
3,3′-二吲哚甲烷對腸炎沙門氏菌生物被膜的抑制作用
2022-02-23 11:33:14郭曉梅張森林
中國測試 2022年1期
張 慧,郭曉梅,白 婧,張森林,孫 群
(1. 四川大學輕工科學與工程學院,四川 成都 610064;2. 四川大學生命科學學院 生物資源與生態環境教育部重點實驗室,四川 成都 610064)
0 引 言
沙門氏菌(Salmonella)是引起微生物性食源性疾病最常見的致病菌之一[1]。每年全球范圍內大約94 000 000人因感染沙門氏菌而引起腸炎,其中約155 000人死亡[2]。2008年到2015年,我國食品中毒事件中,因微生物導致的食物中毒事件占60%以上,而在所有致病菌中,沙門氏菌占比為23%,居于首位[3]。自然界中,幾乎所有細菌都是以生物被膜的形式存在,只有約0.1%的細菌處于浮游狀態[4]。研究表明,生物被膜是造成食品污染、導致微生物性食源性疾病爆發的重要原因。據估計,約65%的食源性微生物污染由生物被膜狀態的致病菌引起[5]。成熟生物被膜胞外基質的阻隔性以及休眠狀態的里層細菌導致其對各種極端環境及壓力的抗性相對于浮游菌進一步增強[6],從而使得常規的消毒滅菌處理效果不佳,而且消毒殺菌劑的大量使用使得細菌產生抗性[7]。有研究表明次氯酸鈉濃度提高5倍也無法完全殺滅生物被膜里層腸炎沙門氏菌[8]。由此可見,生物被膜一旦成熟,難以完全清除,其導致的一系列問題如耐藥性的出現或增強,將會給食品安全造成巨大的潛在威脅。因此,預防生物被膜形成對于保證食品安全是十分迫切且重要的。
生物被膜形成大致分為初始粘附、細菌聚集、成熟和分散4個階段,且在不同階段,細菌所表現的生理特征各不相同[9],初始粘附階段大多數細菌能夠通過菌毛介導的游動或細菌自身的滑動性進行獨立運動來促進細菌粘附[10],而在細菌聚集和成熟階段則主要是通過產生胞外多糖(extracellular polysaccharide, EPS)、胞外 DNA (extracellular DNA,eDNA)、蛋白質和磷脂等物質組成胞外基質(extracellular matrix, ECM)形成并維持生物被膜三維結構[11]。3,3′-二吲哚甲烷 (3,3′-diindolemethane,DIM)是存在于十字花科蕓苔屬蔬菜中的一種植物化學物,此前研究表明DIM具有抗腫瘤、抗癌等多種生物活性[12],但是關于DIM對細菌生物被膜影響的研究較少且作用機制尚不完全清楚,目前缺乏定量評估DIM影響生物被膜形成過程、胞外基質以及微觀結構等的報道。
本研究通過測定生長曲線和細菌代謝活性分析DIM對S. enteritidis生長的影響,利用光學顯微鏡、激光共聚焦顯微鏡(CLSM)和掃描電鏡(SEM)觀察和分析DIM對生物被膜三維結構的影響,探究DIM對生物被膜形成過程、細菌運動性以及與生物被膜三維結構密切相關的胞外基質(EPS、eDNA)的影響,為DIM在食品生產中的應用提供理論基礎。
1 材料與方法
1.1 菌株與試劑
實驗所使用的Salmonella enteritidis為實驗室保存的分離株。
DIM (純度≥98%,CAS:1968-05-4,Aladdin 公司);TSB培養基(青島海博公司);LB培養基(青島海博公司);0.1%結晶紫染色液 (Solarbio公司);PBS片 劑 (Solarbio公 司);Styo 9試劑 (Thermo Fisher公司);2.5%戊二醛固定液(北京雷根公司);Gel extraction kit (Omega 公司);瓊脂粉、乙醇、甲醇、二甲基亞砜(DMSO)、蒽酮等試劑均購于成都鵬世達實驗用品有限公司。
1.2 主要儀器設備
Bio-Rad 680 酶標儀 (Bio-Rad 公司);BX53F 光學顯微鏡(Olympus公司);JSM-7500F掃描電子顯微鏡 (JEOL公司);TCS SP5II激光共聚焦顯微鏡(Leica公司);BD1110超微紫外分光光度計(Bio Drop 公司);Eppendorf 5804R 高速離心機 (Eppendorf公司)等。
1.3 方法
1.3.1 菌株活化
將保存在–20 ℃的S. enteritidis劃線于LB固體培養基活化,挑取單菌落接種于TSB液體培養基中,于 130 r/min、37 ℃ 搖床中培養 12 h 進行二次活化,將二次活化后的S. enteritidis劃線于LB固體培養基,培養18 h后將長有單菌落的固體平板保存于4 ℃。挑取單菌落接種于10 mL TSB液體培養基中,過夜培養活化。吸取1 mL過夜培養液于9 mL TSB液體培養基中,培養至菌液濃度約為1×108CFU/mL,備用。
1.3.2 最小生物被膜抑制濃度 (MBIC)測定
將DIM溶解于DMSO制備濃度為2×10–2mol/L母液。按照方法1.3.1準備菌液。在96孔板的中間選取 7列 8個復孔,第1列每孔加入5 μL DMSO、175 μL 1/10-TSB 培養基 (稀釋 10倍的 TSB 培養基)、20 μL TSB 培養基作為陰性對照;第 2 列每孔加入 5 μL DMSO、175 μL 1/10-TSB 培養基、20 μL按照方法1.3.1準備的菌液作為空白對照;第3列到第7列分別加入5 μL梯度稀釋的DIM溶液、175 μL 1/10 - TSB 培養基、20 μL 菌液,使最后 DIM 濃度分別為 7.8,15.6,31.2,62.5,100 μmol/L。將 96 孔板于28 ℃恒溫培養箱中培養48 h形成生物被膜。取出培養后的96孔板進行結晶紫染色定量生物被膜,具體操作步驟:1) 棄去培養基,用無菌PBS緩沖液輕輕洗滌兩次除去浮游細菌。每孔加入200 μL 95%甲醇溶液,室溫固定15 min后棄去固定液,室溫晾干。2) 每孔加入 200 μL 0.1% 結晶紫室溫染色15 min后棄去染液,用超純水洗滌至洗出液無色,室溫晾干。3) 每孔加入 200 μL 95% 乙醇溶液溶解15 min后,用酶標儀測定OD570。最小生物被膜抑制濃度(MBIC)定義為當處理濃度提高之后,生物被膜抑制率沒有顯著變化時的最小濃度[13],DIM對S. enteritidis生物被膜抑制率采用以下算式計算:
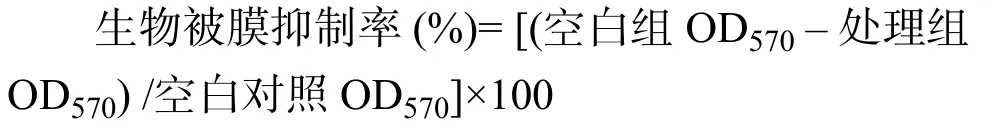
1.3.3 DIM 對S. enteritidis生長的影響
生長曲線測定:如方法1.3.1準備菌液,以1%的接種量接入 DIM濃度為 15.6,31.2 μmol/L的1/10-TSB培養基(稀釋10倍的TSB)中,以DMSO為空白對照。每2 h取一次樣直至48 h結束,測定OD600,繪制生長曲線。
細菌代謝活性測定:XTT作為線粒體脫氫酶的底物,被活細胞還原成水溶性的橙黃色甲臢產物。當XTT與電子偶合劑(例如PMS)聯合應用時,其所產生的水溶性的甲臢產物的吸光度與細胞代謝活性成正比,將XTT鈉鹽和吩嗪甲酯硫酸鹽(PMS)分別以 20 μg/mL和 300 μg/mL的濃度溶解在 PBS溶液中,并按1∶1比例混合,現配現用。同時,按照方法1.3.2培養生物被膜。培養結束后,收集每孔的浮游菌,并用無菌PBS洗滌浮游菌,以850×g反復離心 5 min 后向沉淀中加入 200 μL XTT 與 PMS的混合液,并迅速轉移至96孔板中;同樣,孔板底部的生物被膜用無菌PBS溶液洗滌3次后,向孔中加入 200 μL XTT與 PMS混合液。最后,將 96孔板在 37 ℃、130 r/min 條件下避光孵育 3 h,用酶標儀檢測OD490。
1.3.4 顯微鏡觀察生物被膜結構
普通光學顯微鏡觀察步驟:預先在24孔板每孔中從三個角度豎直放入3片無菌玻片(φ 14 mm),使其形成穩定三角結構,作為S. enteritidis生物被膜的載體。如方法1.3.1準備菌液。在24孔板中間選取4列4個復孔,第1列每孔加入25 μL DMSO、875 μL 1/10-TSB 培養基、100 μL TSB 培養基為陰性對照;第 2 列每孔加入 25 μL DMSO、875 μL 1/10-TSB培養基、100 μL菌液為空白對照;第3列和第4 列每孔加入 25 μL DIM 溶液、875 μL 1/10-TSB 培養基、100 μL菌液,使孔中DIM終濃度分別為15.6,31.2 μmol/L,隨后將孔板置于 28 ℃ 恒溫培養箱中培養48 h。生物被膜培養結束后,取出玻片按方法1.3.2進行結晶紫染色并用光學顯微鏡觀察并拍攝放大400倍下的圖像。
激光共聚焦顯微鏡觀察步驟:如上述方法培養生物被膜結束后,取出覆有生物被膜的玻片,并用0.85% 氯化鈉溶液清洗 3 次,取 100 μL 3.34 μmol/L的Syto 9染液均勻覆蓋在玻片表面,將玻片置于28 ℃培養箱中避光染色15 min后。立即用激光共聚焦顯微鏡觀察生物被膜并采集圖像,并使用ImageJ軟件重建生物被膜三維結構,COMSTAT2.1 (Dr. Claus Sternberg, DTU Systems Biology, Technical University of Denmark, Denmark)軟件計算三維結構的定量參數。
掃描電子顯微鏡觀察步驟:預先在48孔板每孔中從三個角度豎直放入3片無菌玻片(φ 8 mm),使其形成穩定三角結構,作為S. enteritidis生物被膜的載體。按照方法1.3.2培養生物被膜。將覆有生物被膜的玻片用無菌PBS溶液清洗3次后立即用 2.5%戊二醛固定液于 4 ℃固定 3 h。用 30%、50%、60%、70%、90%、95%和100%的乙醇溶液梯度脫水10 min后進行臨界點干燥和噴金處理。掃描電鏡觀察并拍攝放大2000倍和5000倍下的圖像。
1.3.5 生物被膜形成過程的測定
將20 mL如方法1.3.1準備的菌液與 180 mL 1/10-TSB培養基混合均勻后,向96孔板每孔中加入 200 μL,分別在不同時間點 (4,8,12,18,24,34,48,58,72,82,96,120 h)取出樣品,每個時間點 18個復孔,按方法1.3.2進行結晶紫染色,酶標儀檢測OD570,繪制生物被膜形成過程曲線。如方法1.3.2培養S. enteritidis生物被膜,不同之處在于分別在培養的 0,6 ,12,24,36 ,38 h 加入 DIM 溶液使孔中終濃度為 31.2 μmol/L,并于 48 h 停止培養。然后進行結晶紫染色,酶標儀檢測OD570,并計算在不同時間加入DIM時的生物被膜抑制率以評估DIM的抑制的具體階段。
1.3.6S. enteritidis運動性能的檢測
按照方法1.3.1準備菌液。準備3個50 mL游動性檢測培養基(1%胰蛋白胨、0.25%氯化鈉、0.3%瓊脂)和3個50 mL群集性能檢測培養基(1%胰蛋白胨、0.25%氯化鈉、0.7%瓊脂),冷卻到45 ℃左右分別加入1.25 mL DMSO作為空白對照、1.25 mL DIM 溶液使平板中終濃度分別為 15.6 μmol/L和31.2 μmol/L,混合均勻后倒平板,冷卻凝固后于平板中央垂直加入15 μL菌液,水平放置于37 ℃恒溫培養箱,24 h后觀察并拍照。
1.3.7 胞外 DNA (eDNA)的測定
按方法1.3.4在DIM處理下培養S. enteritidis生物被膜,但不在24孔板中放置玻片。生物被膜培養結束后棄去浮游細胞并清洗生物被膜,向每孔加 5 μL EDTA (0.5 M),4 ℃ 條件下靜置 1 h 后,向每孔加入700 μL TEN緩沖液重新懸浮生物被膜細胞。于 4 ℃ 下 18000×g離心 5 min,將 100 μL 上清液轉移到含有300 μL TE緩沖液的EP管中。然后,將混合物和等體積的結合緩沖液(Gel extraction kit)加入吸附柱。室溫靜置2 min 后,于4 ℃ 下18000×g離心 60 s,然后用洗滌緩沖液清洗吸附柱,18000×g反復離心30 s。最后加入無菌水,離心收集eDNA。使用超微紫外分光光度計測定eDNA濃度。
1.3.8 胞外多糖 (EPS)的測定
S. enteritidis生物被膜中胞外多糖的提取方法:如方法1.3.1所述準備S. enteritidis菌液,在直徑為60 mm 無菌培養皿中依次加入125 μL DMSO、4375 μL BHIGSS培養基、500 μL菌液作為空白對照,處理組加入 125 μL DIM 溶液使終濃度分別為 15.6 μmol/L和31.2 μmol/L。加樣結束后將培養皿于在28 ℃條件下培養48 h以便形成生物被膜。棄去浮游細胞后,用無菌PBS洗3次后將生物被膜洗脫至2 mL PBS 中,洗脫液以 9500×g離心 10 min。收集上清液為待測樣液。胞外多糖含量采用蒽酮硫酸法測定,具體檢測步驟如下:1) 精密稱取0.100 g蒽酮試劑,溶解于100 mL 80%的濃硫酸溶液中,定容后輕輕混合均勻配制成蒽酮硫酸溶液,現配現用。2) 精密稱取0.100 g葡萄糖標準品置于100 mL容量瓶中,蒸餾水定容后,稀釋成不同濃度的葡萄糖溶液。3) 向試管中加入2 mL葡萄糖溶液,以蒸餾水為對照,每組實驗3個重復。迅速加入6 mL蒽酮硫酸溶液,振蕩充分混勻,置于95 ℃沸水中加熱15 min后,迅速放在冰水中冷卻15 min,測定在 625 nm處的吸光值,然后繪制標準曲線。4) 取2 mL待測溶液于試管中,同前一樣的步驟,記錄在625 nm處的吸光值,根據標準曲線,計算待測樣品糖含量。
1.3.9 數據處理與統計分析
采用 GraphPad Prism version 7.00 統計軟件進行單因素方差分析(One-way ANOVA)并做圖,以P<0.05代表差異顯著,數據用平均值±標準偏差表示。
2 結果與分析
2.1 DIM 對 S. enteritidis的 MBIC 結果
由圖1可知,DIM在 7.8~31.2 μmol/L范圍內隨著濃度的升高,對S. enteritidis生物被膜的抑制效果呈現濃度依賴性增強。在31.2 μmol/L時對生物被膜的抑制率約35%,當濃度再升高時對生物被膜抑制效果無顯著升高(P>0.05)。由此可知,DIM對S. enteritidis的 MBIC 為 31.2 μmol/L。
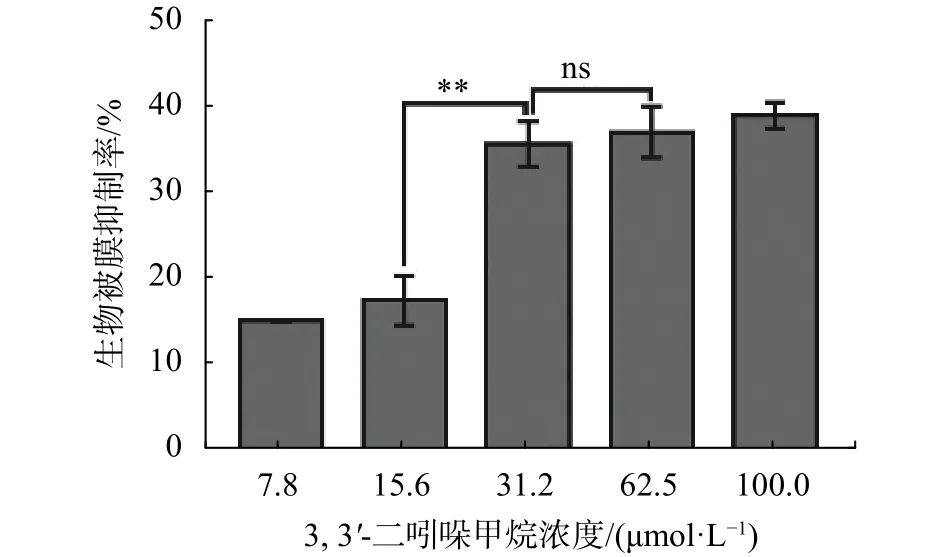
圖1 DIM 對 S. enteritidis 生物被膜的影響
2.2 DIM 對 S. enteritidis生長的影響
對細菌生長和生物被膜結構的影響是評估生物被膜抑制劑效果的兩個重要指標,有前景的生物被膜抑制劑應該是在抑制生物被膜的前提下,對細菌生長影響小或者幾乎沒有影響[14]。DIM對S. enteritidis生長的影響如圖2所示,結果顯示在培養48 h后,空白對照組和處理組的S. enteritidis細胞密度也沒有顯著差異 (P>0.05) (圖2(a)) 。且 DIM 濃度在15.6 ~31.2 μmol/L 范圍內,參與生物被膜形成的細菌代謝活性(被活細胞還原成的水溶性橙黃色甲臢產物在490 nm時的吸光值)以劑量依賴的方式減少,浮游細胞的代謝活性呈劑量依賴性增加,但對照組與處理組之間的代謝活性之和無顯著差異(P>0.05) (圖2(b)),證實 DIM 在測試濃度下對S.enteritidis無抗菌活性,即是指DIM的抗S. enteritidis生物被膜活性不是通過抑制生長實現的。
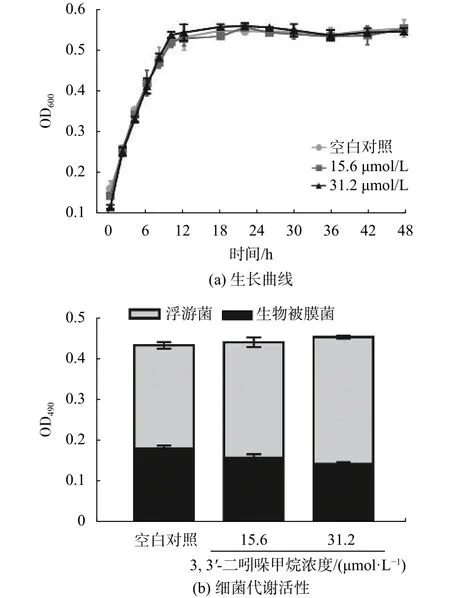
圖2 DIM 對 S. enteritidis 生長的影響(空白對照 :2.5% DMSO)
2.3 DIM 對 S. enteritidis生物被膜結構的影響
經過結晶紫和Syto 9熒光染料對DIM處理后的S. enteritidis生物被膜染色結果如圖3所示。普通光學顯微鏡圖像顯示對照組的生物被膜細胞團簇互相連接,而在實驗組中可明顯觀察到生物被膜細胞團簇變小且分散(圖3 (a))。在激光共聚焦顯微鏡圖像中也觀察到相似的結果,對照組中生物被膜的細胞團簇是互相連接的且熒光強度較強,在15.6~ 31.2 μmol/L范圍內隨著DIM濃度的增加,細胞團簇減少,熒光強度明顯減弱,在31.2 μmol/L處理組中只能觀察到分散的熒光微弱的細胞團簇(圖3 (b))。掃描電鏡圖像可見對照組的生物被膜細菌數量多且排列緊密,經過15.6 μmol/L的DIM處理后的S. enteritidis生物被膜細菌數量明顯減少,31.2 μmol/L的DIM處理的S. enteritidis生物被膜只分布了少數的單個菌落 (見圖4)。
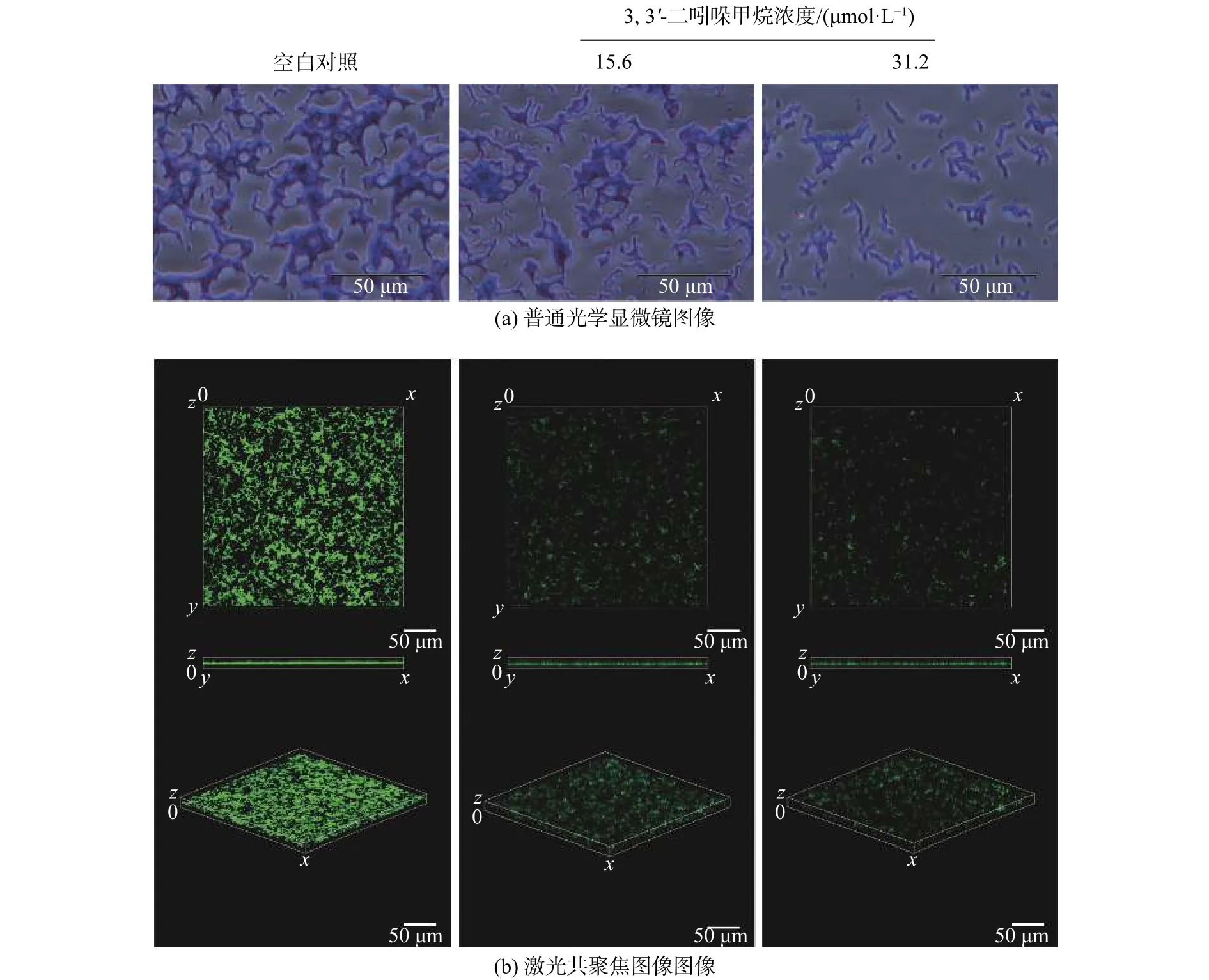
圖3 DIM 對 S. enteritidis 生物被膜結構的影響(空白對照 :2.5% DMSO)

圖4 掃描電子顯微鏡觀察 DIM 對 S. enteritidis生物被膜結構的影響(空白對照 :2.5% DMSO)
激光共聚焦三維重建圖像定量結果如表1所示,對照組中生物被膜厚度約為 25 μm,在 15.6~31.2 μmol/L內隨著DIM濃度的增加生物被膜的厚度呈現濃度依賴性變薄,經過31.2 μmol/L的DIM處理后的生物被膜的厚度僅8 μm,相比對照組顯著減少了約68% (P<0.05);對照組中生物被膜生物量約為9.615 μm3/μm2,經 15.6 μmol/L 和 31.2 μmol/L 的DIM 處理后分別為 2.828 μm3/μm2和 0.808 μm3/μm2,與對照組相比,處理組中單位面積生物量顯著減少了約70%和91% (P<0.05)。結果說明,DIM明顯抑制了生物被膜是三維結構形成且呈現濃度依賴性。
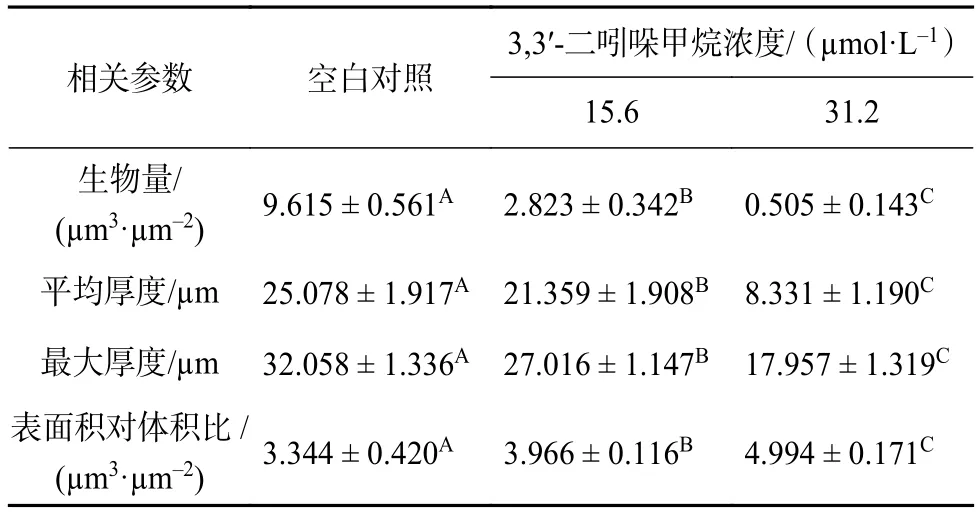
表1 COMSTAT2.1 分析 DIM 對 S. enteritidis生物被膜結構的影響1)
2.4 DIM 對 S. enteritidis生物被膜形成的影響
生物被膜形成大致分為初始粘附、細菌聚集、成熟和分散4個階段,且在不同階段,細菌所表現的生理特征各不相同[9],探究生物被膜形成過程有利于進一步研究DIM抑制生物被膜的機制。S.enteritidis生物被膜形成過程如圖5所示,在0~8 h內OD570值偏低且上升緩慢,此時并未形成穩定的生物被膜;8~34 h內OD570值迅速上升,說明此時孔板中粘附的菌體越來越多;34~72 h內的OD570值基本穩定,說明生物被膜已經成熟,達到一個穩定的狀態;72 h后OD570值不斷下降,說明粘附的菌體不斷脫落,生物被膜逐漸分散。由OD570變化趨勢可知S. enteritidis生物被膜形成過程如下:0~8 h是初始粘附階段,8~34 h 是細菌聚集階段,34~72 h是生物被膜成熟階段,72~120 h是生物被膜分散階段。
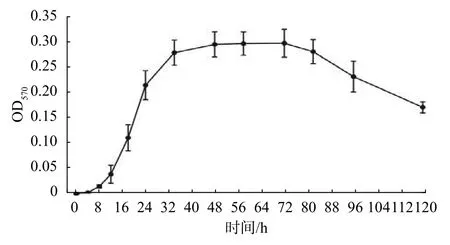
圖5 S. enteritidis 生物被膜形成過程
為探究DIM對S. enteritidis生物被膜形成過程的影響,實驗選擇在生物被膜形成的不同階段加入DIM。如圖6所示,0 h添加DIM時對生物被膜的抑制率約為40%,6 h之后添加DIM,生物被膜抑制率逐漸降低,24 h之后添加DIM,生物被膜抑制率僅為10%。綜上所述,DIM對S. enteritidis生物被膜的抑制作用主要體現在抑制生物被膜形成過程的初始粘附階段和細菌聚集階段。
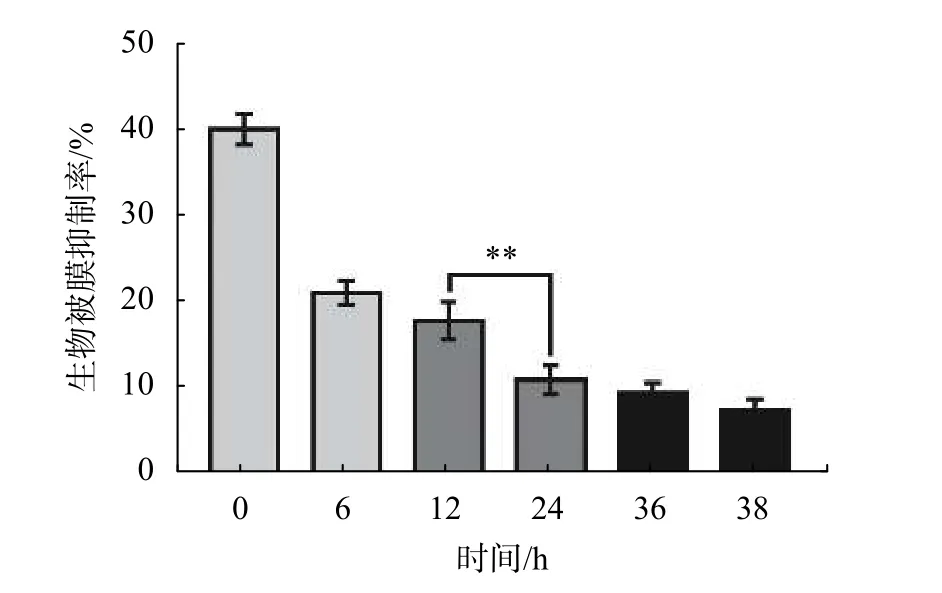
圖6 DIM 對 S. enteritidis 生物被膜的影響(**P < 0.01)
2.5 DIM 對 S. enteritidis運動性的影響
在粘附初始階段,運動性是微生物的一項重要生理活動,對細菌粘附表面和生物被膜的形成起著至關重要的作用[15]。由結果2.4可知DIM抑制了初始粘附階段,因此實驗探究了DIM對細菌運動性是否產生影響。S. enteritidis是一種有鞭毛的細菌,其運動性包含游動和群集/蜂擁兩種,研究表明在不同瓊脂含量的培養基中,細菌表現出的運動性也不同,瓊脂含量為0.3%時,細菌的運動主要表現菌體游動,當瓊脂含量為0.7%時,培養基中間隙較小,主要表現為群集運動[16]。DIM對S. enteritidis運動性的影響如圖7所示,與對照組相比,在15.6 μmol/L至31.2 μmol/L范圍內隨著DIM濃度增加,S. enteritidis游動的直徑逐漸減小,群集性能也逐漸減弱。
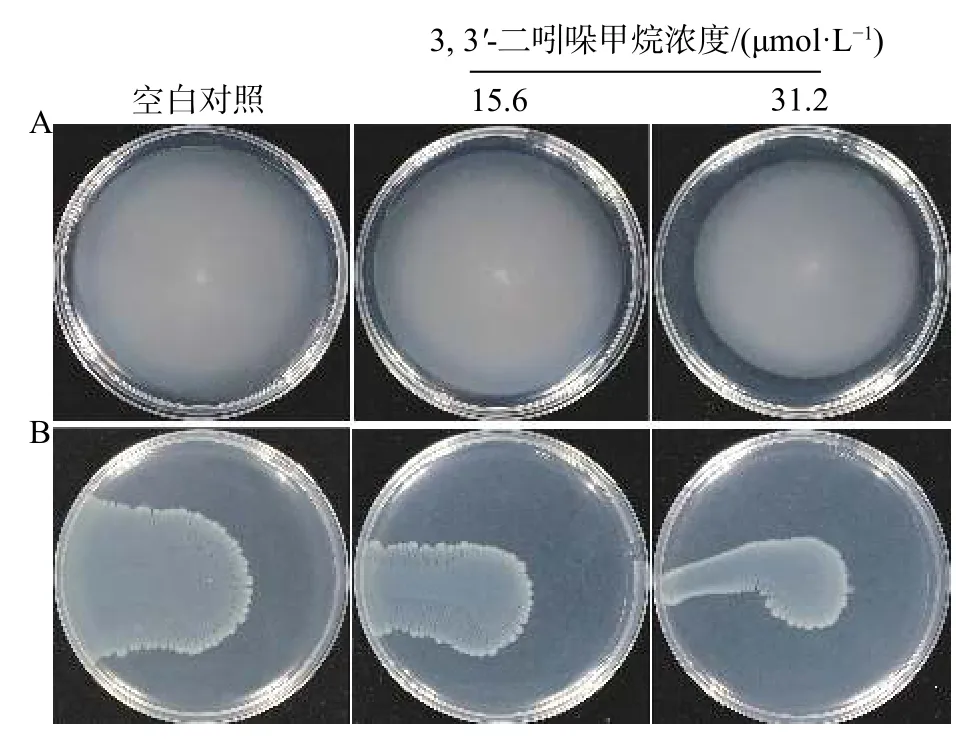
圖7 DIM 對 S. enteritidis 游動 (A)和群集 (B)的影響
2.6 DIM 對 S. enteritidis eDNA 的影響
eDNA是一種普遍存在的胞外基質,研究表明,eDNA存在于細菌細胞表面,通過酸堿相互作用增強了粘附和表面聚集[17]。DIM對S. enteritidiseDNA釋放的影響如圖8所示,DIM顯著抑制了S. enteritidiseDNA 的釋放 (P<0.05),在 15.6 ~31.2 μmol/L 范圍內隨著DIM濃度增加,胞外基質中eDNA濃度從1.6 μg/mL 降低至 1.0 μg/mL,與對照組相比分別降低約46%和67%。
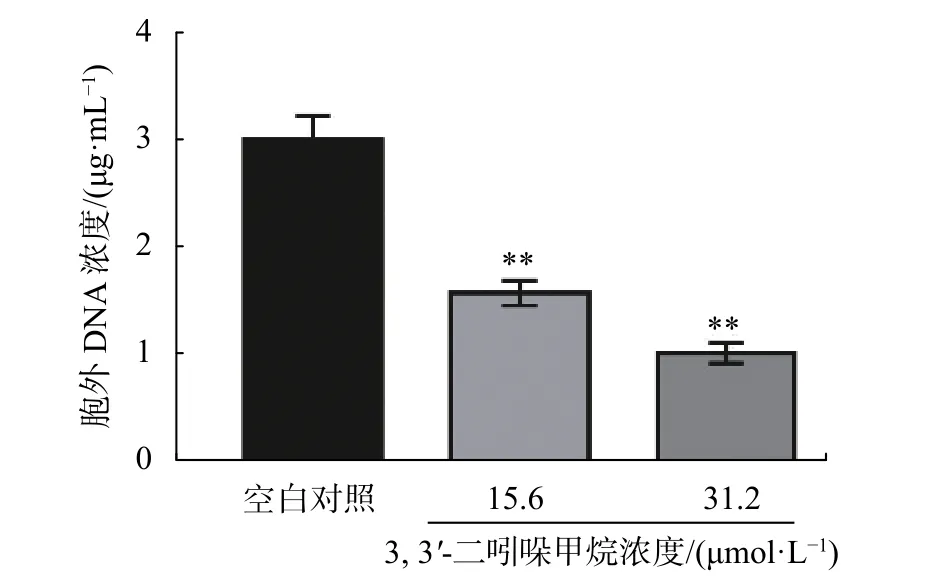
圖8 DIM 對 S. enteritidis eDNA 釋放的影響
2.7 DIM 對 S. enteritidis胞外多糖的影響
胞外多糖是保證生物膜三維結構完整性的重要因素,主要在生物被膜形成過程的細菌增殖階段產生[18]。相關報道表明,有效的生物被膜抑制劑能抑制沙門氏菌胞外多糖的產生[19]。由圖9可知,DIM顯著抑制了S. enteritidis胞外多糖的產生(P<0.05),DIM 在 15.6 μmol/L 至 31.2 μmol/L 范圍內,隨著濃度的增加,胞外多糖濃度逐漸降低,在15.6 μmol/L處理下,胞外多糖含量為5.8 μg/mL,相對于對照組降低約48%,在31.2 μmol/L處理下,胞外多糖濃度降低至3.4 μg/mL,相對于對照組降低約70%。
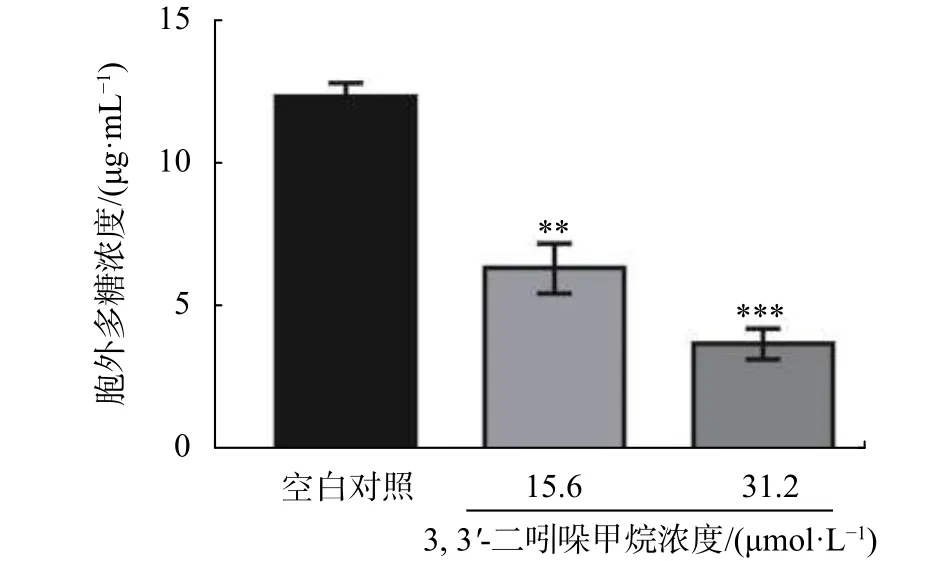
圖9 DIM 對 S. enteritidis 胞外多糖的影響
3 結束語
沙門氏菌是導致食源性疾病的重要致病菌之一,能通過在食品接觸表面甚至食品表面形成生物被膜來避免被清除[20]。生物被膜一旦成熟,其由EPS、eDNA、蛋白質等組成的ECM的阻隔性使消毒劑濃度在生物被膜中由外到內呈現梯度下降[6],導致不能完全消滅里層細菌,造成食品污染。因此,預防生物被膜形成對于保證食品安全尤為重要。
本研究表明,DIM使常見食源性致病菌S.enteritidis生物被膜厚度減少三分之二,有效抑制生物被膜形成三維結構。大多數細菌生物被膜的形成大致包含4個不同階段(初始粘附、細菌聚集、成熟、分散),細菌在不同階段表現出不同的生理活動。通過探究DIM具體作用階段以及對相關生理活動的影響,發現DIM主要抑制生物被膜形成過程中的細菌粘附及聚集,細菌運動和胞外基質分泌分別在初始粘附和細菌聚集階段起著關鍵作用[18]。本研究發現,DIM阻礙了S. enteritidis的游動和群集運動,可能通過抑制細菌運動性以及eDNA、胞外多糖的產生從而抑制S. enteritidis細菌粘附和聚集,導致生物被膜形成失敗。且DIM的抗生物被膜活性不是通過抑制細菌生長實現的,因此具有不易誘導細菌產生抗性的優勢,有良好的應用前景。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
科學大眾(2021年9期)2021-07-16 07:02:54
當代陜西(2021年2期)2021-03-29 07:41:24
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
