miR-130b-3p靶定AMPK基因抑制糖饑餓誘導(dǎo)的宮頸癌細胞自噬性死亡
2022-02-13 07:15:42范彩紅楊磊楊瀾穆紅
實用醫(yī)學(xué)雜志 2022年1期
關(guān)鍵詞:水平
范彩紅 楊磊 楊瀾 穆紅
1天津醫(yī)科大學(xué)一中心臨床學(xué)院(天津300192);2天津市第一中心醫(yī)院檢驗科(天津300192)
宮頸癌是導(dǎo)致全球女性癌癥死亡的第二大原因,每年有50 多萬新發(fā)病例,嚴(yán)重?fù)p害著女性的健康,傳統(tǒng)的手術(shù)、放化療方法效果不盡如人意,因此亟待研究發(fā)展出新的治療方法應(yīng)用于臨床[1]。近些年,隨著對腫瘤細胞代謝的深入探究,發(fā)現(xiàn)腫瘤細胞的葡萄糖代謝途徑與正常細胞有顯著差異,基于此提出了“饑餓療法”的概念[2]。營養(yǎng)剝奪成為治療腫瘤的一種手段,但研究發(fā)現(xiàn),在葡萄糖缺乏等應(yīng)激條件下,癌細胞可以利用靈活的代謝程序維持生存能力[3],這使得營養(yǎng)剝奪治療腫瘤的療效不理想,因此探索其耐受機制,強化腫瘤治療效果極為重要。
自噬是細胞通過溶酶體降解細胞內(nèi)容物,回收并利用的機制[4-5]。一方面,自噬通常被看作是饑餓等應(yīng)激條件下細胞的一種重要存活機制,在一定適當(dāng)?shù)乃酱龠M細胞生存[6];然而另一方面,自噬是細胞死亡的一個閾值,強制過度激活超過自噬閾值會導(dǎo)致細胞死亡[7],例如凋亡相關(guān)因子Par-4 誘導(dǎo)人惡性膠質(zhì)瘤細胞自噬性細胞死亡[8]。自噬性細胞死亡(autophagic cell death,ACD)是一種由自噬介導(dǎo)的細胞死亡形式,具有獨特的形態(tài)學(xué)特征,主要表現(xiàn)為自噬體和自溶體在細胞質(zhì)中的積累。細胞死亡沒有表現(xiàn)出凋亡或壞死樣細胞死亡的特征,可以通過測量質(zhì)膜通透性來評估,并且可通過自噬抑制劑或自噬相關(guān)蛋白基因敲除加以抑制[9]。因此,在營養(yǎng)缺乏刺激下,促進過度自噬并激活A(yù)CD 似乎是一種新的癌癥治療策略。
微小RNA(microRNA,miRNA)是一類長度約為22~25 個核苷酸的小RNA,它們與腫瘤發(fā)生發(fā)展密切相關(guān)[10],并且也是調(diào)控細胞內(nèi)自噬相關(guān)信號通路的重要因子[11]。目前,生酮飲食聯(lián)合化療治療腫瘤,能顯著增加化療藥物藥效并減少副作用[12],但仍存在選擇性差、易產(chǎn)生耐藥性的問題,然而基于miRNA的治療靶向性強、精準(zhǔn)度高,能夠更加有效治療腫瘤[13]。但目前關(guān)于miR-130b-3p與饑餓療法的聯(lián)合應(yīng)用以及機制研究鮮有報道。本研究發(fā)現(xiàn)糖饑餓可以誘導(dǎo)宮頸癌細胞發(fā)生自噬,miR-130b-3p 在防止細胞發(fā)生過度自噬過程發(fā)揮重要作用。本研究新發(fā)現(xiàn)為全面闡明miRNA 調(diào)控腺苷單磷酸激活激酶(AMP-activated protein kinase,AMPK)相關(guān)自噬的分子機制提供了有用的線索,也為探索miRNA輔助饑餓療法下宮頸癌的治療提供了一些新的見解。
1 材料與方法
1.1 材料宮頸癌Siha、Hela 細胞(美國ATCC 公司);細胞培養(yǎng)箱(美國Thermo Fisher 公司);熒光定量PCR 儀(瑞士Roche 公司);流式細胞儀(美國BDSciences 公司);凝膠成像分析儀(美國Alpha Innotech 公司);LipofectamineTM3000(美國Thermo Fisher 公司);GFP-LC3B 質(zhì)粒(湖南科愛醫(yī)療器械有限公司);雷帕霉素、SBI-0206965(美國默克公司);p-ULK1 抗體(美國Abcam 有限公司);兔抗人腺苷單磷酸激活激酶(AMP-activated protein kinase,AMPK)、LC3 抗體(美國CST 公司);β-actin、HRP 標(biāo)記羊抗兔抗體(美國Proteintech 公司);PI 檢測試劑盒(天津三箭生物技術(shù)股份有限公司)。
1.2 實驗方法
1.2.1 細胞培養(yǎng)及轉(zhuǎn)染宮頸癌細胞在含有8%~10%胎牛血清、1%青霉素-鏈霉素雙抗的MEM 細胞培養(yǎng)基中培養(yǎng),于37 ℃,5% CO2的細胞培養(yǎng)箱中孵育。取指數(shù)期細胞計數(shù)后按照所需密度種于孔板中,待細胞貼壁后,利用LipofectaminTM3000 將miR-130b-3p模擬物、NC、miR-130b-3p抑制物、抑制物NC、pcDNA3.1::AMPK、pcDNA3.1、pcDNA3.1::AMPK-wtUTR 和pcDNA3.1::AMPK-mutUTR、GFPLC3B 質(zhì)粒分別轉(zhuǎn)染細胞。
1.2.2 實驗分組根據(jù)處理條件,以及轉(zhuǎn)染物質(zhì)的不同分為以下幾組:正常糖含量組與低糖組、低糖+DMSO 組與低糖+雷帕霉素組、NC 組與模擬物組、抑制物NC 組與抑制物組、pEGFP::wtUTR組和pEGFP::mutUTR 組、抑制物NC+SBI 組與抑制物+SBI 組、pcDNA3.1 組與pcDNA3.1::AMPK 組、pcDNA3.1+SBI 組與pcDNA3.1::AMPK+SBI 組。
1.2.3 實時熒光定量PCR(RT-qPCR)用TB Green 進行RT-qPCR 檢測miR-130b-3p、內(nèi)參U6、AMPK mRNA、內(nèi)參β-actin mRNA 表達水平,按公式:Folds=2-ΔΔCt,△△Ct=(Ct1-Ct2)-(Ct3-Ct4),計算出每組細胞中基因的相對表達水平。miR-130b-3p引物序列U:5'-AGTGCAGGGTCCGAGGTAT-3',D:5'-TGCAATGATGAAAGGGCAT-3';U6 引物序列F:5'-CGCTTCGGCAGCACAT-3',R:5'-ATTTGCGTGTCATCCTTGC-3';ampk 引物序列F:5'-GCGACAAGCCCACCTGAT-3',R:5'-ATTTGCGTGTCATCCTTGC-3';β-actin 引物序列F:5'-AGTTGCGTTACACCCTTTCTTG-3',R:5'-TGTCACCTTCACCGTTCCAGT-3'。
1.2.4 過表達質(zhì)粒載體的構(gòu)建按照商品使用說明(TaKaRa,日本),使用AMPK-RT,通過逆轉(zhuǎn)錄反應(yīng)合成AMPK mRNA 編碼區(qū)的cDNA,再依次使用兩對引物akF1、akR1 和akF2、akR2 通過PCR 擴增含有Kozak 序列和AMPK 基因編碼序列的DNA 片段,純化后酶切DNA 片段,連接到pcDNA3.1 載體的KpnI-XhoI 克隆位點,使用T4 DNA 連接酶構(gòu)建pcDNA3.1::AMPK載體。AMPK-RT:5'-AACAAAGAGGGAGGGAT-3';AMPK F1:5'-CCGCCACGATGCGCAGACTCAGTTC-3';R1:5'-TTGCATGGCGCTTTCTGTTTATTGTGCAAG-3';AMPK F2:5'-CGGAATTCGCCGCCACGATGCG-3';AMPK R2:5'-CCGCTCGAGGCATGGCGCTTTCTGT-3'
1.2.5 Western blotPBS 清洗6 孔板中細胞后,加入150 μL RIPA 裂解液,靜置30 min 后,用刮板刮取細胞蛋白,BCA 試劑盒進行蛋白定量,取總蛋白30 μg 上樣,配制10 %的SDS-PAGE 凝膠分離,電泳后轉(zhuǎn)膜,5%脫脂牛奶封閉2 h,隨后分別加入一抗,4 ℃過夜后,TBST 清洗4 次,之后加入二抗室溫孵育1 h,TBST 清洗4 次,最后化學(xué)顯色液顯色,Image J 分析灰度值。
1.2.6 細胞死亡率檢測消化收集各組細胞,加入600 μL 固定液清洗細胞,100 μL 固定液重懸細胞,加入PI 染液5 μL,避光孵育5 min,立即上流式細胞儀檢測,實驗重復(fù)3 次。
1.2.7 GFP-LC3B實驗細胞(1×105/孔)接種在爬片上并在12 孔板中過夜培養(yǎng),使用LipofectamineTM3000 轉(zhuǎn)染GFP-LC3B 質(zhì)粒6 h 后,更換新鮮培養(yǎng)基48 h 后刺激,用4%多聚甲醛固定細胞20 min,PBS沖洗3 次。使用熒光顯微鏡拍攝圖像。為了定量,在每個實驗條件下至少計數(shù)100 個細胞中GFPLC3B 斑點,并使用Image J 中的共定位工具對綠色和紅色熒光共定位進行定量。
1.2.8 熒光報告基因?qū)嶒濼argetScan、miRDB、PicTar、DIANA 在線數(shù)據(jù)庫預(yù)測到miR-130b-3p 與AMPK mRNA3'-UTR 存在靶向結(jié)合位點,根據(jù)生物信息學(xué)預(yù)測結(jié)果,將AMPK mRNA3'-UTR 模擬物及突變體,分別構(gòu)建嵌入到熒光報告載體pEGFP 下游BamHI-XhoI克隆位點上,該模擬物合成由上海吉瑪制藥技術(shù)有限公司完成。將構(gòu)建好的重組載體質(zhì)粒分別與miR-130b-3p 模擬物及NC 共轉(zhuǎn)染宮頸癌細胞,48 h后分別測定熒光強度,實驗重復(fù)3次。
1.3 統(tǒng)計學(xué)方法實驗數(shù)據(jù)利用GraphPadPrism 8.0 軟件分析并繪圖。在對照組中獲得的數(shù)據(jù)的平均值被定義為1 或100%,以使不同試驗組中表達的靶基因的水平標(biāo)準(zhǔn)化。用SPSS 16.0 軟件對實驗數(shù)據(jù)的正態(tài)分布進行了驗證。不成對t檢驗用于對數(shù)據(jù)進行統(tǒng)計評估,P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié)果
2.1 低糖誘導(dǎo)宮頸癌細胞自噬和耐受與正常糖含量組相比,低糖組細胞死亡率差異無統(tǒng)計學(xué)意義(均P>0.05,圖1A);低糖顯著誘導(dǎo)GFP-LC3B斑點的形成(均P<0.001,圖1B);與正常糖含量組相比,低糖組LC3 蛋白水平均顯著性增加、p-ULK1(Ser757)蛋白水平顯著降低(均P<0.001,圖1C)。
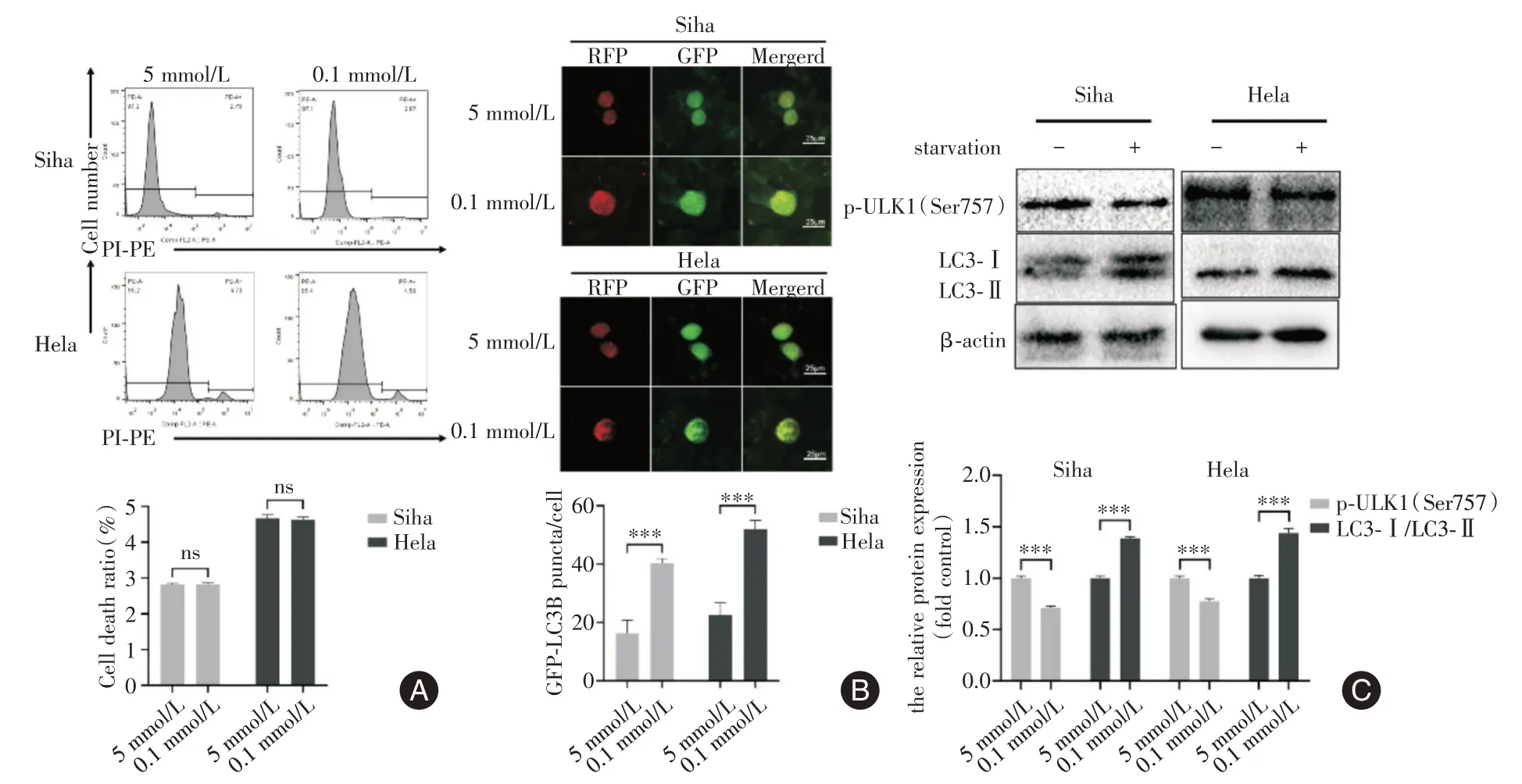
圖1 低糖條件下對宮頸癌細胞死亡、自噬水平的影響Fig.1 Effects of low glucose on death and autophagy of cervical cancer cells
2.2 過度自噬誘導(dǎo)宮頸癌細胞死亡與低糖+DMSO組相比,低糖+雷帕霉素組宮頸癌細胞死亡水平率顯著升高(均P<0.001,圖2)。

圖2 過度自噬誘導(dǎo)宮頸癌細胞死亡Fig.2 Excessive autophagy induces death of cervical cancer cells
2.3 低糖條件下miR-130b-3p 和AMPK 變化情況低糖條件下,AMPK 蛋白水平呈現(xiàn)顯著性降低趨勢(均P<0.01,圖3A);RT-qPCR 實驗結(jié)果顯示,與正常糖含量組相比,低糖組AMPK mRNA 顯著性下降;miR-130b-3p 表達水平顯著性升高(均P<0.05,圖3B-C)。

圖3 低糖條件下miR-130b-3p、AMPK mRNA 和蛋白水平變化Fig.3 Changes of miR-130b-3p,AMPK mRNA and protein levels under low glucose conditions
2.4 miR-130b-3p 直接靶定AMPK 基因Target-Scan、miRDB、Pic Tar、DIANA 在線數(shù)據(jù)庫預(yù)測結(jié)果(圖4A)顯示,miR-130b-3p 與AMPK mRNA 3'-UTR存在靶向結(jié)合位點,提示AMPK可能是miR-130b-3p的靶基因。與NC 組相比,模擬物組AMPK mRNA水平以及蛋白水平顯著降低(均P<0.05,圖4B);而轉(zhuǎn)染抑制物后則趨勢相反(均P<0.05)。熒光報告基因?qū)嶒灲Y(jié)果證實,轉(zhuǎn)染miR-130b-3p 模擬物可顯著抑制pEGFP::wtUTR組的熒光強度(P<0.05),而對于pEGFP::mutUTR 的熒光強度無顯著影響(P>0.05,圖4C)。
2.5 miR-130b-3p抑制物誘導(dǎo)ACD與抑制物NC組相比(圖5A),抑制物組死亡率顯著高于對照組(均P<0.001);抑制物NC+SBI 組與抑制物+SBI 組細胞死亡率差異無統(tǒng)計學(xué)意義(均P>0.05)。轉(zhuǎn)染miR-130b-3p抑制物顯著增加GFP-LC3B斑點細胞的形成(P<0.01,圖5B);與抑制物NC組相比,抑制物組LC3蛋白水平顯著升高(均P<0.05,圖5C);p-ULK1(Ser757)蛋白水平呈現(xiàn)顯著降低趨勢(均P<0.05)。
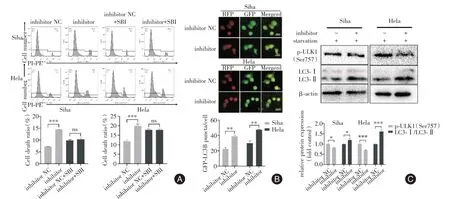
圖5 miR-130b-3p 抑制物對細胞自噬和死亡的影響Fig.5 Effects of miR-130b-3p inhibitor on autophagy andcell death
2.6 AMPK 誘導(dǎo)ACD與pcDNA3.1 組相比(圖6A),pcDNA3.1::AMPK 組的死亡率顯著增加(均P<0.01);pcDNA3.1+SBI 組與pcDNA3.1::AMPK+SBI 組細胞死亡率差異無統(tǒng)計學(xué)意義(均P>0.05)。過表達AMPK 顯著誘導(dǎo)細胞內(nèi)GFP-LC3B斑點的形成(P<0.01,圖6B);與pcDNA3.1組相比,pcDNA3.1::AMPK 組LC3 蛋白顯著升高(均P<0.01,圖6C);p-ULK1(Ser757)顯著性降低(P<0.01)。
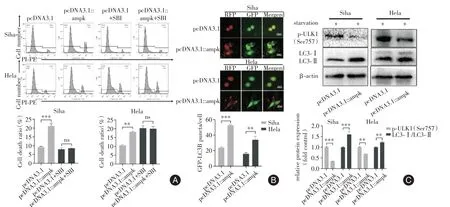
圖6 過表達AMPK 對細胞自噬和死亡的影響Fig.6 Effects of overexpression of AMPK on cell autophagy and death
3 討論
研究表明不同類型腫瘤細胞的低糖耐受能力是不同的,低糖條件雖然可以加速腦膠質(zhì)瘤細胞的死亡[14],但并不會誘導(dǎo)肝癌細胞發(fā)生死亡[15],為了檢測宮頸癌細胞是否存在低糖耐受現(xiàn)象,本研究采用低糖刺激,結(jié)果發(fā)現(xiàn)宮頸癌Siha 和Hela 細胞死亡率無差異,這證明其也存在低糖耐受現(xiàn)象。有研究報道[16]稱在葡萄糖缺乏的應(yīng)激過程中,細胞通過抑制mTOR 表達從而激活自噬。低糖條件下,在宮頸癌細胞中,本研究也發(fā)現(xiàn)自噬相關(guān)蛋白LC3、GFP-LC3B 斑點顯著增加,這些結(jié)果表明,葡萄糖饑餓同樣會引起宮頸癌細胞的自噬。然而,眾所周知自噬是把“雙刃劍”,適當(dāng)程度的自噬,有助于細胞生存,但過度的自噬就會導(dǎo)致細胞的死亡,即ACD 的發(fā)生。在多骨髓瘤細胞中,高水平的自噬可導(dǎo)致半胱氨酸蛋白酶非依賴性自噬細胞死亡[17]。為進一步驗證過度自噬是否同樣誘導(dǎo)宮頸癌細胞ACD,本研究采用饑餓與雷帕霉素聯(lián)合刺激,結(jié)果發(fā)現(xiàn)宮頸癌細胞LC3 蛋白、GFP-LC3B 斑點顯著增加,細胞死亡率也顯著增加,這也說明了過度自噬同樣可以誘導(dǎo)宮頸癌細胞ACD 的發(fā)生。
葡萄糖缺乏等應(yīng)激條件下,會激活多種上游信號通路參與自噬調(diào)節(jié),包括AMPK/mTOR。AMPK是真核細胞中最重要的分子能量傳感器之一,其在維持能量穩(wěn)態(tài)中起關(guān)鍵作用[18],一方面,AMPK磷酸化激活位點Thr172 活化后,通過抑制ULK1 抑制性磷酸化位點Ser757 的活化促進自噬誘導(dǎo);另一方面,AMPK 負(fù)性調(diào)節(jié)mTOR 誘導(dǎo)自噬,然而AMPK 的持續(xù)激活導(dǎo)致糖酵解和葡萄糖氧化的解偶聯(lián),增加胞漿蛋白水平和酸中毒,并引起過度自噬至有害水平,誘導(dǎo)ACD 的發(fā)生[19]。本研究中,低糖條件下,AMPK mRNA 以及蛋白表達量水平降低表明AMPK 可能參與到糖饑餓下自噬性細胞死亡的調(diào)節(jié),為更直觀地證明關(guān)于AMPK 作用猜想,筆者過表達AMPK 發(fā)現(xiàn)其促進低糖下細胞自噬,誘導(dǎo)細胞死亡,并且這種死亡可以被自噬抑制劑SBI-0206965 所逆轉(zhuǎn),因此AMPK 的過表達可以誘導(dǎo)宮頸癌細胞過度自噬進而導(dǎo)致ACD,低表達水平的AMPK 抑制了宮頸癌細胞ACD。
miRNAs 通過靶向不同信號通路中的基因,與ACD 調(diào)節(jié)過程密切相關(guān)。研究發(fā)現(xiàn),轉(zhuǎn)染miR-20b抑制劑后,抑制METTL3 的表達,從而誘導(dǎo)了內(nèi)皮細胞的ACD[20],miR-22-3p 的過度表達抑制凋亡蛋白IAP 的表達,誘導(dǎo)自噬細胞引起的死亡[21]。本研究中,低糖條件下,miR-130b-3p 水平升高,這表明miR-130b-3p 可能也參與到糖饑餓下自噬性細胞死亡的調(diào)節(jié)。因此轉(zhuǎn)染miR-130b-3p 抑制物后,降低miR-130b-3p 水平,結(jié)果發(fā)現(xiàn)宮頸癌細胞自噬顯著升高,細胞死亡率顯著增加,并且這種死亡可以被自噬抑制劑SBI-0206965 所逆轉(zhuǎn),因此抑制miR-130b-3p 的表達可以誘導(dǎo)宮頸癌細胞過度自噬促進細胞ACD 的發(fā)生。此外,miRNAs 通過與靶基因3'UTR 區(qū)域互補結(jié)合來調(diào)節(jié)其表達水平。有趣的是,在本研究中利用生物信息學(xué)軟件預(yù)測到AMPK 3'UTR 中miR-130b-3p 的假定靶定位點。為進一步驗證二者靶定關(guān)系,分別轉(zhuǎn)染了miR-130b-3p 模擬物和抑制物,miR-130b-3p 模擬物降低AMPK mRNA 和蛋白水平,miR-130b-3p 抑制物則趨勢相反,這些結(jié)果說明二者有靶定關(guān)系。miR-130b-3p 模擬物導(dǎo)致編碼野生型3'UTR 的DNA 序列的報告基因表達水平下降,但沒有3'UTR 序列或編碼突變型3'UTR 的序列的報告基因水平?jīng)]有明顯下降,這進一步說明了miR-130b-3p 與AMPK具有直接靶定關(guān)系。
總之,本研究發(fā)現(xiàn)miR-130b-3p 下調(diào)AMPK 的表達,抑制過度自噬誘導(dǎo)的ACD,實現(xiàn)宮頸癌細胞低糖下的生存。本研究強調(diào)了miRNA 在宮頸癌治療中的重要作用,為輔助饑餓療法治療宮頸癌提供新的參考。目前,饑餓療法還未廣泛應(yīng)用臨床治療,miRNA 作為治療靶點治療腫瘤仍僅限于少數(shù)癌癥種類,故miRNA 輔助饑餓療法仍有很長的路要走。
猜你喜歡
美與時代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
