FAM83H 及其天然反義轉錄本FAM83H-AS1在肺腺癌和卵巢癌發生發展中的作用
2022-02-16 08:52:46劉夢真孫蓉蓉劉艷華張有為
中國醫藥導報 2022年36期
劉夢真 孫蓉蓉 劉艷華 張有為
1.徐州醫科大學臨床學院,江蘇徐州 221009;2.江蘇省徐州市中心醫院腫瘤內科,江蘇徐州 221009
惡性腫瘤已成為我國居民的主要死因,探索腫瘤發生發展中的關鍵基因具有重要意義[1]。FAM83H 位于染色體8q24.3,是FAM83 家族成員之一,最初因其在牙齒發育中的重要作用被關注,FAM83H 在肝細胞癌、腎癌、胃癌、胰腺癌和宮頸癌等腫瘤中高表達,并促進癌細胞的生長、侵襲和轉移[2-6]。FAM83H-AS1 位于FAM83H 的對側,是其天然反義轉錄本,二者以頭對頭(5’-5’)形式互補重疊。研究表明,FAM83H-AS1 是多種腫瘤潛在的診斷、預后標志物和治療靶點[7-9]。本研究擬在泛癌種水平研究FAM83H 和FAM83H-AS1 在腫瘤發生發展中的生物學作用。
1 材料與方法
1.1 實驗材料
CCK-8 購于Signalway Antibody(CP002);Transwell小室購于Costar(3422);Trizol 購于美國Invitrogen 公司(1596026);逆轉錄試劑盒購于Fermentas(#K1622);SYBR Green PCR 試劑盒(#K0223)、BCA 蛋白定量試劑盒(PICPI23223)、兔抗人FAM83H 多克隆抗體(PA5-55094)購于Thermo;兔抗人GAPDH 單克隆抗體(#5174)購于CST;羊抗兔HRP 標記二抗(A0208)、Annexin V-FITC 細胞凋亡檢測試劑盒(C1036)購于上海碧云天生物技術有限公司。
1.2 生物信息學分析
在線分析FAM83H 基因在惡性腫瘤中的表達及預后,測試數據集來自基因表達譜動態分析(gene expression profiling interactive analysis,GEPIA)工具(http://gepia2.cancer-pku.cn)[10]。
1.3 細胞培養和轉染
siFAM83H 與siFAM83H-AS1 干涉載體由上海基爾頓生物公司構建。A549、SKOV3 細胞均用含10%胎牛血清的RPMI-1640 培養基在37℃,5%CO2培養箱中培養。取對數生長期的細胞進行轉染,分為空載對照組(siNC 組)和實驗組(siFAM83H 組、siFAM83H-AS1組),同時設置空白對照組(control 組),細胞轉染利用脂質體Lipofectamine 2000(美國Invitrogen 公司)介導的方法,轉染48 h 后進行后續實驗。本研究所有實驗步驟均重復3 次。
1.4 Real-time PCR 實驗
胰酶消化并收集各組細胞,Trizol 試劑盒提取總RNA,并對提取的RNA 進行濃度和純度檢測。在37℃60 min、85℃5 min、4℃5 min 的條件下,將RNA 逆轉錄合成cDNA。將合成的cDNA 按Real-time PCR 試劑盒說明書進行擴增,PCR 擴增:95℃10 min(95℃15 s,60℃45 s),40 個循環;熔解曲線:95℃15 s,60℃1 min,95℃15 s,60℃15 s;以GAPDH 作為內參,檢測FAM83H、FAM83H-AS1 的表達。引物序列見表1。

表1 引物序列
1.5 Western blot 檢測
RIPA 法提取細胞總蛋白,BCA 法檢測蛋白濃度。上樣后SDS-PAGE 凝膠電泳,轉膜至PVDF 膜,5%脫脂奶粉室溫封閉1 h,一抗室溫孵育2 h,TBST 洗滌3 次,每次5 min,二抗(1∶1 000)37℃孵育1 h,TBST 洗滌3 次,每次5 min,ECL 系統顯影(GAPDH 作內參)。
1.6 CCK-8 檢測細胞增殖
胰酶消化細胞,鏡下記數將細胞懸液濃度調整為3×104個/ml。按3×103個/孔,將細胞種至96 孔培養板,設置3 個復孔。在轉染0、24、48、72 h 后,加入CCK-8 試劑,酶標儀檢測細胞在450 nm 波長下的吸光度值。
1.7 流式細胞術檢測細胞凋亡
轉染后的細胞用胰酶消化,計數后制備成1×106個/ml的細胞懸液,取1 ml 懸液離心收集細胞,Annexin V-FITC 結合液重懸細胞,4℃避光孵育15 min后,加入碘化丙啶染色液,4℃避光孵育5 min 后,進行流式細胞儀檢測。
1.8 Transwll 檢測細胞遷移
胰酶消化細胞,用含1% FBS 的培養基制備3×105個/ml 的細胞懸液,每個Transwell 小室接種300 μl細胞懸液,下層加入700 μl 的含10% FBS 的培養基,每組細胞設置3 個復孔。培養24 h 后,取出小室,PBS 清洗2 次,甲醛固定10 min,PBS 洗滌2 次,0.5%結晶紫染色30 min,PBS 洗滌3 次,晾干后于顯微鏡下觀察拍照(200×),計數不同視野下的細胞數取平均值。
1.9 統計學方法
采用SPSS 21.0 軟件進行數據分析。計量資料用均數±標準差()表示,比較采用t 檢驗;計數資料用百分率表示,比較采用χ2檢驗。以P<0.05 為差異有統計學意義。
2 結果
2.1 FAM83H、FAM83H-AS1mRNA 在泛癌種水平的表達
GEPIA 在線工具分析結果顯示,FAM83H mRNA在乳腺浸潤癌、胸腺瘤、肺腺癌(lung adenocarcinoma,LUAD)、肺鱗癌、肝細胞癌、胃癌、胰腺癌、結腸癌、膀胱尿路上皮癌、直腸癌、宮頸鱗癌和腺癌、子宮內膜癌和子宮肉瘤、卵巢漿液性囊腺瘤(ovarian serous cystadenocarcinoma,OV)中表達上調;在多形成性膠質細胞瘤、急性髓系白血病和皮膚黑色素瘤中表達下調。FAM83H-AS1 mRNA 在膀胱尿路上皮癌、乳腺浸潤癌、宮頸鱗癌和腺癌、結腸癌、LUAD、肺鱗癌、OV、胰腺癌、前列腺癌、直腸癌、胃癌、子宮內膜癌和子宮肉瘤中表達上調,在急性髓系白血病、皮膚黑色素瘤和睪丸癌中表達下調。見圖1。
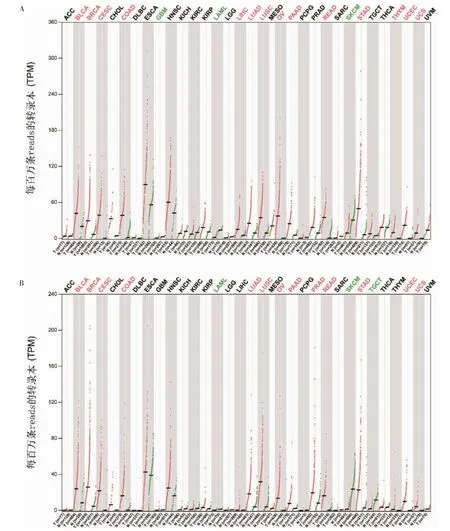
圖1 FAM83H、FAM83H-AS1 mRNA 在泛癌種水平的表達
2.2 FAM83H、FAM83H-AS1 mRNA 表達與LUAD和OV 的預后
在LUAD 中,FAM83HmRNA 高表達組與低表達組生存曲線比較,差異有統計學意義(P<0.05)。見圖2。
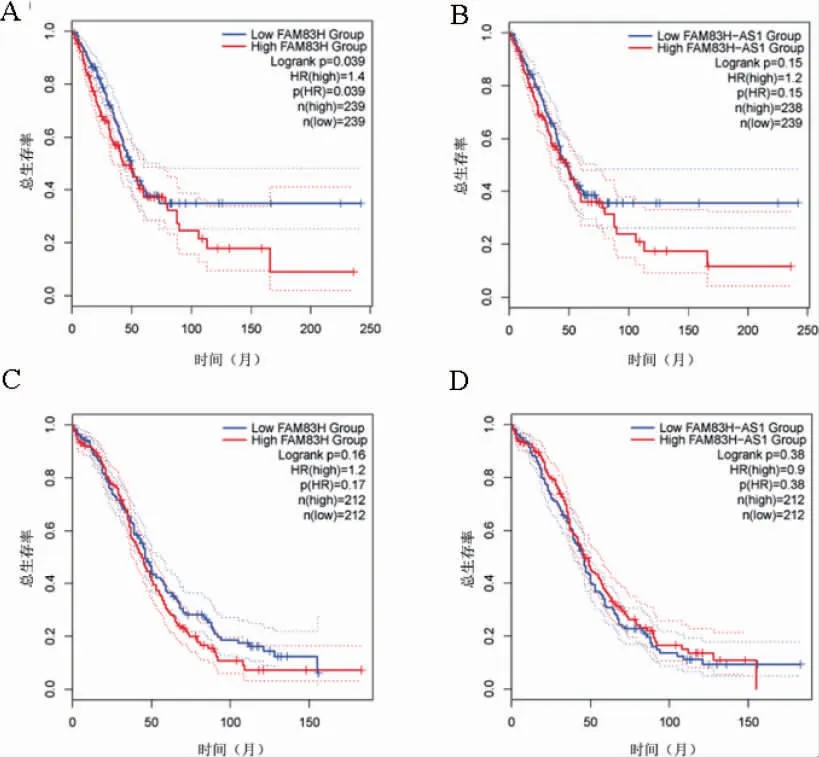
圖2 FAM83H、FAM83H-AS1 mRNA 表達與LUAD 和OV 的預后
2.3 FAM83H、FAM83H-AS1 敲低后各組FAM83H、FAM83H-AS1 表達比較
A549 和SKOV3 細胞中,siFAM83H 組FAM83H mRNA 及蛋白表達低于siNC 組(P<0.05);siFAM83HAS1 組FAM83H-AS1 mRNA 表達低于siNC 組(P<0.05)。見圖3。
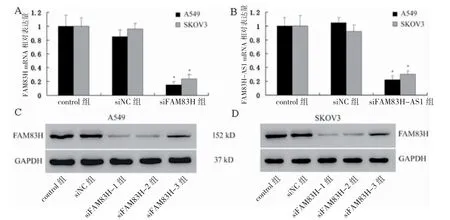
圖3 FAM83H、FAM83H-AS1 敲低后各組FAM83H、FAM83H-AS1 表達比較(n=3)
2.4 FAM83H、FAM83H-AS1 敲低后各組細胞增殖能力比較
A549 和SKOV3 細胞中,siFAM83H 組轉染24、48、72 h 后細胞增殖能力低于siNC 組(P<0.05)。siFAM83H-AS1 組轉染24、48、72 h 后細胞增殖能力低于siNC 組(P<0.05)。見圖4。
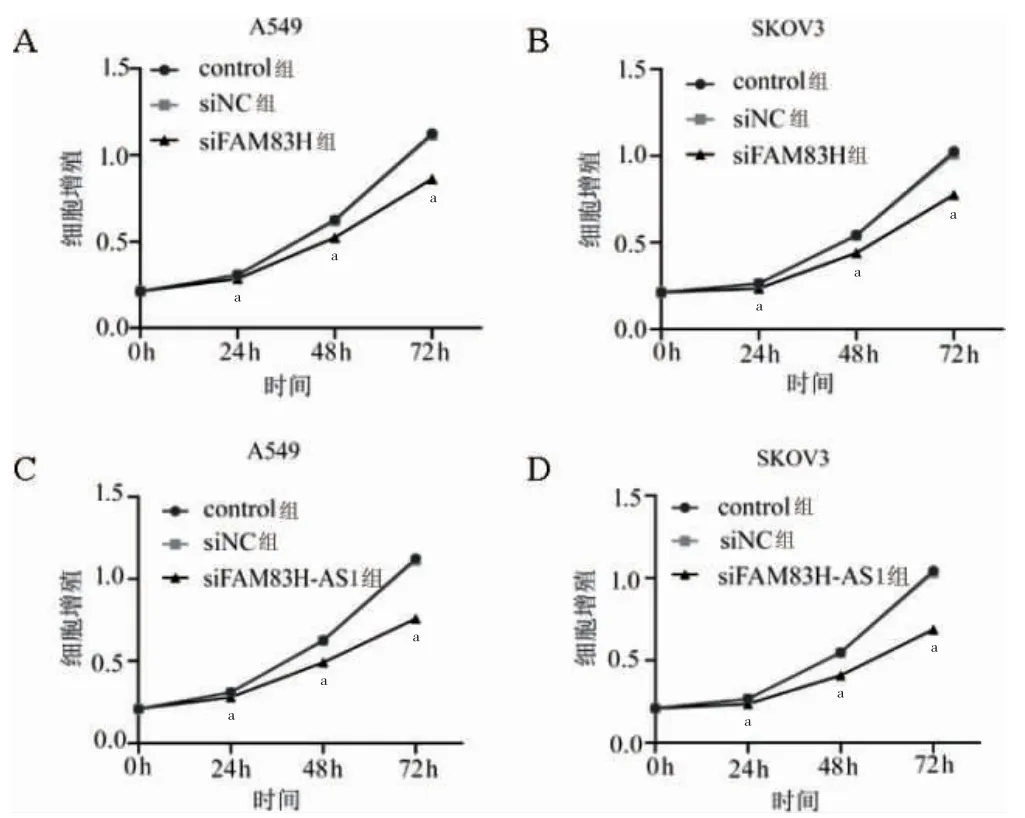
圖4 FAM83H、FAM83H-AS1 敲低后細胞增殖能力比較(n=3)
2.5 FAM83H、FAM83H-AS1 敲低后各組細胞凋亡數量比較
A549 和SKOV3 細胞中,siFAM83H 組、siFAM83HAS1 組凋亡數量高于siNC 組(P<0.05)。見圖5。
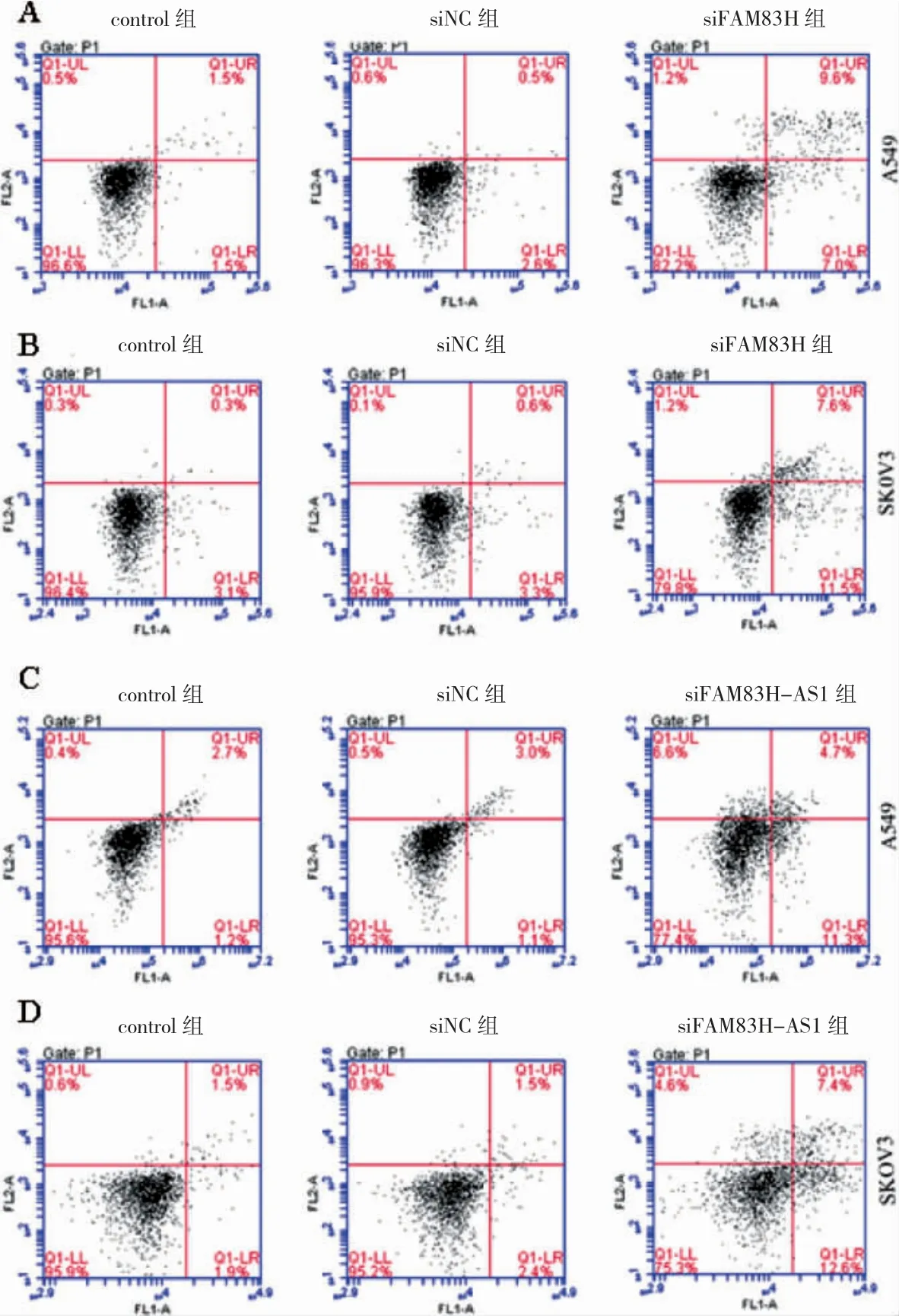
圖5 FAM83H、FAM83H-AS1 敲低后各組流式結果
2.6 FAM83H、FAM83H-AS1 敲低后各組細胞遷移數比較
A549 和SKOV3 細胞中,siFAM83H 組、siFAM83HAS1 組細胞遷移數低于siNC 組(P<0.05)。見圖6。
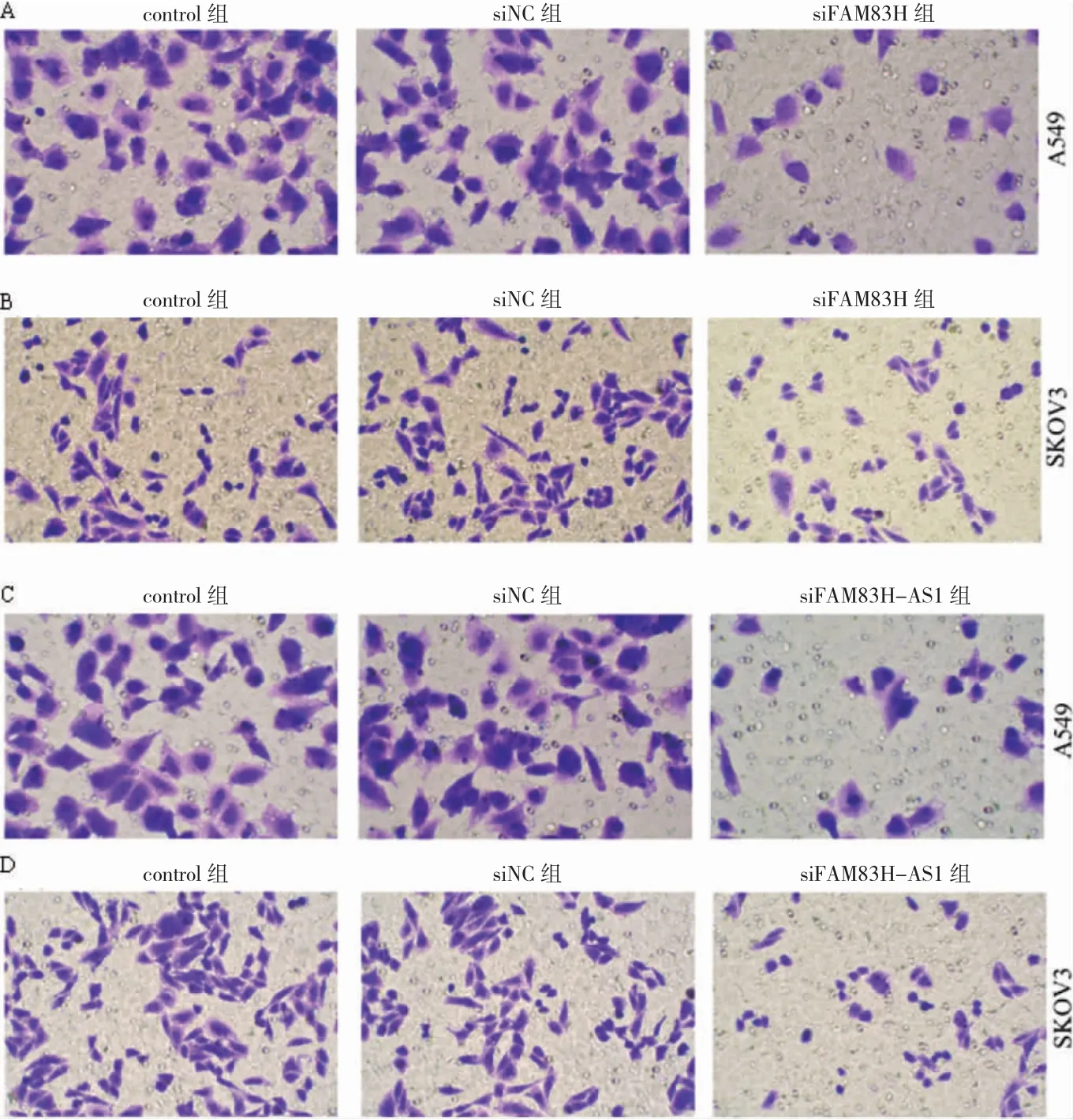
圖6 FAM83H、FAM83H-AS1 敲低后各組Transwll 檢測結果(結晶紫染色,200×)
2.7 FAM83H、FAM83H-AS1 表達的關聯性
FAM83H 與FAM83H-AS1 以頭對頭(5’-5’)形式互補重疊,推測二者存在相互調控關系。A549 和SKOV3細胞中,FAM83H 后敲低,siFAM83H 組 與siNC 組siFAM83H-AS1 mRNA 比較,差異無統計學意義(P>0.05);FAM83H-AS1 敲低后,siFAM83H-AS1 組 與siNC 組siFAM83HmRNA 和蛋白比較,差異無統計學意義(P>0.05)。見圖7。尚不能認為FAM83H 與FAM83HAS1 之間存在調控關系。
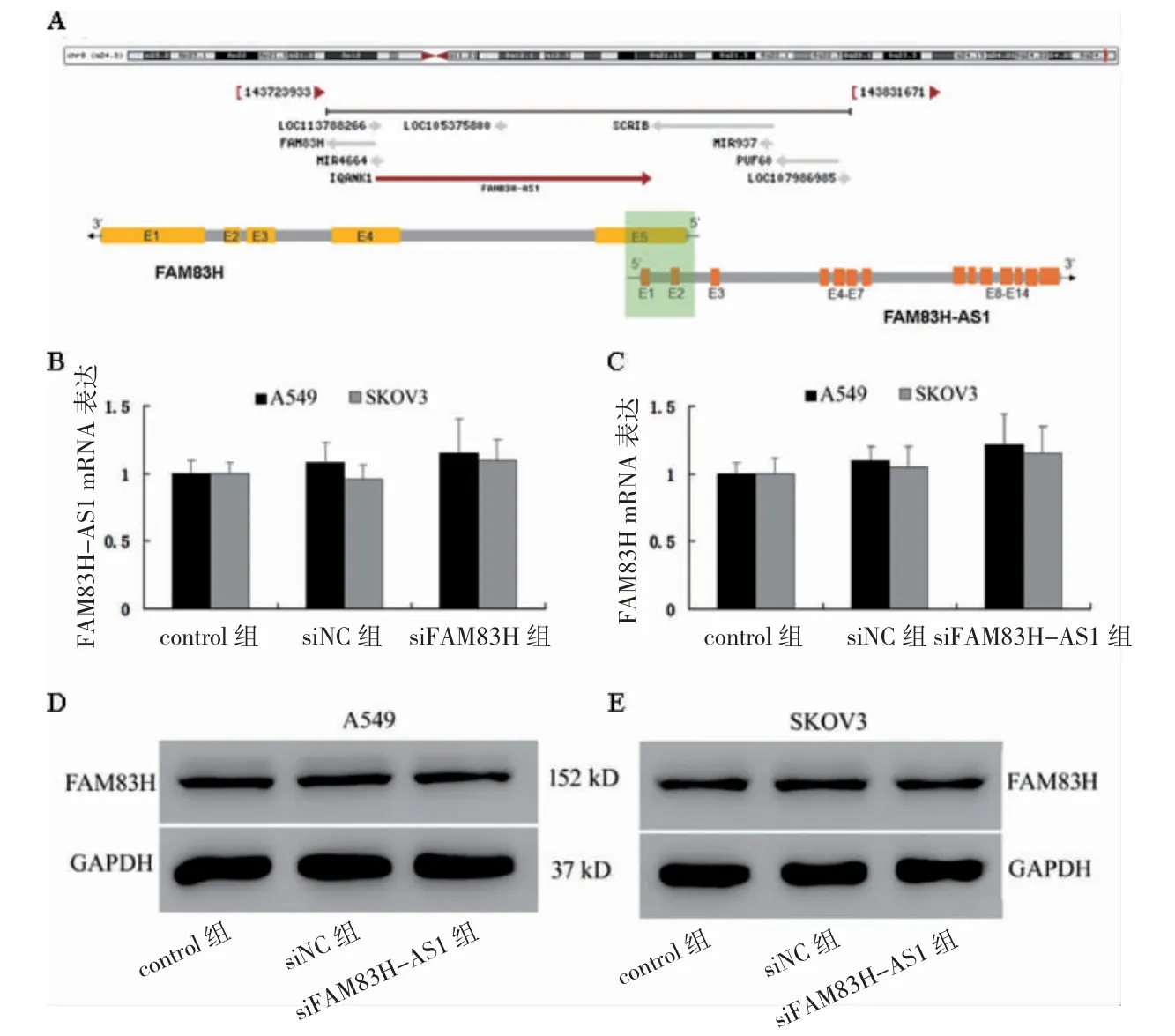
圖7 FAM83H、FAM83H-AS1 表達的關聯性(n=3)
3 討論
本研究結果顯示,FAM83H 和FAM83H-AS1 在多種腫瘤中上調,敲低FAM83H 或FAM83H-AS1 后,可抑制A549、SKOV3 細胞增殖和遷移,誘導細胞凋亡。提示二者在LUAD 和卵巢癌中具有癌基因的作用。
既往研究證實,FAM83H 基因在人類多種腫瘤中表達上調,且與不良預后相關,其機制涉及不同信號通路[11-13]。肝細胞癌中FAM83H 誘導細胞周期蛋白和基質金屬蛋白酶-2 表達,癌基因MYC 結合到FAM83H 啟動子區,促進其轉錄[2];胃癌中FAM83H 與Scrib 表達密切相關,FAM83H 和Scrib 可能通過穩定β-catenin 參與胃癌的進展[4];胰腺癌中FAM83H 過表達與CD8+T 細胞浸潤減少及Ras-PI3K-mTOR 信號通路有關[5];FAM83H 在非小細胞肺癌組織中的表達增加,與患者的臨床分期密切相關[14]。但FAM83H在卵巢癌中的表達尚未見報道,而本研究首次明確FAM83H 在LUAD 和卵巢中的生物學功能。
FAM83H-AS1 作為FAM83H 的天然反義轉錄本,其表達升高促進腫瘤發生發展[15-17]。FAM83H-AS1是LUAD 組織明顯上調的長鏈非編碼RNA 之一,FAM83H-AS1 通過調控Met/EGFR 信號,促進肺癌細胞增殖、侵襲和轉移[18];FAM83H-AS1 還可以結合hnRNP K 以增強抗凋亡基因RAB8B 和RAB14 的翻譯,從而抑制LUAD 凋亡[19]。FAM83H-AS1 在卵巢癌組織中高表達,其表達水平與巨噬細胞、中性粒細胞、樹突狀細胞等免疫細胞浸潤程度相關,下調其表達可明顯抑制卵巢癌細胞的增殖和侵襲[20-21];此外,FAM83HAS1 通過穩定HUR 蛋白參與卵巢癌的輻射抵抗和細胞轉移[22]。本研究進一步證實FAM83H-AS1 在LUAD和卵巢癌中的生物學功能。
有研究報道,內源性反義長鏈非編碼RNA 參與人類細胞基因表達的表觀遺傳調控[23],其表達水平與宿主基因的表達水平呈正相關或負相關,提示編碼和非編碼轉錄本在特定的生物學途徑中存在協同調節[24]。在食管癌中,FAM83H-AS1 與其正義鏈FAM83H在轉錄和翻譯水平上存在一致性調節,FAM83H-AS1可能通過調控FAM83H 的表達而發揮癌基因作用[25]。盡管FAM83H 與FAM83H-AS1 在泛癌種水平的表達趨向一致,但是本研究未在細胞水平證實二者對彼此表達的影響,FAM83H 與FAM83H-AS1 可能獨立發揮作用,具體機制仍待進一步研究。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
