棉花色素腺體發(fā)育相關(guān)基因GhERF105-like的克隆與表達(dá)分析
2022-02-15 02:48:26錢(qián)玉源王廣恩權(quán)月偉崔淑芳米換房李俊蘭
華北農(nóng)學(xué)報(bào) 2022年6期
張 曦,錢(qián)玉源,王廣恩,劉 祎,權(quán)月偉,崔淑芳,米換房,李俊蘭
(1.河北省農(nóng)林科學(xué)院 棉花研究所,農(nóng)業(yè)部黃淮海半干旱區(qū)棉花生物學(xué)與遺傳育種重點(diǎn)實(shí)驗(yàn)室,國(guó)家棉花改良中心河北分中心,河北 石家莊 050051;2.邯鄲市農(nóng)業(yè)科學(xué)院,河北 邯鄲 056001)
棉花是關(guān)系國(guó)計(jì)民生重要的經(jīng)濟(jì)作物。棉花纖維是重要的紡織工業(yè)原料,棉籽油、棉籽蛋白、棉籽殼等棉副產(chǎn)品在食品、養(yǎng)殖、工業(yè)等領(lǐng)域具有巨大應(yīng)用潛力[1]。但由于棉花根、莖、葉、花、種子等器官中含有棉酚等毒性物質(zhì),能夠?qū)ρ堋⑸窠?jīng)、內(nèi)臟及生殖系統(tǒng)等造成損害,嚴(yán)重限制了棉副產(chǎn)品的綜合利用[2]。棉花色素腺體(Pigment gland)又稱(chēng)棉酚腺體,是貯藏棉酚及其衍生物質(zhì)的細(xì)胞間隙[3]。棉酚等酚醌類(lèi)物質(zhì)由色素腺體中的分泌細(xì)胞產(chǎn)生[4]。通常棉酚含量與棉花色腺體數(shù)量正相關(guān)[5]。敲除色素腺體形成相關(guān)基因可顯著降低棉酚含量[6-10]。棉花色素腺體也是選育低酚棉品種的重要指示性狀[11]。因此,發(fā)掘和利用棉花色素腺體發(fā)育相關(guān)基因,解析其對(duì)腺體發(fā)育和棉酚合成的調(diào)控機(jī)制,可以為高效開(kāi)展低酚棉育種提供基因資源和理論依據(jù)。
研究表明,棉花色素腺體形成主要由gl1~gl6基因位點(diǎn)控制,一些基因位點(diǎn)還存在相應(yīng)的復(fù)等位基因[12]。gl1位點(diǎn)控制下胚軸、莖稈、葉柄和棉鈴無(wú)腺體性狀。Zang等[13]圖位克隆了gl1位點(diǎn)候選基因GoSPGF。GoSPGF編碼1個(gè)GRAS(GAI-RGA-and-SCR)轉(zhuǎn)錄因子,沉默該基因后植株莖稈無(wú)腺體、棉酚含量顯著降低。gl2、gl3以及Gl2e位點(diǎn)控制植株各部位無(wú)腺體性狀[14]。Ma等[6]和Cheng等[15]分別精細(xì)定位了顯性無(wú)腺體基因Gl2e,篩選到1個(gè)bHLH-MYC轉(zhuǎn)錄因子在有腺體和無(wú)腺體材料間顯著差異表達(dá)。Ma等[6]克隆了該基因,命名為GoPGF(Gossypium pigment gland formation gene)。沉默GoPGF基因后植株表現(xiàn)全株無(wú)腺體。在gl2gl3突變體中,GoPGF基因發(fā)生單核苷酸突變,造成翻譯提前終止,植株呈隱性無(wú)腺體表型。GoPGF在A亞組中的同源基因CGF3(Cotton Gland Formation 3)編碼區(qū)存在1個(gè)轉(zhuǎn)座子插入突變,在D亞組中同源基因的啟動(dòng)子區(qū)存在大量變異[7]。在子葉色素腺體延緩發(fā)生的比克氏棉(G.bickiiProkh)中,GoPGF基因也與腺體形成相關(guān)[16]。除GoPGF和GoSPGF外,還發(fā)現(xiàn)一些腺體發(fā)育相關(guān)基因在不同程度上影響棉花腺體形成及棉酚代謝。澳洲棉(G.australe)GRAS蛋白基因GauGRAS1影響莖和葉柄中色素腺體形成及棉酚合成[9]。MYB轉(zhuǎn)錄因子CGP1(Cotton Gland Pigmentation 1)在色素腺體中特異表達(dá)。敲除CGP1基因植株呈現(xiàn)類(lèi)似無(wú)腺體表型,但CGP1并不影響腺體結(jié)構(gòu)和腺體密度,而是影響棉酚合成與腺體中的色素沉積。CGP1可與GoPGF互作形成異二聚體,進(jìn)而調(diào)控棉花棉酚合成及其他代謝[8]。CGF1編碼1個(gè)bHLH轉(zhuǎn)錄因子,沉默CGF1基因?qū)е轮仓晟叵袤w數(shù)量減少78%。CGF2基因編碼NAC轉(zhuǎn)錄因子,沉默CGF2影響色素密度、腺體結(jié)構(gòu)及腺體發(fā)育。沉默CGF1、CGF2及CGF3基因均可顯著降低植株棉酚含量[7]。此外,一些基因如GhWD40[17]、RanBP2[18]、GhNAC201[19]等,在腺體發(fā)育過(guò)程中或不同腺體類(lèi)型棉花間顯著差異表達(dá),可能與棉花色調(diào)腺體形成或棉酚合成密切相關(guān)。
APETALA2/ethylene response factor(AP2/ERF)轉(zhuǎn)錄因子存在于所有植物中,參與植物營(yíng)養(yǎng)生長(zhǎng)、生殖生長(zhǎng)、生物和非生物脅迫應(yīng)答等生命活動(dòng)[20]。研究發(fā)現(xiàn),AP2/ERF基因在有腺體和無(wú)腺體棉間或色素腺體發(fā)育過(guò)程中顯著差異表達(dá),暗示AP2/ERF轉(zhuǎn)錄因子在棉花色素腺體發(fā)育或棉酚代謝過(guò)程中也發(fā)揮重要作用[6-8,21-22]。姜鵬飛等[23]對(duì)有腺體棉Z12及其隱性無(wú)腺體近等基因系Z12YW的幼嫩葉片進(jìn)行轉(zhuǎn)錄組測(cè)序,篩選到13個(gè)差異表達(dá)轉(zhuǎn)錄因子,其中7個(gè)為AP2/ERF轉(zhuǎn)錄因子。Wu等[10]克隆1個(gè)在有腺體和無(wú)腺體近等基因系間顯著差異表達(dá)的AP2/ERF家族基因GhERF105(GH_A12G2166),沉默GhERF105后新生葉片葉脈的腺體數(shù)量減少,新生莖部仍密生腺體。葉片棉酚含量水平顯著降低。河北省農(nóng)林科學(xué)院棉花研究所品種資源研究室對(duì)有腺體棉中棉所12和無(wú)腺體棉中棉所12顯性無(wú)腺體幼胚進(jìn)行轉(zhuǎn)錄組測(cè)序,篩選到1個(gè)顯著差異表達(dá)的AP2/ERF轉(zhuǎn)錄因子(Gh_A12G1784),該基因在其他不同腺體類(lèi)型棉花材料間也顯著差異表達(dá)[7-8]。
本研究克隆了Gh_A12G1784基因,命名為GhERF105-like(GenBank登錄號(hào):MZ239205),并對(duì)其基因序列及表達(dá)模式進(jìn)行了分析,旨在為探析AP2/ERF轉(zhuǎn)錄因子在棉花色素腺體發(fā)育中的生物學(xué)功能提供數(shù)據(jù)基礎(chǔ),為開(kāi)展低酚棉分子育種提供候選基因。
1 材料和方法
1.1 植物材料與處理
有色素腺體棉花品種中棉所12(中12)、遼棉7號(hào)、邯5158及無(wú)色素腺體棉花品種/系中棉所12顯性無(wú)腺體(中12顯無(wú))、遼棉7號(hào)顯性無(wú)腺體(遼7顯無(wú))和新陸中1號(hào)由中國(guó)農(nóng)業(yè)科學(xué)院棉花研究所提供;無(wú)色素腺體棉花品種邯無(wú)198由邯鄲市農(nóng)業(yè)科學(xué)院提供。
中12和中12顯無(wú)在大田種植,常規(guī)管理。掛牌標(biāo)記自交時(shí)間,分別取16,26,35 DPA(Days post anthesis)幼胚,液氮速凍,-80 ℃保存,用于驗(yàn)證GhERF105-like在2個(gè)材料幼胚發(fā)育過(guò)程中的表達(dá)變化。
所有供試有腺體和無(wú)腺體棉品種/系在光照培養(yǎng)室中用營(yíng)養(yǎng)缽?fù)僚嘀恋?片真葉完全展開(kāi)時(shí)取葉片,液氮速凍,-80 ℃保存,用于分析基因在有腺體和無(wú)腺體棉間表達(dá)水平的差異。
中12和中12顯無(wú)分別在種子萌發(fā)期、苗期和開(kāi)花期取樣,液氮速凍,-80 ℃保存,用于分析基因的時(shí)空表達(dá)模式。種子萌發(fā)期:種子胚根長(zhǎng)約1 cm左右時(shí)剝?nèi)ッ拮褮ぃ≌C拮选C缙冢河没舾裉m營(yíng)養(yǎng)液種植水培苗,在第2片真葉完全展開(kāi)時(shí)分別取子葉、根、莖及葉片。開(kāi)花期:取大田種植棉花的當(dāng)天盛開(kāi)花和長(zhǎng)約1 cm的幼蕾。
中12在光照培養(yǎng)室中用營(yíng)養(yǎng)缽培養(yǎng)至第2片真葉完全展開(kāi)時(shí)分別噴施脫落酸(Abscisic acid,ABA,100 μmol/L)、茉莉酸甲酯(Methyl jasmonate,MeJA,400 μmol/L)和油菜素內(nèi)酯(Brassinosteroid,BR,1 μmol/L),于處理后0,3,6 h分別取葉片;乙烯利(400 mg/L)處理0,3,6,9,12,24 h分別取葉片。中12水培至第2片真葉完全展開(kāi)時(shí)分別用含200 mmol/L NaCl或20% PEG-6000的營(yíng)養(yǎng)液處理幼苗,于處理0,3,6,9 h分別取葉片。所有樣品液氮速凍,-80 ℃保存,用于分析基因的表達(dá)變化。
1.2 棉花總RNA提取和cDNA第一鏈合成
使用RNAprep Pure多糖多酚植物總RNA提取試劑盒(天根生化科技(北京)有限公司)提取樣品總RNA,瓊脂糖凝膠電泳檢測(cè)其質(zhì)量,-80 ℃保存?zhèn)溆谩J褂肞rimeScriptTMⅡ 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA第一鏈,-20 ℃保存?zhèn)溆谩?/p>
1.3 基因表達(dá)分析
用NCBI Primer-BLAST工具(https://www.ncbi.nlm.nih.gov/)設(shè)計(jì)GhERF105-like的熒光定量PCR引物(qGhERF105-like-F:5′-CATGGAAACTCCA
CCACCCT-3′和qGhERF105-like-R:5′-AACCGGGTTT
GGGCTTATGT-3′),以Histone3(qHiston3F:5′-CGGT
GGTGTGAAGAAGCCTCAT-3′和qHiston3R:5′-AATT
TCACGAACAAGCCTCTGGAA-3′)[6]為內(nèi)參基因。qRT-PCR使用2×Sybr Green qPCR Mix(北京艾德萊生物科技有限公司),反應(yīng)體系及反應(yīng)參數(shù)參照試劑盒說(shuō)明書(shū)。每個(gè)樣品3次重復(fù),用2-ΔΔCt法計(jì)算基因相對(duì)表達(dá)量。
1.4 基因序列克隆
根據(jù)TM-1(NAU-NBI_v1.1)序列,使用NCBI Primer-BLAST工具設(shè)計(jì)克隆引物(fGhERF105-like-F:5′-CACCACGCTCCCCAATTTCCACT-3′和fGhERF105-like-R:5′-GCTTAGGCTTTAGGCATCCCATC
C-3′)。以中12葉片cDNA第一鏈為模板進(jìn)行PCR擴(kuò)增。PCR反應(yīng)體系:cDNA模板1 μL,正反向引物(10 μmol/L)各1 μL,10×LA PCR Buffer Ⅱ(Mg2+plus) 5 μL,2.5 mmol/L dNTP Mixture 8 μL,5 U/μL TaKaRa LA Taq 0.5 μL,補(bǔ)充ddH2O至總體積50 μL。PCR反應(yīng)條件:94 ℃預(yù)變性5 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸2 min,30個(gè)循環(huán);72 ℃總延伸10 min。PCR產(chǎn)物與pGEM-T Easy載體(Promega)連接后轉(zhuǎn)化大腸桿菌DH5α菌株。陽(yáng)性克隆送蘇州金唯智生物科技有限公司測(cè)序。
1.5 序列生物信息學(xué)分析
將獲得的GhERF105-like序列在GenBank數(shù)據(jù)庫(kù)中進(jìn)行BlastX比對(duì),利用MEGA 7.0比對(duì)GhERF105-like同源蛋白序列和構(gòu)建系統(tǒng)進(jìn)化樹(shù)。使用在線(xiàn)網(wǎng)站分析GhERF105-like蛋白理化性質(zhì)(http://www.expasy.org/tools/protparam.html)、二級(jí)結(jié)構(gòu)(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)、三級(jí)結(jié)構(gòu)(http://www.sbg.bio.ic.ac.uk/phyre2/html/)、蛋白功能域(http://smart.embl-heidelberg.de/)、蛋白磷酸化位點(diǎn)(http://www.cbs.dtu.dk/services/NetPhos/)及蛋白氨基酸序列的保守基序(http://meme-suite.org)等。
2 結(jié)果與分析
2.1 GhERF105-like在有腺體和無(wú)腺體棉花間的表達(dá)分析
在有腺體棉幼胚中,色素腺體在14~20 DPA開(kāi)始發(fā)育[7,24]。GhERF105-like在無(wú)腺體棉中棉所12顯性無(wú)腺體(中12顯無(wú))幼胚中表達(dá)量極低,在幼胚發(fā)育不同時(shí)期差異不顯著;在有腺體棉中棉所12(中12)的16,26,35 DPA幼胚中表達(dá)量逐步極顯著增加(P<0.01),且在26,35 DPA極顯著高于中12顯無(wú)(圖1)。GhERF105-like在不同有腺體棉品種/系葉片中的表達(dá)量不同,且均顯著高于顯性或隱性無(wú)腺體棉品種/系(P<0.05)(圖2)。表明GhERF105-like與棉花色素腺體發(fā)育或棉酚合成密切相關(guān)。

不同大寫(xiě)字母表示1%水平差異顯著(P<0.01)。圖7同。Different capital letters are significantly different at 1% level(P<0.01).The same as Fig.7.

不同小寫(xiě)字母表示5%水平差異顯著(P<0.05)。圖7同。Different letters are significantly different at 5% level(P<0.05).The same as Fig.7.
2.2 GhERF105-like基因的克隆與序列分析
以中12葉片cDNA為模板,利用引物fGhERF105-like-F和fGhERF105-like-R,用RT-PCR的方法獲得目的片段(圖3)。
GhERF105-like編碼區(qū)為711 bp,編碼236個(gè)氨基酸(圖4)。GhERF105-like蛋白的分子質(zhì)量為26.3 ku,理論等電點(diǎn)pI為7.72,為不穩(wěn)定性(不穩(wěn)定系數(shù)70.44)親水(總平均親水性-0.611)蛋白。SOPMA預(yù)測(cè)GhERF105-like的二級(jí)結(jié)構(gòu)包括22.88%的α-螺旋,13.14%的延伸鏈,3.81%的β-轉(zhuǎn)角和60.17%的無(wú)規(guī)則卷曲。GhERF105-like在第91—155位氨基酸存在1個(gè)AP2結(jié)構(gòu)域(圖4)。用NetPhos 3.1預(yù)測(cè)潛在磷酸化位點(diǎn),表明GhERF105-like有23個(gè)絲氨酸、7個(gè)蘇氨酸和2個(gè)酪氨酸潛在修飾位點(diǎn)。其中第92位氨基酸處的酪氨酸磷酸化位點(diǎn)、第121,124位氨基酸處的蘇氨酸磷酸化位點(diǎn)及第142位氨基酸處的絲氨酸磷酸化位點(diǎn)位于AP2結(jié)構(gòu)域內(nèi)(圖4)。三維結(jié)構(gòu)預(yù)測(cè)表明,GhERF105-like 87~208位氨基酸序列與乙烯應(yīng)答因子ERF096蛋白序列高度匹配(52%覆蓋度,100%置信度)。根據(jù)ERF096晶體結(jié)構(gòu)預(yù)測(cè)GhERF105-like的三維結(jié)構(gòu)見(jiàn)圖5。

彩色箭頭方向?yàn)镹→C末端。模型維度 X.40.874;Y.48.799;Z.44.154。Image coloured by rainbow N→C terminus.Model X.40.874;Y.48.799;Z.44.154.
2.3 GhERF105-like同源蛋白進(jìn)化及保守基序分析
將GhERF105-like與其他植物中同源AP2/ERF蛋白構(gòu)建系統(tǒng)進(jìn)化樹(shù),結(jié)果表明,GhERF105-like與棉屬及其他雙子葉植物中同源蛋白的序列相似度高于單子葉植物中同源蛋白(圖6-A)。進(jìn)一步對(duì)這些蛋白的保守基序進(jìn)行分析,可以發(fā)現(xiàn)GhERF105-like在棉屬黃褐棉(TYJ42243.1)、達(dá)爾文氏棉(TYH24288.1)、雷蒙德氏棉(XP 012439108.1)、海島棉(KAB2053713.1)中蛋白基序組成一致。基序1和基序2在不同物種中高度保守,除基序10外其他基序保守性也較高。單子葉植物中AP2/ERF蛋白基序組成與雙子葉植物明顯不同(圖6-B)。
2.4 GhERF105-like的時(shí)空表達(dá)模式
GhERF105-like在中12不同組織中的表達(dá)量除在莖中顯著高于中12顯無(wú)外(P<0.05),在其他組織中均極顯著高于中12顯無(wú)(P<0.01),且在2個(gè)棉花材料的子葉和蕾中豐度均較高,表明GhERF105-like表達(dá)水平與植株是否存在腺體密切相關(guān),并且傾向于在幼嫩組織中高豐度表達(dá)(圖7-A)。在中12葉片中,GhERF105-like受BR誘導(dǎo)上調(diào)表達(dá),處理6 h的表達(dá)量顯著高于處理0 h對(duì)照。在ABA處理下,GhERF105-like表達(dá)量先下調(diào)再上調(diào),處理6 h顯著高于處理3 h。在MeJA處理下,GhERF105-like下調(diào)表達(dá),在處理3 h達(dá)極顯著水平(P<0.01),之后表達(dá)略有上升(圖7-B)。噴施Eth后,GhERF105-like也呈下調(diào)表達(dá)趨勢(shì),在處理3,9 h的表達(dá)極顯著低處理0 h對(duì)照(圖7-C)。NaCl和PEG脅迫處理下GhERF105-like下調(diào)表達(dá)。NaCl處理3 h,GhERF105-like表達(dá)與對(duì)照相比極顯著下降(P<0.01),之后表達(dá)量略有上升后又降低,在處理9 h再次極顯著低于對(duì)照(P<0.01)。GhERF105-like在PEG處理下的表達(dá)變化趨勢(shì)與NaCl處理相同,但在處理3,9 h與對(duì)照的差異僅達(dá)顯著水平(P<0.05)。以上結(jié)果表明,在本試驗(yàn)濃度下,ABA和BR誘導(dǎo)GhERF105-like上調(diào)表達(dá),而MeJA、Eth、NaCl、PEG等因素對(duì)GhERF105-like的表達(dá)起抑制作用。
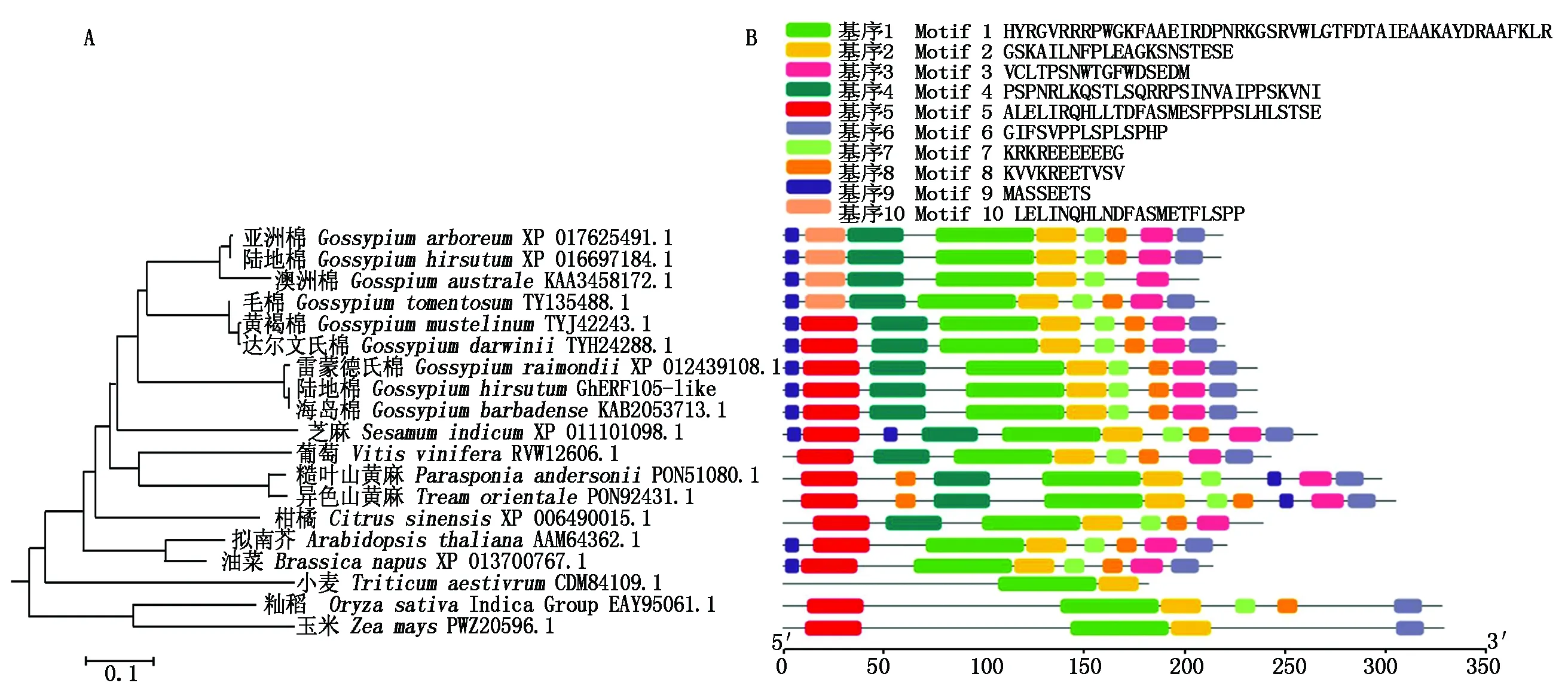
圖6 GhERF105-like同源蛋白的進(jìn)化樹(shù)(A)及保守基序(B)分析Fig.6 Phylogenetic(A)and motif composition(B)analysis of GhERF105-like homologous proteins

A.GhERF105-like在有腺體棉中棉所12和中棉所12顯性無(wú)腺體同一組織部位的表達(dá)差異;B、C.中棉所12中GhERF105-like受植物激素誘導(dǎo)的表達(dá)模式;D.中棉所12中GhERF105-like非生物脅迫下的表達(dá)模式。A.Differential expression of GhERF105-like in same tissue of two cotton germplasms;B,C.Expression patterns of GhERF105-like under hormone treatment in Zhongmiansuo 12;D.Expression patterns of GhERF105-like under NaCl or PEG treatment in Zhongmiansuo 12.
3 結(jié)論與討論
棉酚的生物毒性限制了棉籽、棉株等棉副產(chǎn)品的綜合利用[1]。棉花色素腺體的有無(wú)及數(shù)量與棉酚含量顯著相關(guān),通常腺體有無(wú)影響棉酚代謝,但棉酚合成并不影響腺體形成[5-6]。研究棉花色素腺體發(fā)育相關(guān)基因及其對(duì)棉酚合成的調(diào)控機(jī)制,可為低酚棉分子育種提供理論依據(jù)和基因資源。
AP2/ERF轉(zhuǎn)錄因子超家族主要存在于植物中,在植物生長(zhǎng)發(fā)育、脅迫應(yīng)答、物質(zhì)代謝等生命活動(dòng)中發(fā)揮重要作用[20]。轉(zhuǎn)錄組分析檢測(cè)到AP2/ERF轉(zhuǎn)錄因子在不同腺體類(lèi)型材料間差異表達(dá)[6-8,22-23]。陸地棉GhERF105基因影響葉片等部分器官腺體形成及棉酚合成[10]。表明AP2/ERF轉(zhuǎn)錄因子也參與調(diào)控棉花色素腺體發(fā)育及棉酚代謝。本研究中,AP2/ERF轉(zhuǎn)錄因子GhERF105-like(Gh_A12G1784)基因在不同有腺體棉品種中的表達(dá)及在有腺體棉中12不同器官中的表達(dá)均顯著或極顯著高于無(wú)腺體棉。Gh_A12G1784在其他研究中也因在有腺體和無(wú)腺體棉花間的顯著差異表達(dá)而被作為候選基因[7-8]。以上結(jié)果表明,GhERF105-like與棉花色素腺體形成或棉酚代謝密切相關(guān)。
GhERF105-like在擬南芥中的同源基因ERF105主要參與冷害[25]、強(qiáng)光[26]、病害[27]等應(yīng)答途徑。系統(tǒng)進(jìn)化分析表明,擬南芥ERF102~ERF105基因非常相近,其對(duì)激素的應(yīng)答、生物學(xué)功能等也相似[25]。ERF102在病原毒素刺激下通過(guò)ETH和JA路徑參與生物脅迫應(yīng)答[28]。ERF105通過(guò)ABA路徑在抵御丁香假單胞桿菌(Pseudomonassyringae)過(guò)程中發(fā)揮作用[27]。ERF103~ERF105通過(guò)影響色氨酸衍生次生代謝物合成顯著抑制真菌增殖,但不影響植株生長(zhǎng)[29]。超表達(dá)ZmERF105基因增強(qiáng)玉米大斑病抗性及PR(Pathogenesis-related)基因表達(dá)[30]。棉花受病蟲(chóng)害侵染后色素腺體密度和棉酚含量會(huì)發(fā)生改變[31]。本研究中,GhERF105-like應(yīng)答ABA、BR、Eth、MeJA等激素及NaCl和PEG脅迫處理,可能通過(guò)這些激素介導(dǎo)的信號(hào)路徑參與調(diào)控腺體形成、棉酚代謝或脅迫應(yīng)答反應(yīng)。
ERF102還參與病原毒素刺激下的細(xì)胞程序性死亡(Programmed cell death,PCD)[28]。伴隨陸地棉色素腺體發(fā)育,中心分泌細(xì)胞逐漸溶解形成腔體,可觀察到細(xì)胞壁膨脹、DNA斷裂和降解、液泡中出現(xiàn)自溶體等PCD現(xiàn)象[32]。本研究中,GhERF105-like在中12 16 DPA幼胚中的表達(dá)量極低,隨著色素腺體的發(fā)育表達(dá)量逐步極顯著增加,其在Stoneville 7A有腺體及其無(wú)腺體近等基因系間也有相似的表達(dá)趨勢(shì)[7],表明GhERF105-like可能在腺體發(fā)育中后期或棉酚代謝過(guò)程中發(fā)揮作用。腺體發(fā)育過(guò)程中的PCD現(xiàn)象也發(fā)生在腺體發(fā)育的中后期,推測(cè)GhERF105-like也可能參與PCD過(guò)程。
綜上所述,GhERF105-like與棉花色素腺體發(fā)育或棉酚代謝密切相關(guān),可能通過(guò)ABA、BR、Eth、MeJA等路徑參與調(diào)控腺體發(fā)育、棉酚代謝及生物/非生物脅迫應(yīng)答。
