硫化氫通過NCOA4介導的鐵自噬調(diào)控鐵死亡而減輕CSE誘導的肺泡上皮細胞損傷*
2022-02-16 02:59:54潘子涵蔣思敏陳亞紅
中國病理生理雜志 2022年1期
關鍵詞:檢測
王 瑩, 廖 莎, 潘子涵, 蔣思敏, 樊 靜, 陳亞紅, 張 靜
(北京大學第三醫(yī)院呼吸與危重癥學科,北京 100191)
慢性阻塞性肺疾病(簡稱慢阻肺),是世界范圍內(nèi)導致人類死亡的主要原因之一,吸煙被認為是慢阻肺發(fā)生發(fā)展的主要因素。煙草煙霧含有顆粒、自由基和活性物質等約4 500種化學成分,提示可能有多種損傷及細胞死亡機制參與CSE 介導的細胞損傷[1]。氣道上皮細胞是肺臟與外界進行氣體交換的第一道屏障,近期研究表明,CSE 誘發(fā)的氣道上皮細胞鐵死亡參與了慢阻肺的進程[2]。鐵死亡(ferropto?sis)是Dixon等[3]在2012年發(fā)現(xiàn)的一種新型細胞死亡形式,其特征是鐵介導的Fenton 反應產(chǎn)生活性氧(re?active oxygen species,ROS)引起的脂質過氧化,活性鐵是其基礎。而細胞內(nèi)不穩(wěn)定的鐵可通過鐵自噬(ferritinophagy)途徑產(chǎn)生。鐵自噬是一種新型的自噬類型,此過程的發(fā)生依賴于特異性受體核受體輔激活因子4(nuclear receptor coactivator 4,NCOA4),它能將鐵蛋白運輸?shù)阶允尚◇w,而后與溶酶體融合,使鐵蛋白降解為活性鐵[4]。因此,抑制鐵自噬-鐵死亡可能會對保護氣道上皮細胞進而阻斷慢阻肺進展提供新的思路。
硫化氫(hydrogen sulfide,H2S)是繼一氧化氮和一氧化碳之后發(fā)現(xiàn)的第3 種新型內(nèi)源性氣體信號分子。本課題組長期圍繞H2S 在呼吸系統(tǒng)的作用進行研究,發(fā)現(xiàn)其具有抗氧化、抗炎和抗凋亡等多種功能[5]。然而,H2S 是否具有抗肺泡上皮細胞鐵死亡的作用,尚屬未知。因此,本項工作通過使用CSE 作為刺激物建立肺泡上皮損傷細胞模型,圍繞鐵自噬-鐵死亡信號通路,探討H2S 在肺泡上皮細胞中的作用,以期為慢阻肺的防治提供參考資料。
材料和方法
1 細胞株和主要試劑
人源肺泡上皮細胞BEAS-2B 購自ATCC 上海細胞庫;RPMI-1640 培養(yǎng)液購自HyClone;澳洲胎牛血清購自Gibco;香煙(白沙牌,含焦油11 mg、煙堿0.9 mg 及煙氣一氧化碳12 mg);硫氫化鈉(sodium hydro?sulfide,NaHS)購自Sigma-Aldrich;CCK-8 試劑盒購自日本同仁公司;ROS 檢測試劑盒購自北京索萊寶公司;BODIPY 581/591 C11 脂質ROS(lipid ROS,Lip ROS)檢測試劑盒購自Invitrogen;丙二醛(malondial?dehyde,MDA)和谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)檢測試劑盒購自上海碧云天生物技術有限公司;鐵離子檢測試劑盒、抗NCOA4 抗體(1∶1 000)、抗鐵蛋白重鏈1(ferritin heavy chain 1,F(xiàn)TH1)抗體(1∶1 000)、抗環(huán)加氧酶2(cyclooxygenase 2,COX2)抗體(1∶1 000)、抗谷胱甘肽過氧化物酶4(glutathione peroxidase 4,GPX4)抗體(1∶1 000)和抗GAPDH 抗體(1∶5 000)均購自Abcam;HRP 標記的山羊抗兔IgG(1∶10 000)購自北京中杉金橋公司。
2 主要方法
2.1 藥物準備NaHS 作為H2S 供體用PBS 溶解配成10 mmol/L儲存液,?80 ℃冰箱凍存?zhèn)溆茫贑SE加入前30 min 預處理。CSE 需現(xiàn)用現(xiàn)配,1 支香煙對應10 mL 無胎牛血清培養(yǎng)液,點燃香煙,包式吸收管內(nèi)緩慢勻速抽吸,再經(jīng)0.22 μm 濾器除菌,而后使用分光光度計在320 nm 處測定液體吸光度(absorbance,A),范圍在4.00±0.05內(nèi)可用,即為100%的CSE。
2.2 細胞培養(yǎng)BEAS-2B 細胞用含有10% 胎牛血清、1×105U/L 青霉 素和100 mg/L 鏈 霉素的RPMI-1640 培養(yǎng)液置于37 ℃、5% CO2的培養(yǎng)箱中培養(yǎng),取對數(shù)生長期細胞進行實驗。除特殊說明外,其他實驗均將4×105個BEAS-2B 細胞接種于6 孔板中,細胞完全貼壁后,按照分組給予藥物刺激處理24 h。
2.3 CCK-8 法檢測BEAS-2B 細胞活力取對數(shù)期的BEAS-2B 細胞以每孔5×103個的密度接種于含100 μL 完全培養(yǎng)液的96 孔板中,細胞完全貼壁后用不同濃度的CSE或NaHS處理24 h。棄培養(yǎng)液后PBS洗3 次,每孔加入含10% CCK-8 檢測液的培養(yǎng)液100 μL,37 ℃孵育2 h后490 nm處使用酶標儀測量A值。
2.4 Fe2+含量檢測藥物處理完成后收集細胞沉淀,加入100 μL裂解液,使用杜恩斯勻漿器冰上勻漿10 次,離心取上清。按照鐵離子檢測試劑盒說明書加入探針,于593 nm 處測得A值,繪制標準曲線并計算鐵離子濃度。
2.5 脂質過氧化分析取對數(shù)期的BEAS-2B 細胞以每孔5×104個的密度接種于共聚焦皿中,細胞完全貼壁后用CSE 或NaHS 處理24 h。棄上清,加入含BODIPY?581/591 C11(7 μmol/L)的無血清培養(yǎng)液,37 ℃孵育30 min,PBS 洗3 次,最后活細胞狀態(tài)下使用共聚焦顯微鏡采集熒光圖片。
2.6 流式細胞術檢測細胞內(nèi)ROS和Lip ROS含量藥物處理完成后收集細胞沉淀,用無血清培養(yǎng)液稀釋DCFH-DA 熒光探針(終濃度為2μmol/L)或BODIPY?581/591 C11 熒光探針(終濃度為7 μmol/L),每孔0.5 mL/孔重懸細胞,37 ℃孵育30 min。吸棄含探針的培養(yǎng)液后用無血清培養(yǎng)液潤洗細胞2次,每個樣品加入無血清培養(yǎng)液150 μL重懸,流式細胞儀上機檢測。
2.7 ELISA 法檢測細胞內(nèi)MDA 及GSH-Px含量藥物處理完成后收集細胞沉淀,細胞裂解液裂解細胞,BCA 蛋白定量試劑盒測定裂解液上清中的蛋白濃度。按照廠家說明書測定檢測裂解液上清中MDA和GSH-Px 的含量。最后繪制標準曲線,計算各自濃度。
2.8 Western blot 法檢測相關蛋白的表達藥物處理完成后收集細胞沉淀,細胞裂解液裂解細胞,使用BCA 蛋白定量試劑盒進行蛋白定量,蛋白變性后存于?20℃。按照40 μg 上樣量上樣,經(jīng)SDS-PAGE 后電轉,5% 脫脂奶粉室溫封閉1 h,加入目的蛋白Ⅰ抗4℃搖床孵育過夜。回收Ⅰ抗,1× TBST 洗3 次每次5 min,再加入Ⅱ抗室溫孵育1 h,TBST 洗3 次后ECL 顯影,應用ImageJ軟件進行灰度分析。
3 統(tǒng)計學處理
采用SPSS 26.0 統(tǒng)計軟件進行數(shù)據(jù)分析。數(shù)據(jù)均采用均數(shù)±標準差(mean±SD)表示。多組間均數(shù)比較采用單因素方差分析(one-way ANOVA),兩兩比較采用LSD-t檢驗。以P<0.05為差異有統(tǒng)計學意義。
結 果
1 不同濃度CSE 刺激對鐵死亡標志物GPX4 及COX2表達的影響
Western blot 檢測結果顯示,隨著CSE 濃度的增加,BEAS-2B 細胞內(nèi)COX2 表達水平呈上升趨勢,且濃度大于3% 時,表達量加劇,增加至1.5 倍以上(P<0.01);而GPX4蛋白表達水平與CSE濃度呈反比,在5%濃度時開始顯著降低(P<0.01),見圖1。
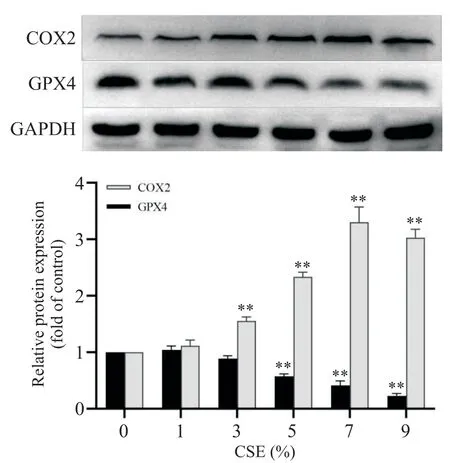
Figure 1.Effects of treatment with different concentrations of CSE for 24 h on the protein expression of COX2 and GPX4 in BEAS-2B cells.The expression of target pro?teins was measured by Western blot.Mean±SD. n=4.**P<0.01 vs control(0% CSE)group.圖1 不同濃度CSE 處理24 h 對BEAS-2B 細胞中COX2 和GPX4蛋白表達的影響
2 NaHS及CSE處理對BEAS-2B細胞活力的影響
不同濃度的CSE 處理24 h 后BEAS-2B 細胞活力的變化如圖2A 所示,CSE 對BEAS-2B 細胞活力的影響具有濃度依賴性,且當濃度在5% 時細胞活力顯著下降至60% 左右(P<0.01)。結合文獻報道[2],選擇5%作為后續(xù)的CSE刺激濃度。
進一步檢測NaHS對肺泡上皮細胞的保護作用,結果如圖2B 所示。與CSE 刺激組相比,NaHS(200和400 μmol/L)預處理可明顯增強CSE刺激后BEAS-2B 細胞的活力(P<0.01),并呈劑量依賴性。值得注意的是,單純NaHS處理細胞也可增強BEAS-2B細胞活力(P<0.05)。
3 NaHS 及CSE 處理對BEAS-2B 細胞Fe2+含量的影響
接下來提取細胞內(nèi)Fe2+評估CSE及NaHS預處理對細胞內(nèi)活性鐵的影響,結果如圖3 所示。與空白對照組相比,CSE 處理可以使細胞內(nèi)Fe2+含量呈現(xiàn)約2 倍的升高(P<0.01);而相比于CSE 單純刺激組,NaHS 預處理組Fe2+含量有所下降,且高劑量組差異有統(tǒng)計學意義(P<0.05)。
4 NaHS 及CSE 處理對細胞內(nèi)ROS 及Lip ROS 的產(chǎn)生和氧化-抗氧化系統(tǒng)的影響
通過對Lip ROS、ROS、MDA 及GSH-Px 含量進行檢測評估氧化還原系統(tǒng),結果如圖4所示。單純CSE處理能刺激綠光的大量產(chǎn)生,使細胞處于氧化狀態(tài),而NaHS預處理可減輕CSE介導的細胞氧化程度。

Figure 2.Effects of CSE and NaHS on the viability of BEAS-2B cells.A:effects of treatment with different concentrations of CSE for 24 h on BEAS-2B cell viability;B:effects of 30 min of NaHS pretreatment on the viability of BEAS-2B cells treated with 5% CSE for 24 h.Mean±SD. n=4.*P<0.01 vs control(0% CSE or 0% CSE+0 μmol/L NaHS)group;##P<0.01 vs 5%CSE group.圖2 CSE和NaHS對BEAS-2B細胞活力的影響
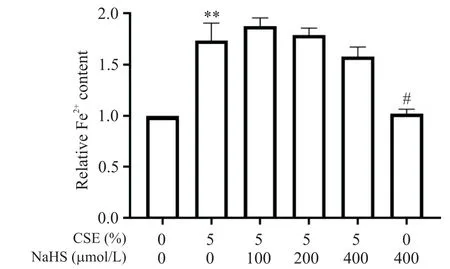
Figure 3.Cellular iron content in BEAS-2B cells of each group.The cells were treated with 5% CSE for 24 h with or without 30 min of NaHS pretreatment.Mean±SD. n=4.**P<0.01 vs control(0% CSE+0 μmol/L NaHS)group;#P<0.05 vs 5% CSE group.圖3 各組BEAS-2B細胞鐵含量
除此之外我們還對細胞內(nèi)的ROS 及Lip ROS 進行了流式細胞術檢測,結果如圖5A、B 所示。與空白對照組相比,CSE 處理使細胞內(nèi)Lip ROS及總ROS的水平增加2 倍以上;而給予NaHS 預處理后Lip ROS及ROS 顯著減少(P<0.05),且與NaHS 濃度呈正相關。此外,MDA 結果也與ROS 趨勢相符,見圖5C。GSH-Px是體內(nèi)重要的抗氧化系統(tǒng)組分之一,如圖5D所示,CSE 引起的GSH-Px 水平降低可被NaHS 拮抗,且在高濃度時具有統(tǒng)計學意義(P<0.05)。
5 NaHS 及CSE 處理對細胞內(nèi)鐵自噬-鐵死亡相關蛋白表達情況的影響
為了探究NaHS 抵抗CSE 引起的細胞損傷的機制,我們對細胞內(nèi)鐵自噬-鐵死亡相關蛋白進行了檢測,結果如圖6 所示。與對照組相比,單純CSE 可導致鐵自噬-鐵死亡相關蛋白NCOA4、FTH1和COX2的水平顯著升高(P<0.01),同時GPX4 表達水平顯著降低(P<0.05);而不同濃度NaHS可顯著逆轉CSE單純刺激的結果,使鐵自噬-鐵死亡相關蛋白NCOA4、FTH1 及COX2 的表達水平呈現(xiàn)不同程度的下降,GPX4蛋白有所上升(P<0.05),且這些蛋白的變化具有濃度依賴性。
討 論
慢阻肺是危害我國民眾健康的常見病,隨著疾病的進展,可能會導致呼吸衰竭,并且造成嚴重的經(jīng)濟和社會負擔,嚴重影響生命安全和生活質量[6]。慢阻肺的發(fā)病以活性氧為基礎,活性氧通過激活促炎轉錄,誘導細胞死亡、細胞衰老和DNA 損傷,損害抗蛋白酶防御能力等途徑,促進和增強氧化應激級聯(lián)反應,進而導致氣道炎癥及肺氣腫[7]。香煙煙霧、空氣污染和生物學煙霧是肺部氧化應激的主要外源性來源,并且多種細胞損傷方式參與這些危險因素的致病過程[8],鐵死亡是其中一種細胞死亡形式。鐵死亡是一種由脂質過氧化以鐵依賴的方式引發(fā)的細胞死亡形式。已有文獻顯示CSE 具有激活鐵死亡的作用。例如,給予CSE 刺激后,血管平滑肌細胞出現(xiàn)鐵死亡特異性標志物PTGS2 上調(diào)、脂質過氧化增加和谷胱甘肽消耗增加等鐵死亡特征性變化[9]。而在肺部,Park 等[10]對支氣管上皮細胞進行CSE 暴露研究,結果表明CSE 可能通過內(nèi)質網(wǎng)應激和誘導缺氧引起線粒體動態(tài)穩(wěn)態(tài)紊亂進而誘導支氣管上皮細胞鐵死亡。此外,目前最有說服力的一項研究來源于Yoshida 等[2],他們的實驗結果顯示CSE 可通過NCOA4 介導的鐵自噬促進不穩(wěn)定的鐵積累,導致人肺上皮細胞脂質過氧化和鐵死亡,參與了慢阻肺的發(fā)病。而我們的實驗結果顯示,隨著CSE 濃度的增加,鐵死亡標志蛋白COX2表達水平逐漸升高,GPX4逐漸降低,且單純CSE 刺激可導致細胞內(nèi)總ROS、Lip ROS、脂質過氧化產(chǎn)物MDA 及活性鐵離子顯著增加,抗氧化酶GSH-Px 的水平顯著降低,以及鐵自噬相關蛋白NCOA4 和FTH1 的大量增加,證實CSE介導的肺泡上皮細胞損傷機制中確實有鐵死亡參與。
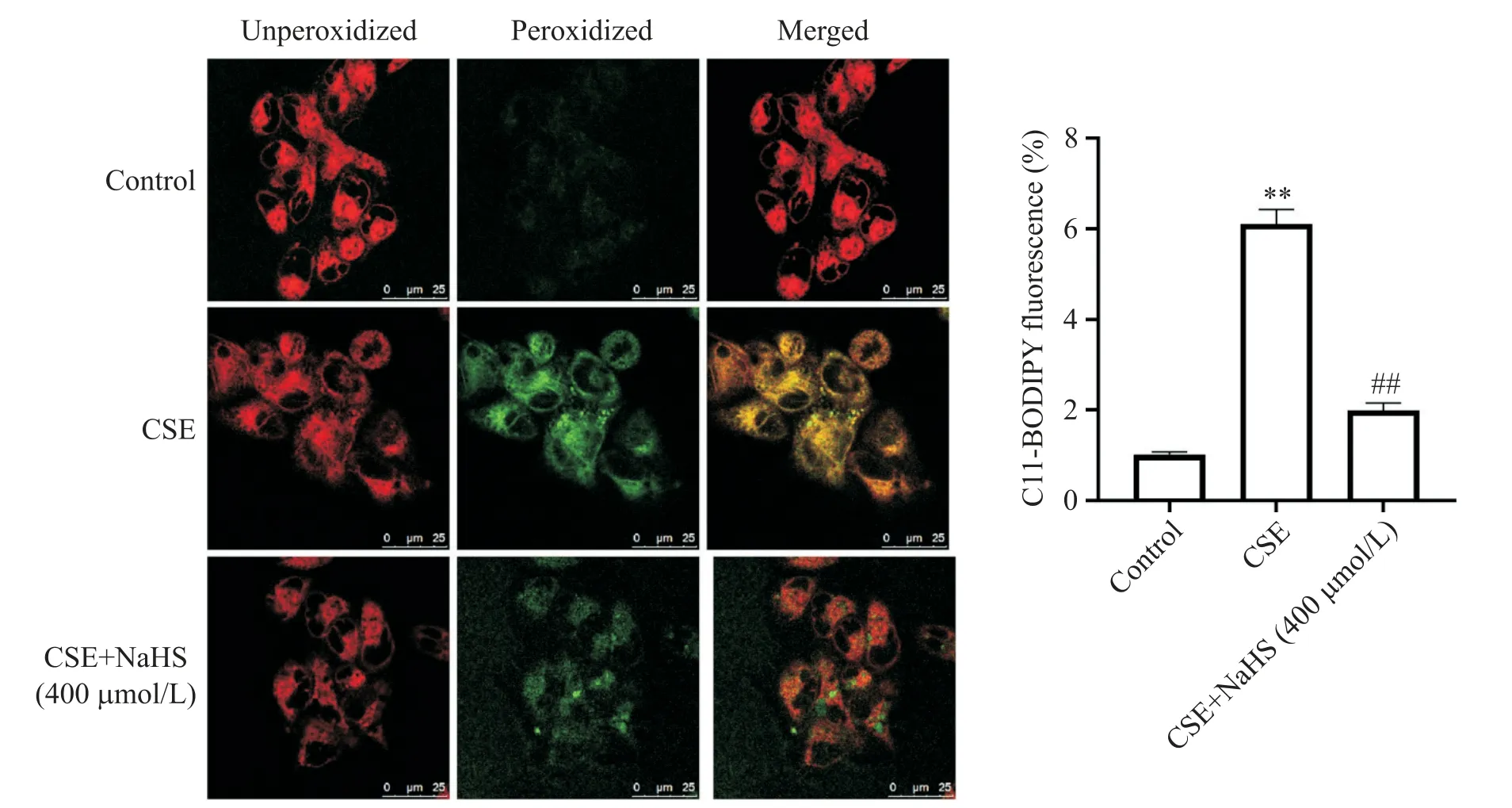
Figure 4.Effects of NaHS and CSE on membrane lipid peroxidation in BEAS-2B cells measured by confocal fluorescence microscopy,using C11-BODIPY as a probe(scale bar=50 μm).The cells were treated with 5% CSE for 24 h with or without 30 min of NaHS(400 μmol/L)pretreatment.BODIPY?581/591 C11 reagent is a sensitive fluorescent reporter for lipid peroxida?tion,and the fluorescence shifts from red to green upon oxidation in live cells.Mean±SD. n=4.**P<0.01 vs control group;##P<0.05 vs CSE group.圖4 共聚焦熒光顯微鏡檢測NaHS及CSE處理對BEAS-2B細胞膜脂質過氧化的影響
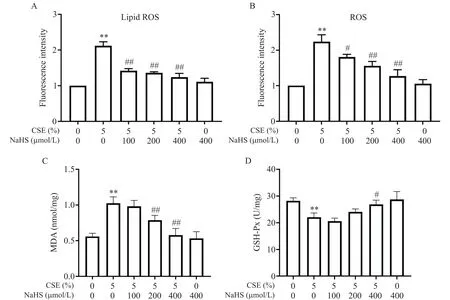
Figure 5.Effects of NaHS and CSE on the levels of lipid ROS(A),ROS(B),MDA(C)and GSH-Px(D)in BEAS-2B cells.The cells were treated with 5% CSE for 24 h with or without 30 min of NaHS pretreatment.Mean±SD. n=4.**P<0.01 vs control(0% CSE+0 μmol/L NaHS)group;#P<0.05,##P<0.01 vs 5% CSE group.圖5 NaHS和CSE對BEAS-2B細胞中脂質ROS、ROS、MDA和GSH-Px水平的影響

Figure 6.Effects of NaHS and CSE on the expression levels of ferritinophagy-ferroptosis-related proteins in BEAS-2B cells detected by Western blot.The cells were treated with 5% CSE for 24 h with or without 30 min of NaHS pretreatment.A:the expres?sion of GPX4 and COX2;B:the expression of NCOA4 and FTH1.Mean±SD. n=4.*P<0.05,**P<0.01 vs control(0%CSE+0 μmol/L NaHS)group;#P<0.05,##P<0.01 vs 5% CSE group.圖6 Western blot檢測NaHS和CSE對BEAS-2B細胞中鐵自噬-鐵死亡相關蛋白表達的影響
自噬是在應激、饑餓或缺氧期間維持細胞穩(wěn)態(tài)的生理過程[11]。2014 年Mancias 等[12]通過定量蛋白質組學鑒定NCOA4 為介導鐵自噬的特異性受體,提出鐵自噬的概念,由此將自噬與鐵死亡聯(lián)系在一起。越來越多的研究也證實鐵自噬具有調(diào)控鐵死亡的作用。例如,Masaldan 等[13]的研究顯示,衰老細胞中的鐵積累與鐵自噬能力受損和鐵死亡受抑制有關。Latunde-Dada等[14]的研究顯示,半胱氨酸缺失介導的鐵蛋白降解通過NCOA4 介導的自噬途徑釋放鐵。Gao 等[15]報道,通過抑制自噬或抑制NCOA4 的表達可抑制鐵自噬的發(fā)生,從而導致鐵蛋白酶相關的細胞不穩(wěn)定鐵和Lip ROS的積聚。基于此,在慢阻肺中抑制鐵自噬進而抑制鐵死亡的策略有待研究。
H2S 是一種有臭雞蛋氣味的內(nèi)源性氣體信號分子。在本研究中,H2S處理能明顯提高CSE 暴露下肺泡上皮細胞的活力,對細胞具有明顯的保護作用,這與先前對不同細胞進行H2S 預處理的研究結果一致[16-18]。且目前已有文獻報道H2S 可通過抑制鐵死亡發(fā)揮抗損傷作用,例如H2S 能通過調(diào)節(jié)Sirt6 而抑制炎癥,降低PFC 和BV2 細胞的鐵死亡,緩解1 型糖尿病小鼠焦慮樣和抑郁樣行為[19]。胱硫醚γ-裂解酶/H2S 系統(tǒng)可作為預防骨骼肌鐵死亡的靶點[20]。氧化還原失衡是鐵死亡的核心機制,脂質過氧化是鐵死亡發(fā)生過程中最關鍵的一步,而抗氧化是H2S重要的防護機制[21]。基于這些理論,我們推測H2S抗CSE介導的細胞損傷的機制可能與調(diào)控鐵自噬-鐵死亡途徑、抑制細胞內(nèi)活性鐵的產(chǎn)生及脂質過氧化過程有關。
為驗證這一假設,接下來我們對H2S處理后的細胞內(nèi)氧化還原標志物如Lip ROS、ROS、MDA 和GSHPx 水平,細胞內(nèi)活性鐵水平,鐵自噬標志蛋白NCOA4 和FTH1,以及鐵死亡標志蛋白COX2 和GPX4 進行了檢測。與預期結果一致,H2S 處理能夠顯著增加抗氧化酶GSH-Px 及GPX4 的水平,同時抑制NCOA4、FTH1 和COX2 的表達及活性鐵的釋放,減少ROS、Lip ROS 和MDA 的產(chǎn)生,且NaHS 濃度越高效果越顯著,證實H2S抗CSE誘導的損傷作用至少部分是通過抑制鐵自噬、抵抗鐵死亡信號通路來實現(xiàn)的。
綜上所述,本研究說明H2S能通過減輕細胞內(nèi)的脂質過氧化失衡和鐵超載而減輕CSE 誘導的肺泡上皮細胞鐵死亡,這可能是通過抑制NCOA4 介導的鐵自噬調(diào)控途徑來實現(xiàn)的。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:36
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:34
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:50
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:48
