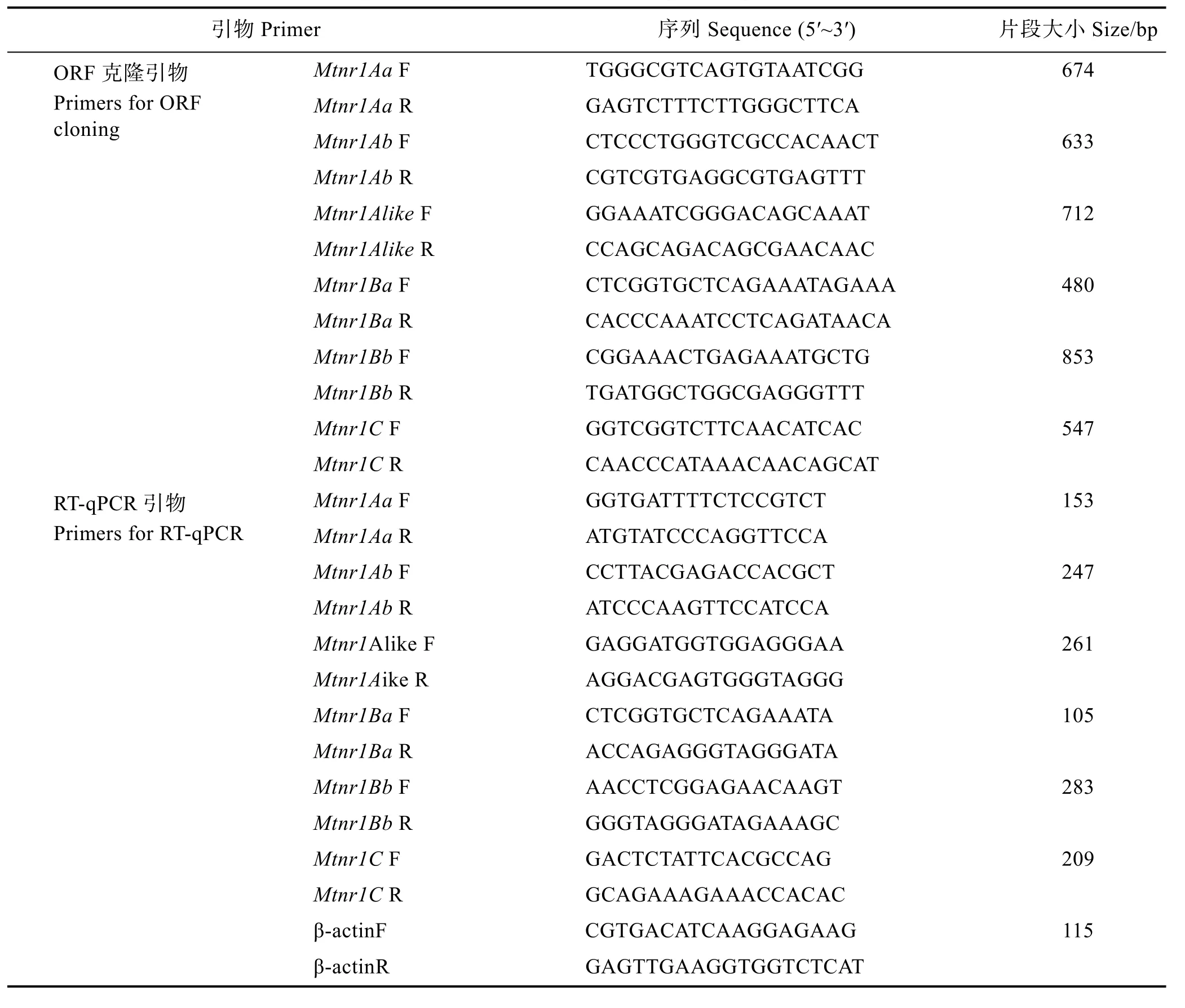
表1 草魚褪黑激素受體Mtnr1基因克隆擴增引物Tab.1 Amplification primers used for cloning Mtnr1 genes from C. idella
1.3 統計分析
實驗結果均采用平均值±標準誤(Mean±SE)表示,為確保實驗過程中數據的準確性,設置4個生物學重復,每個生物學重復設置3個技術重復。采用SPSS 18.0進行統計分析,采用單因素方差分析(one-way ANOVA)的Tukey法對Mtnr1不同亞型的相對表達量進行兩兩比較,顯著性水平設置為0.05。
2 結果與分析
2.1 Mtnr1基因亞型cDNA序列分析
利用DNAStar將克隆獲得的cDNA進行拼接,并通過NCBI中的Blast對獲取的序列進行正確性驗證,本實驗克隆得到草魚6個Mtnr1 cDNA序列,分別為Mtnr1Aa、Mtnr1Ab、Mtnr1Alike、Mtnr1Ba、Mtnr1Bb和Mtnr1C,并將其提交至NCBI的GenBank,序列號分別為MT883605、MT883607、MT883608、MT883606、MT883609和MT883610,其全長分別為2045、2036、2031、2799、2535和 2477 bp (表 2)。ProtParam軟件預測發現,草魚Mtnr1基因編碼蛋白的氨基酸數、分子量和等電點分別見表2。Mtnr1Aa、Mtnr1Ab、Mtnr1Alike、Mtnr1Ba、Mtnr1Bb和 Mtnr1C基因編碼蛋白均由20種氨基酸組成,含量最高的氨基酸分別為纈氨酸(12.6%)、纈氨酸(11.3%)、亮氨酸(11.6%)、亮氨酸(13.0%)、亮氨酸(11.6%)和亮氨酸(12.5%),含量最低的氨基酸分別為色氨酸(1.7%)、組氨酸(1.4%)、谷氨酰胺(1.5%)、組氨酸(1.1%)、組氨酸(1.2%)和組氨酸(1.4%)。帶正(負)電氨基酸殘基數分別為 28(17)、18(32)、19(33)、17(35)、16(30)和 11(31)。原子組成分別為C1853H2853N461O475S18、C1877H2905N463O479S17、C1836H2845N463O458S15、C1820H2883N489O470S18、C1819H2825N463O463S13和C1839H2885N481O484S20。蛋白質不穩定系數分別為35.19、35.32、30.93、38.67、35.74和 34.75,均屬于穩定蛋白。總平均親水性分別為0.560、0.560、0.556、0.490、0.675和0.502,均為親水性蛋白。脂肪酸指數分別為113.77、116.36、120.90、115.24、119.28和109.58。
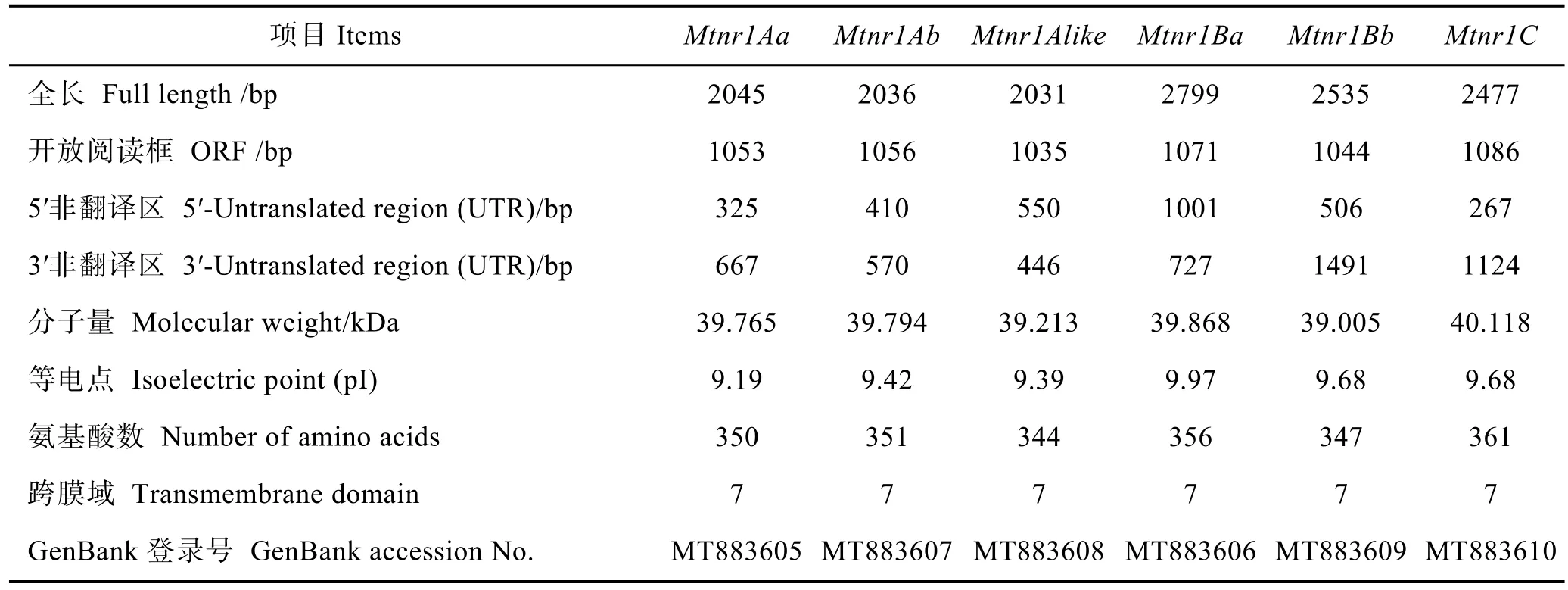
表2 草魚褪黑激素受體Mtnr1 cDNA基本信息Tab.2 The information of Mtnr1 cDNA of C. idella
2.2 Mtnr1亞型蛋白結構分析
利用TMpred在線軟件對草魚Mtnr1的6種亞型蛋白跨膜域進行分析。結果顯示,草魚Mtnr1的6種亞型均存在7個跨膜結構,包括N端、跨膜區域、細胞膜內和膜外區域以及C端,表明其為非胞內蛋白。通過Cell-PLoc 2.0在線軟件預測Mtnr1的6種亞型編碼蛋白亞細胞定位,顯示均定位于細胞膜。通過SignalP4.1預測草魚Mtnr1的6種基因編碼蛋白質的氨基酸序列,均未發現信號肽序列,說明6種亞型編碼蛋白均為非分泌型蛋白。利用 Phyre2在線軟件構造草魚Mtnr1的6種模擬蛋白3D亞型與人Mtnr1A和Mtnr1B蛋白以及斑馬魚的6種模擬結構對比,結果顯示,蛋白結構較為相似(圖1)。此外,通過多重氨基酸序列比較可知,Mtnr1的6種亞型具有 NRY結構域、CYICHS結構域、NAXXY結構域和G蛋白偶聯受體結合位點(圖2)。利用NetNGlyc 1.0和NetPhos 3.1預測功能位點,結果發現,6種亞型受體的糖基化位點和磷酸化位點數分別為Mtnr1Aa(Asn4、Asn9和 Asn117/22、Mtnr1Ab (Asn7、Asn13和Asn121)/28、Mtnr1Alike (Asn37和 Asn144)/17、Mtnr1Ba(Asn4、Asn10和Asn122)/24、Mtnr1Bb(Asn10和Asn121)/25和 Mtnr1C (Asn7、Asn25和 Asn121)/30。
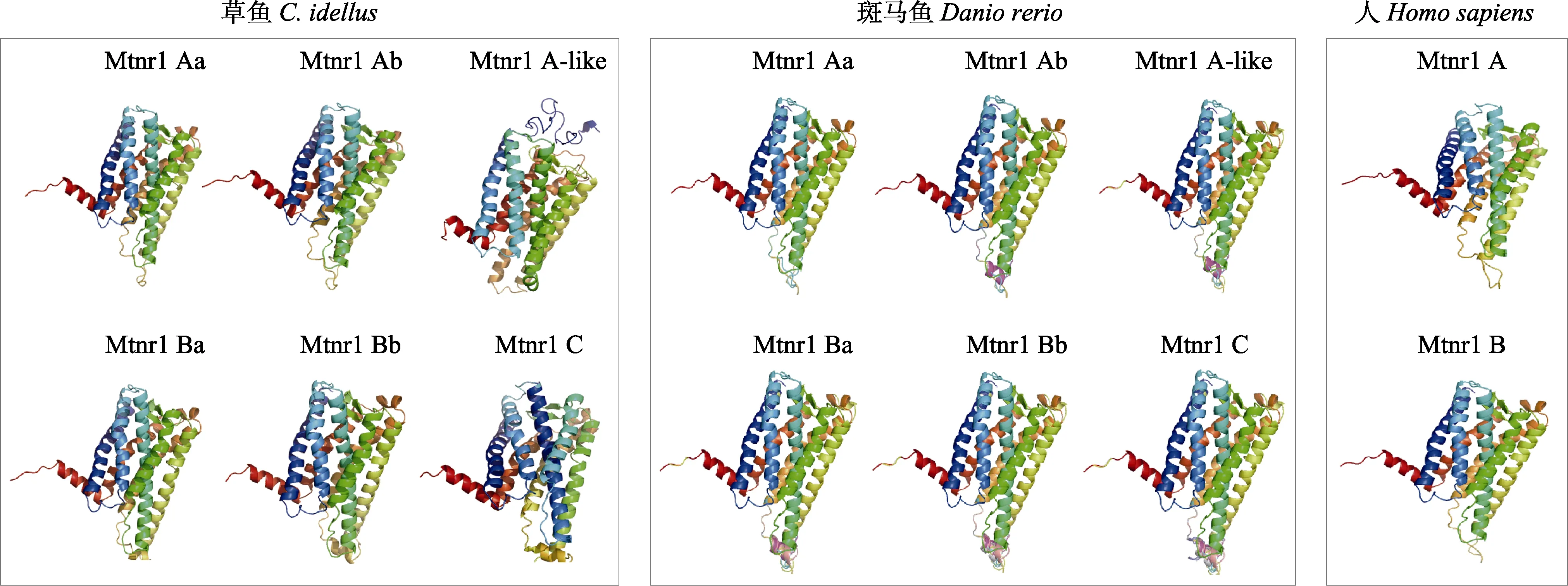
圖1 草魚、斑馬魚和人的基于Mtnr1亞型氨基酸序列預測的蛋白3D結構Fig.1 Protein structure prediction of C. idellus, Danio rerio and Homo sapiens based on sequences of Mtnr1

圖2 草魚不同Mtnr1亞型氨基酸序列多重比對分析結果Fig.2 Multiple alignment of the deduced amino acid sequence of grass carp Mtnr1 subtypes
2.3 Mtnr1同源性及系統進化分析
基于ClustalW,對草魚Mtnr1的6種不同亞型編碼的氨基酸序列同已有物種的Mtnr1序列同源性進行比較分析,結果見表3。6種亞型與其他魚對應的亞型同源性很高,分別為93.1%~99.4%、84.8%~95.2%、82.8%~96.8%、90.1%~97.5%、79.7%~98.3%和90.6%~95.6%。另外,運用MEGA 5.2軟件構建草魚Mtnr1的6種亞型氨基酸序列的系統進化樹(圖3),Bootstrap重復次數設為1000次迭代。結果表明,構建的進化樹與氨基酸的多重比較結果較一致。草魚Mtnr1的6種亞型均與其他魚類的對應亞型聚為一支,且與鯉魚、鯽魚和斑馬魚親緣關系最近。
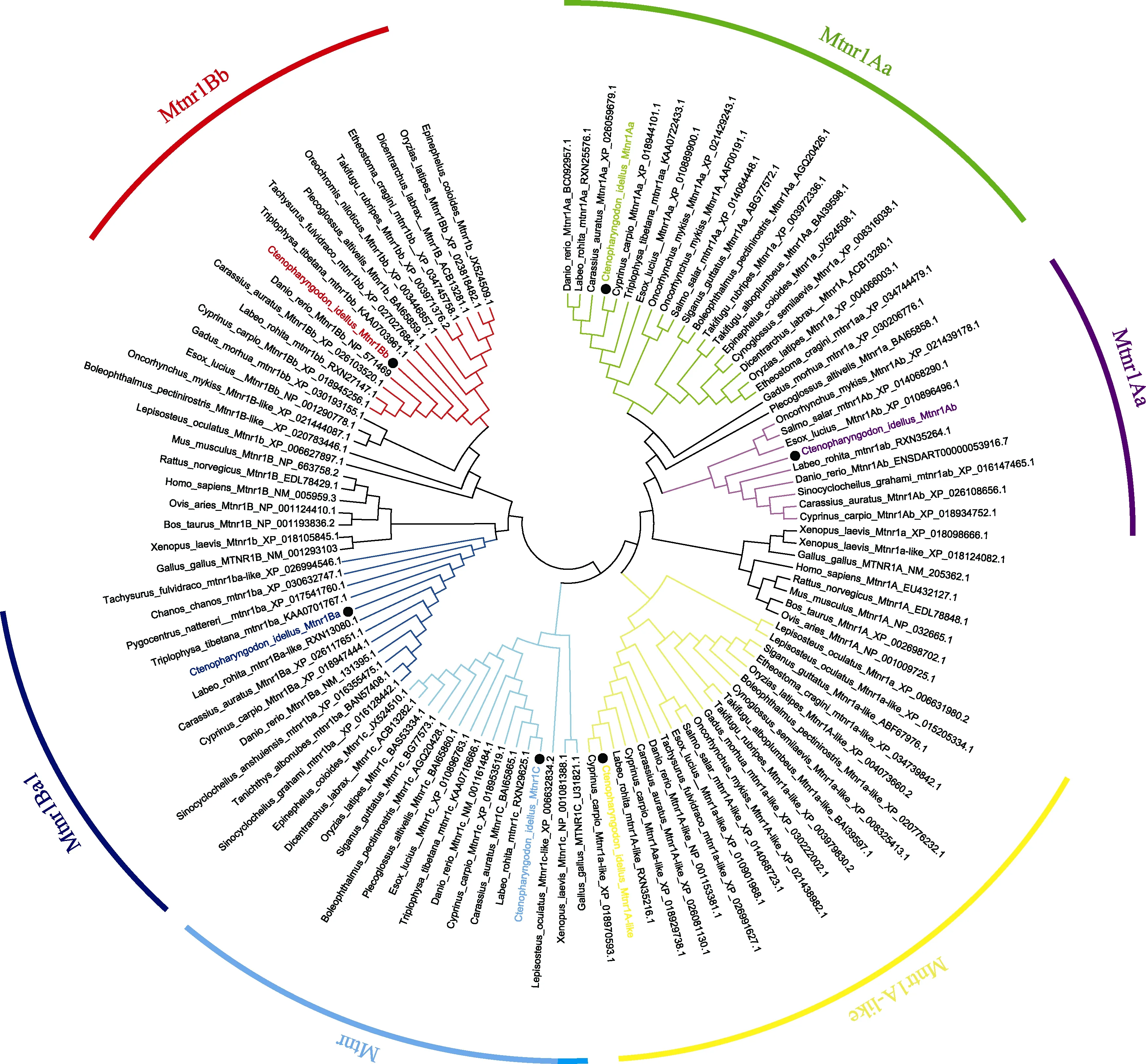
圖3 采用MEGA 5.2軟件構建的基于草魚Mtnr1氨基酸序列的NJ系統進化樹Fig.3 Phylogenetic tree based on the protein sequences of Mtnr1 from C. idellus and other vertebrate species using the neighbor-joining (NJ) method in MGEA 5.2
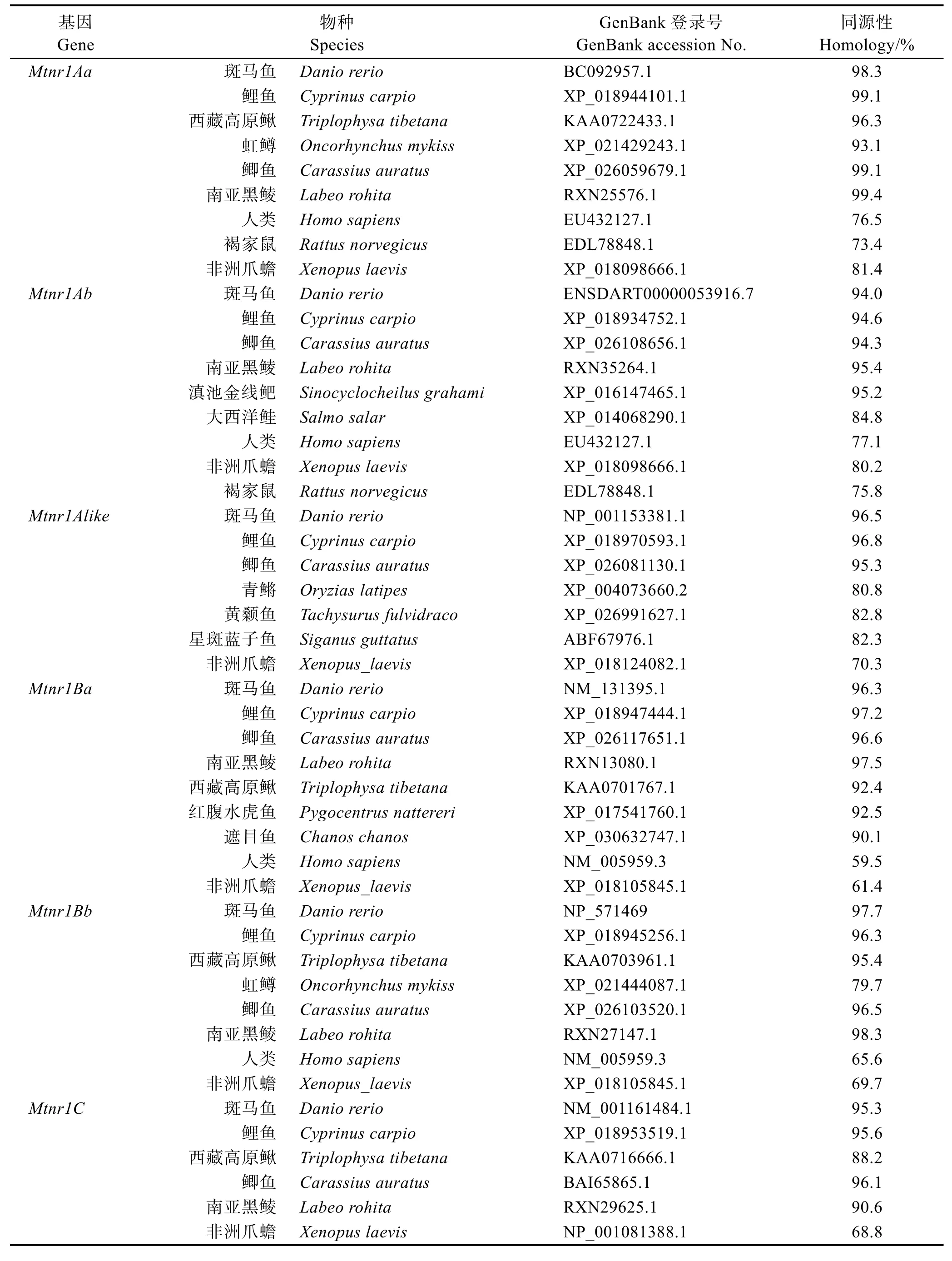
表3 草魚Mtnr1氨基酸序列與其他物種對應氨基酸序列的同源性Tab.3 Homologies of deduced amino acid sequences of six Mtnr1 isoforms gene of C. idellus and other species
2.4 草魚Mtnr1組織表達模式
采用RT-qPCR技術檢測了Mtnr1基因在草魚肝臟、心臟、鰓、腦、肌肉、前腸、中腸、后腸和腎臟9個組織的表達情況。結果顯示,Mtnr1的6種亞型在所檢測的組織中均有表達(圖4)。其中,Mtnr1Aa基因在腦組織中表達量最高(P<0.05),腎臟、鰓、中腸、心臟次之,而在肝臟組織中表達量最低(P<0.05)。Mtnr1Ab基因在腦組織中表達量最高(P<0.05),然后依次為腎臟、肌肉、后腸、肝臟和心臟,在鰓、前腸及中腸中表達量最低(P<0.05),而鰓、前腸以及中腸之間表達量無顯著差異(P>0.05)。Mtnr1Alike基因在腎臟中表達量最高(P<0.05),然后依次為腦、前腸、后腸和中腸,而在肝臟中表達量最低(P<0.05)。Mtnr1Ba基因 mRNA水平在腦中表達量最高,其次在腎臟、前腸和中腸表達量居中,隨后心臟、鰓、肌肉和后腸中表達量次之,且它們之間無顯著性差異(P>0.05),在肝臟中表達量最低(P<0.05)。Mtnr1Bb在腦中含量最高,其次為心臟,在其余組織中含量最低且它們之間無顯著性差異(P>0.05)。Mtnr1C基因表達量在腎臟組織中最高(P<0.05),其次為腦、中腸和心臟,在肝臟組織中含量為最低(P<0.05)。此外,從對亞型表達熱圖可知(圖5),6種亞型均在腦和腎臟組織中大量表達,其次為腸道組織,進一步發現,在腦組織中Mtnr1Aa基因表達量最高(P<0.05),其次為Mtnr1Ba、Mtnr1C、Mtnr1Alike和 Mtnr1Bb,而 Mtnr1Ab基因表達量最低(P<0.05) (圖5)。在腎臟組織中發現Mtnr1Ba表達量最高,其次為Mtnr1C、Mtnr1Alike、Mtnr1Aa和 Mtnr1Ab,而 Mtnr1Bb基因表達量最低(P<0.05) (圖5)。另外,在前腸中發現Mtnr1Ba基因表達量最高(P<0.05),其次為 Mtnr1Alike、Mtnr1C、Mtnr1Aa和Mtnr1Bb,而Mtnr1Ab表達量最低(P<0.05)。中腸中 Mtnr1Ba 基因表達量最高(P<0.05),其次為Mtnr1C、Mtnr1Alike、Mtnr1Aa和Mtnr1Bb,而Mtnr1Ab表達量最低(P<0.05)。后腸中Mtnr1Ba和Mtnr1Alike基因表達量最高(P<0.05),且二者之間無顯著差異(P>0.05),其次為Mtnr1C、Mtnr1Aa和Mtnr1Ab,而 Mtnr1Bb表達量最低(P<0.05) (圖 5)。
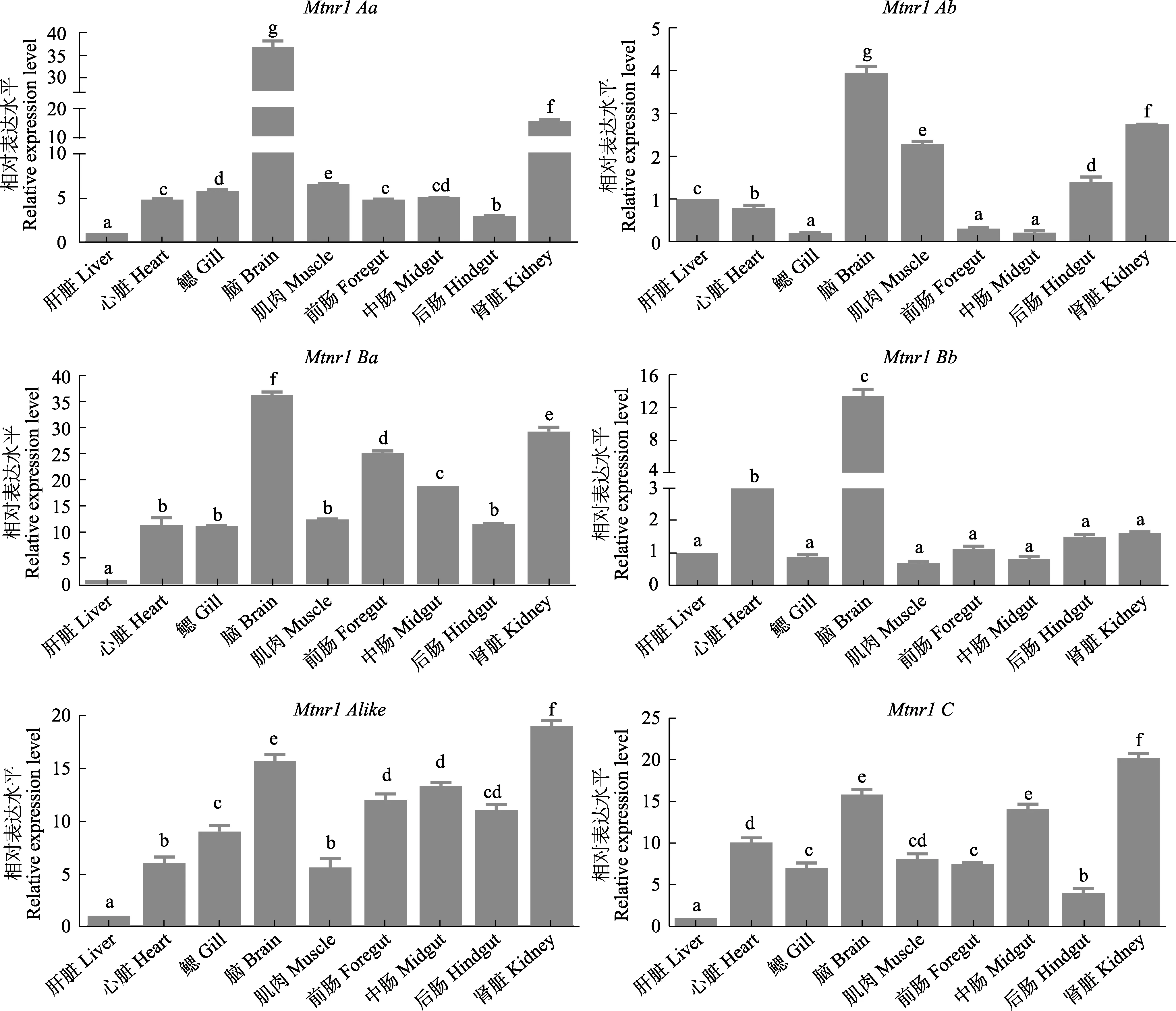
圖4 草魚Mtnr1基因6種亞型在不同組織中的表達模式Fig.4 Expression of 6 isoforms of Mtnr1 mRNA in different tissues of C. idellus
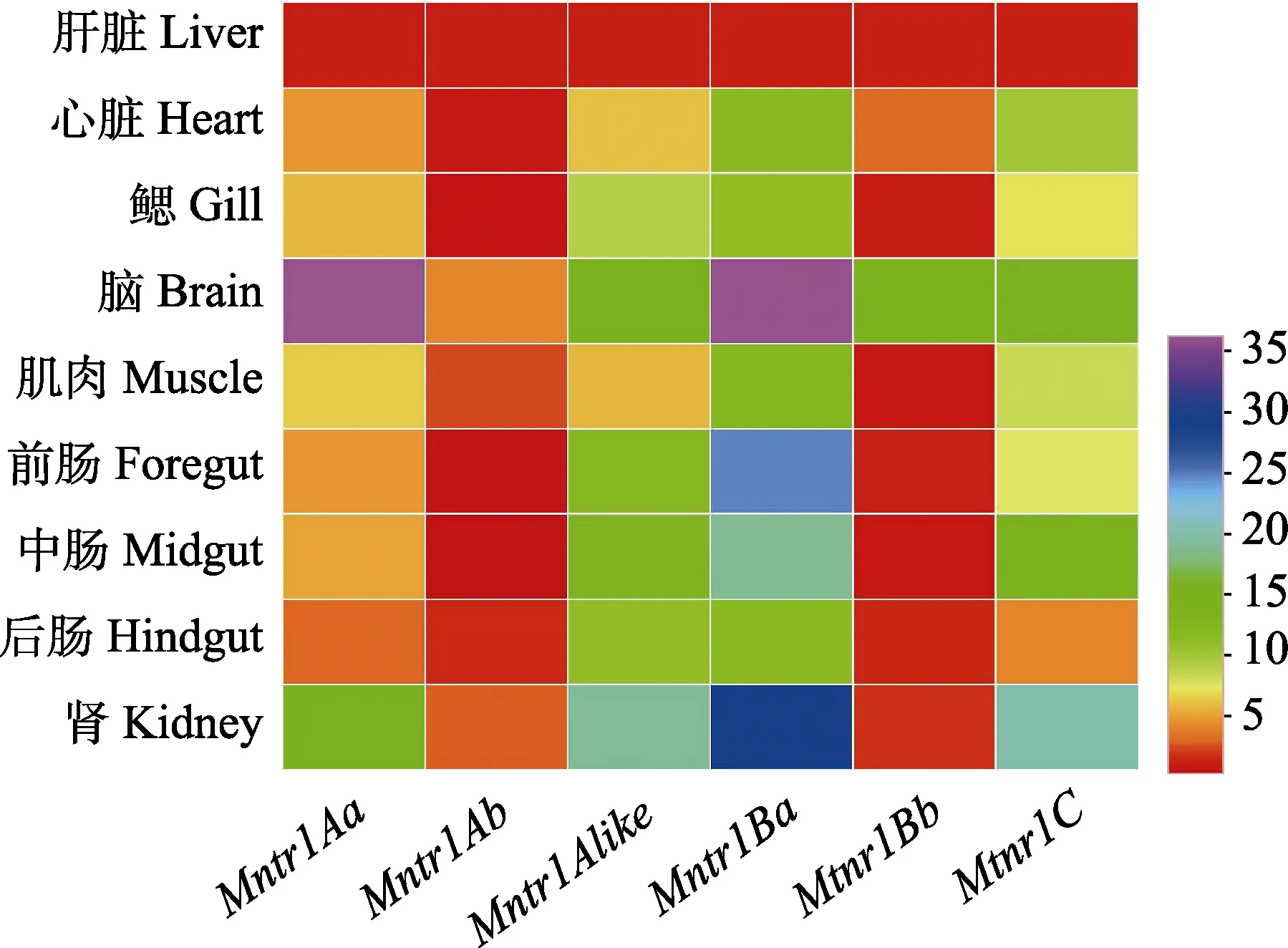
圖5 Mtnr1亞型的組織表達熱圖Fig.5 Heatmap of expression pattern of the Mtnr1 family mRNA in different tissues of C. idellus
3 討論
本研究成功克隆得到草魚Mntr1基因6個亞型的cDNA全長序列,全長序列長度為2031~2799 bp,編碼的氨基酸個數為344~361,無信號肽序列,均屬于非分泌型蛋白。Mtnr1的6種亞型由7個跨膜結構(Ⅰ~Ⅶ)構成,屬于典型的G 蛋白偶聯受體(Shiu et al,1998)。多序列比對結果顯示,草魚Mtnr1存在NRY結構域、CYICHS結構域、NAXXY結構域和 G蛋白偶聯受體結合位點以及多個糖基化和磷酸化位點,這與大西洋鮭和大彈涂魚的Mtnr1基因研究結果一致(Hong et al, 2014; Ciani et al, 2019; Klosen et al,2019),暗示草魚Mtnr1在進化中具有較高的保守性。另外,6種亞型的3D結構與斑馬魚和人的Mntr1空間結構相似,進一步證實其進化保守性。同時,同源性比較發現,草魚 Mtnr1基因6種亞型(Mtnr1Aa、Mtnr1Ab、Mtnr1Alike、Mtnr1Ba、Mtnr1Bb和 Mtnr1C)與其他魚對應的Mntr1亞型同源性很高,分別為93.1%~99.4%、84.8%~95.2%、82.8%~96.8%、90.1%~97.5%、79.7%~98.3%、和90.6%~95.6%。系統進化樹結果顯示,草魚Mtnr1基因6種亞型與鯉魚、鯽魚和斑馬魚等鯉科(Cyprinidae)魚類的各亞型聚為一支,表明草魚Mtnr1與鯉科魚類親緣關系較近,這與它們的進化地位一致。
Mtnr1的6種亞型基因在草魚腦、肝臟、心臟、鰓、肌肉、前腸、中腸、后腸和腎臟9個組織中均有表達,表明Mntr1可能廣泛參與機體各種生理活動(Hong et al, 2014)。本研究發現,Mtnr1Aa、Mtnr1Ab、Mtnr1Ba和Mtnr1Bb 4種亞型集中在腦組織中表達,暗示其主要參與神經調節相關的活動(Klosen et al,2019)。這與大西洋鮭(Ciani et al, 2019)、星點東方鲀(Ikegami et al, 2009b)和金魚(Ikegami et al, 2009a)中的研究相似。如 Ikegami等(2009b)研究發現,Mel1a 1.4、Mel1a 1.7和Mel1b均在星點東方鲀腦組織、視網膜、腦垂體和外圍組織中表達,且在腦組織中表達量最高。另外,褪黑激素在生物鐘網絡中既是視交叉上核(suprachiasmatic nuclei, SCN)的時鐘輸出,又是內部同步器,表明它的主要功能是充當時鐘輸如“time giver” (Pevet et al, 2011)。而褪黑激素功能發揮依賴于Mtnr1的表達,Mtnr1在腦組織中的高表達,間接證實褪黑激素在草魚機體神經調節過程中具有重要作用。
此外,本研究顯示,Mtnr1Ba基因在草魚前腸、中腸和后腸中均高表達,Mtnr1Alike在后腸中高表達,Mtnr1的表達呈現組織差異性。這與哺乳動物和水禽類中的研究類似,如大鼠MT2在其結腸中mRNA水平最高,而 MT1基因在十二指腸內表達量最高,在回腸和空腸中表達低(Stebelová et al, 2010);MT1和 MT2在人胃腸道肌間神經叢、腸上皮和胃腸血管中均有表達,且在大腸上皮表達量最高(S?derquist et al, 2015);鴨腸道 2-125碘褪黑素結合位點密度大小依次為回腸、空腸>十二指腸、結腸>盲腸(Lee et al,1993)。另外,研究表明,Mtnr1介導褪黑激素作用腸道粘膜肌層和腸肌間神經叢發揮免疫功能,抵抗各種病原微生物的侵襲(Esteban-Zubero et al, 2017)。本研究中,Mtnr1基因在腸道不同部位表達量存在差異,暗示Mtnr1在草魚腸道不同部位可能發揮不同的免疫功能,但尚需進一步研究。
值得注意的是,本研究發現,除Mtnr1Alike和Mtnr1C亞型外,Mtnr1Aa、Mtnr1Ab和Mtnr1Ba也在腎臟中高表達,這與金魚中的研究結果類似(Ikegami et al, 2009a),而與歐洲鱸魚(Dicentrarchuslabrax)(Sauzet et al, 2008)鰓、星點東方鲀(Ikegami et al,2009b)神經和垂體表達相異,究其原因可能是亞型基因表達存在物種和組織的特異性。腎臟作為魚體主要的免疫器官,免疫功能相關基因在該組織中呈現高表達(侯吉倫等, 2019),在哺乳動物小鼠的胸腺和腎臟免疫器官中已經證實,Mtnr1mRNA表達量較高(Carrillo-Vicoet al, 2003)。本研究結果顯示,草魚Mntr1亞型mRNA在腎臟中表達量高,暗示 Mtnr1可能與免疫相關。而褪黑激素主要通過與高親和力的特異性Mntr1蛋白結合,隨后啟動信號級聯反應在靶細胞上介導各種生理活動(Klosen et al, 2019)。另外,褪黑激素可以直接作用于機體的免疫活性過程,如Cuesta等(2008)研究發現,金頭鯛(Sparus aurata)注射褪黑激素后,血液中過氧化物酶、吞噬作用和活性氧中間體等活性以及干擾素-1β、主要組織相容性復合體、病毒相關效應和淋巴標記等基因表達顯著上升。同樣地,相對于對照組,注射褪黑激素后中華絨螯蟹(Eriocheir sinensis)血淋巴中超氧化物歧化酶活性顯著增加,丙二醛的含量顯著下調(She et al, 2019)。目前,盡管本研究Mntr1基因在腎臟中高表達,但僅推測Mntr1亞型介導了草魚的免疫反應,而草魚受到病原或褪黑激素刺激后,Mntr1亞型的表達和免疫效應情況并不清楚,需要后續開展實驗進一步探究。
綜上所述,本研究首次在草魚組織中成功克隆得到Mntr1的6種亞型的cDNA序列,并對其序列結構進行了分析,通過對 6種亞型在組織表達模式的研究,初步表明 Mtnr1Aa和 Mtnr1Ba基因與草魚神經調節和免疫反應有關,為今后深入研究Mtnr1Aa和Mtnr1Ba基因在草魚神經調節和免疫功能方面提供了理論依據。
