云頂山亞高山草甸土壤可培養細菌數量特征
2022-02-24 00:05:51張寶貴朱茜劉曉嬌吳佳靈常麗娟
種子科技 2022年1期
張寶貴 朱茜 劉曉嬌 吳佳靈 常麗娟
摘 ? ?要:作為土壤生物的主要組成部分,土壤微生物在整個陸地生態系統物質循環和能量轉換過程中發揮著非常重要的作用,且對環境變化極其敏感。以呂梁山中麓云頂山亞高山草甸為研究區域,以不同海拔(2 450 m、2 500 m和2 550 m)、不同深度(0~10 cm和20~30 cm)土壤為研究對象,采用可培養方法測定了土壤細菌的數量,同時測定了土壤理化性質并進行了相關分析。結果表明,研究區不同海拔、不同深度可培養細菌數量介于3.13×109 ~10.4×109 CFU/g,且同一海拔表層土壤細菌數量均顯著高于30 cm深處(P<0.05),不同海拔0~10 cm與20~30 cm深度土壤細菌數量均隨著海拔高度的升高而顯著下降(P<0.05)。細菌數量與土壤全碳(STC)和土壤溫度(ST)呈極顯著正相關(P<0.01),與土壤酸堿度(pH值)呈顯著正相關(P<0.05),表明土壤全碳、土壤溫度及土壤pH值是影響該區域土壤細菌數量的主要環境因子。研究結果可為區域應對氣候變暖提供理論依據與數據支撐。
關鍵詞:云頂山;亞高山草甸;可培養細菌;數量特征
文章編號:1005-2690(2022)01-0017-05 ? ? ? 中國圖書分類號:S812.2 ? ? ? 文獻標志碼:B
土壤微生物是地球上多樣性最高、物種最豐富的生物類群之一,參與了土壤中幾乎所有的物質轉化過程,對森林、草原、濕地及農田等生態系統具有重要影響,是驅動地球生物化學過程的關鍵因素[1]。作為土壤圈層能量交換的中介,土壤微生物可以直接分解土壤中的有機質,促進腐殖質形成[2],提供植被生長所需的營養[3],還能間接反映土壤的肥力狀況[4]。雖然土壤微生物生物量只占土壤有機質的3%左右,卻是植物養分轉化和循環的驅動力,在土壤肥力和生態系統評價中起著非常重要的作用[5]。
在當前氣候變化具有區域差異性的背景下,人類活動對土壤生態系統的干擾不斷加強,使得我國土壤微生物群落結構更為復雜[6]。作為響應氣候變暖的產物,亞高山草甸目前已經引起諸多國內外學者的重視。學者主要針對亞高山草甸植物物種的高氮保留能力[7]、物種擴張和土壤養分的影響[8]開展了相關研究,而對地下微生物研究鮮有報道。王萍(2011)[9]研究發現,亞高山草甸土壤微生物多樣性在棄耕地恢復中能夠得到較好恢復;宋三多(2011)[10]研究發現,在不同雪被厚度覆蓋下,亞高山草甸土壤細菌種群結構無明顯變化,但對細菌的分布比例影響較大;謝勇等(2017)[11]研究發現,封育后亞高山草甸土壤細菌數量和土壤微生物總數顯著降低;朱雪梅(2009)[12]研究發現若爾蓋亞高山草甸不同土壤層次的細菌數量有明顯差異,隨著剖面層次增加,細菌數量逐漸變小。研究亞高山草甸土壤微生物有助于理清高海拔地區土壤微生物對環境變化的響應特征。
云頂山位于山西省呂梁山中部,生態環境復雜,具有大面積的亞高山草甸,屬于典型的中緯度、低海拔亞高山草甸[13]。作為全球氣候變化的敏感區域,山西省亞高山草甸成為諸多學者關注研究的熱點。近幾年對山西境內土壤微生物研究多是結合典型植被類型進行宏觀方面的研究[14]。相關學者主要對山西境內亞高山草甸植被群落、物種多樣性等地上生物進行了時空變化格局的研究[15-17]。上官鐵梁、張峰、劉明光、劉瑩、李素清、張金屯、楊斌勝等則針對云頂山這一地區的特定植物群落進行了集中研究[18-22]。
以上研究多集中在環境類型和人為干擾對植被群落多樣性產生的影響上,沒有對該區域土壤微生物進行研究。基于此,本研究以山西省境內云頂山不同海拔、不同深度亞高山草甸土壤為研究對象,通過可培養技術研究云頂山亞高山草甸土壤細菌數量特征,同時分析細菌數量與環境因子之間的相關性,找出限制該區域細菌數量的關鍵環境因子。研究結果可為系統認識區域內亞高山草甸土壤微生物響應氣候變化提供理論依據與數據支撐。
1 研究地區與研究方法
1.1 研究區概況
云頂山地處呂梁山中麓,位于山西省呂梁市交城縣、方山縣和太原市婁煩縣的交界處(111°30'~111°47'E,37°50'~38°02'N),最高海拔2 720 m,年平均氣溫為3~4 ℃,最熱月為7月,平均氣溫16.1 ℃,最冷月為1月,平均氣溫-10.6 ℃,年均降水量600~800 mm,降水集中于6—9月,相對濕度70.9%,無霜期100 d,屬于溫帶大陸性季風氣候;土壤類型從低海拔到高海拔依次為褐土、山地褐土、山地淋溶褐土、山地棕壤、亞高山草甸土;植被類型從山麓到山頂依次為1 200~1 750 m落葉闊葉林帶、1 750~2 200 m針闊葉混交林帶、2 200~2 600 m寒溫性針葉林帶、2 600~2 720 m亞高山灌叢草甸帶。
1.2 樣地設計與樣品采集
樣品采集于2019年9月,根據云頂山亞高山草甸分布的實際范圍,從底部到頂部按50 m高差確定3個采樣點,分別為海拔2 450 m(YDL)、2 500 m(YDM)、2 550 m(YDT)。在每個海拔高度處,選取地勢平坦的區域,設置3個間隔5 m的平行樣地,在每個樣地的中心挖取土壤剖面。
由于山地土層較薄,根據實際情況分別采集表層土(0~10 cm)和剖面底部(20~30 cm)土壤樣品,盛于微生物采樣袋中,隨后放置于儲有冰袋的低溫保存箱中并運回實驗室,保存于-20 ℃冰箱備用。同時采集用于土壤理化性質分析的樣品。為保證采樣的準確性,采樣過程嚴格按照無菌要求進行。采樣過程中使用一次性無菌PE手套,以免對土壤造成污染。采樣點信息見表1。
1.3 細菌的分離培養
細菌的分離培養采用稀釋涂布平板法培養,使用牛肉膏蛋白胨培養基,具體操作步驟如下。在超凈工作臺中稱取2 g新鮮土壤放入無菌三角瓶中,加入高溫滅菌后的18 mL無菌生理鹽水(0.85%),以室溫250 rmp搖振30 min,靜置數分鐘后取土壤懸液上清液200 μL加入800 μL無菌生理鹽水中并混合均勻,從此溶液中吸取200 μL土壤懸液加入到800 μL無菌生理鹽水中。
重復以上步驟將溶液稀釋至10-7數量級。為了確保微生物培養計數的準確性,可培養細菌進行預試驗,用各種稀釋濃度的土壤懸液進行涂布并培養,最終確定最適合細菌生長的濃度為10-6數量級。用移液槍吸取對應濃度的土壤懸液涂布到滅菌的固體培養基平板上,同一樣品同一類微生物進行3個重復處理,放置于25 ℃恒溫培養箱培養7 d后計數。
1.4 土壤理化性質的測定
土壤含水量(SWC)采用烘干法(105 ℃,24 h)測定;土壤pH值采用METTLER TOLEDO FE28-Standard酸度計測定;土壤電導率(EC)、土壤總溶解性固體(TDS)和土壤鹽度(SSC)采用電導率儀FE38 Standard測定;土壤全碳(STC)采用C/N分析儀2100(Elementar Vario-EL,Germany)測定;土壤溫度ST采用土壤溫度傳感器(S-
TMB-M006)測定。
1.5 數據處理與分析
采用Excel 2010處理所有數據;用SPSS 17.0進行相關統計分析,包括單因素方差分析和相關性分析;用Origin 8.0繪圖。
2 結果與分析
2.1 不同海拔、不同深度土壤理化性質變化特征
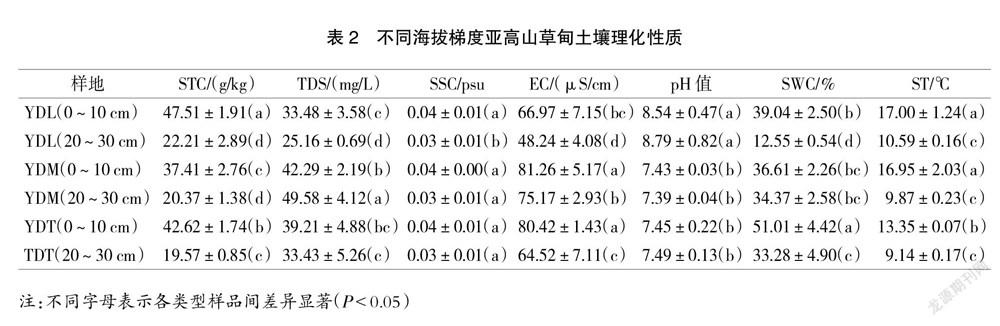
云頂山亞高山草甸不同海拔、不同深度下的土壤理化性質特征,詳見表2。從表2可知,STC介于19.57~47.51 g/kg之間。從橫向來看,同一海拔下隨著土壤深度增加,STC呈現顯著下降趨勢(P<0.05);從縱向來看,同一深度STC隨著海拔高度的增加而呈現先下降后上升的顯著趨勢(P<0.05)。
EC介于48.24~81.26 μS/cm之間。
SWC介于12.55%~51.01%之間,且變化趨勢與STC一致。
ST介于9.41~17.00 ℃之間,從橫向來看,同一海拔下ST變化范圍與STC一致;從縱向來看,同一深度下ST隨著海拔的升高呈現下降趨勢。
TDS介于25.16~49.58 mg/L之間,從橫向來看,隨著土壤深度的增加,不同海拔樣品變化趨勢不同,高海拔與低海拔地區呈現顯著下降趨勢(P<0.05),中海拔則呈現相反趨勢;從縱向來看,同一深度TDS變化趨勢與STC相同,但未達到顯著水平。
土壤pH值和SSC變化范圍較小。其中pH值介于7.39~8.79之間,呈弱堿性;SSC介于0.03~0.04 psu之間。兩者的變化趨勢均與STC一致,但未達到顯著水平。
2.2 不同海拔、不同深度土壤可培養細菌數量變化特征
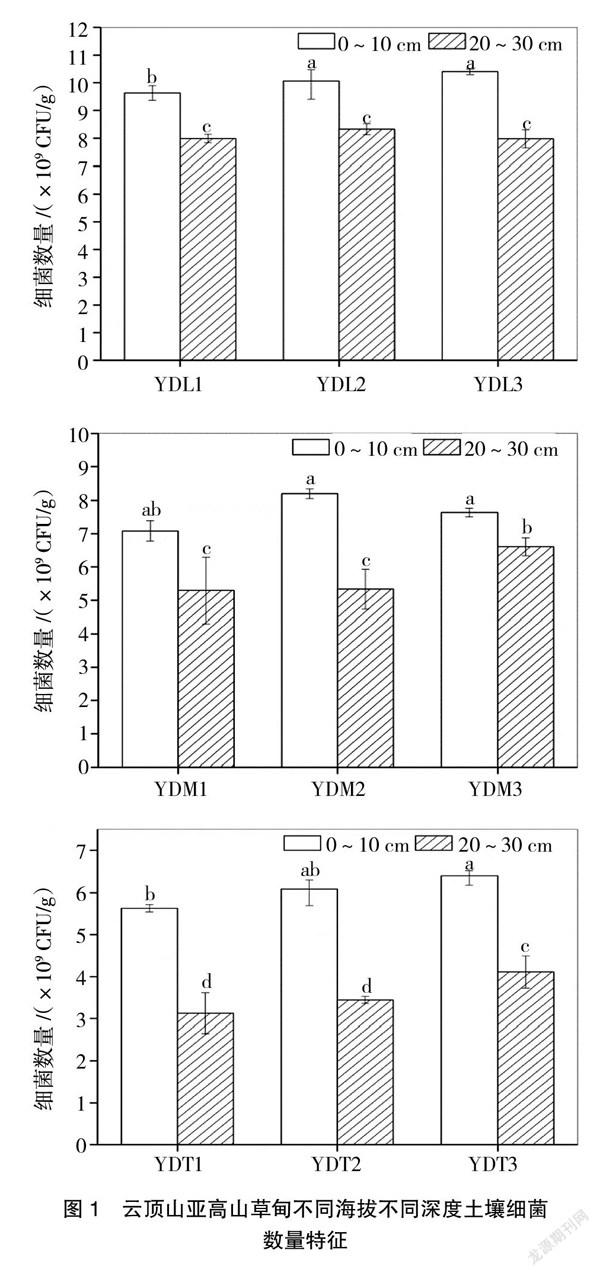
從圖1可以看出,研究區域可培養細菌數量介于3.13×109~10.4×109 CFU/g之間,YDL3樣點0~10 cm可培養細菌數量最高,為10.4×109 CFU/g,而YDT1樣點20~30 cm數量最少,為3.13×109 CFU/g,低了1個數量級。此外,從圖1可以看出,同一海拔所有3個重復均表現為0~10 cm細菌數量顯著高于20~30 cm細菌數量(P<0.05),說明土壤深度對細菌數量存在顯著影響。
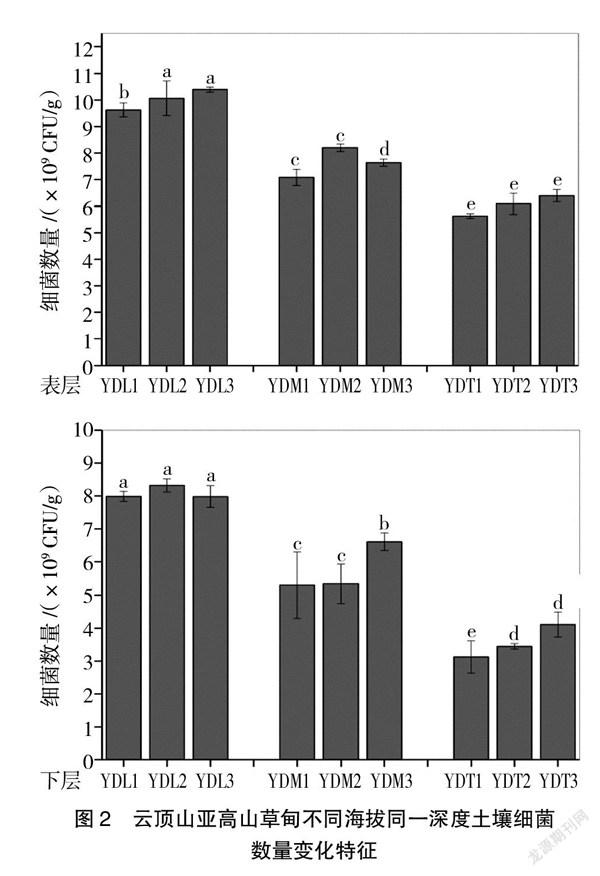
縱向來看,從圖2可以看出,海拔高度對細菌數量影響顯著。不同海拔高度0~10 cm與20~30 cm深度土壤細菌數量均表現為隨著海拔高度的升高而顯著下降(P<0.05),具體表現為YDL樣點土壤細菌數量顯著高于YDM與YDT樣點(P<0.05),且YDM與YDT樣點之間土壤細菌數量也存在顯著性差異。
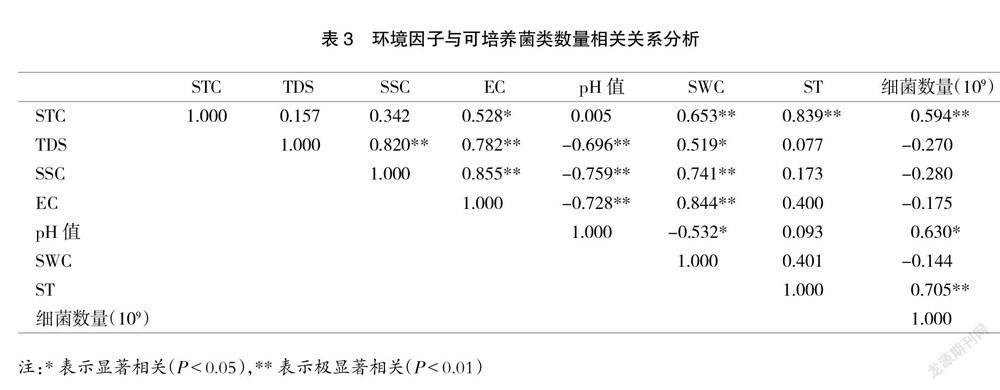
2.3 環境因子與土壤微生物數量的相關分析
采用SPSS 17.0進行環境因子與可培養細菌數量的相關性進行分析。從表3可知,可培養細菌數量與STC和ST均呈極顯著正相關關系(P<0.01),與pH值均呈顯著正相關關系(P<0.05)。
3 討論
細菌是土壤微生物中數量最多的一個類群,占土壤微生物總量的70%~90%,在土壤肥力形成過程中起著重要的作用[23]。研究結果表明,云頂山亞高山草甸土壤細菌數量介于3.13×109 ~10.4×109 CFU/g之間。而相關學者研究發現若爾蓋亞高山草甸土壤細菌的數量介于0.5×105~28.5×105 CFU/g之間,這可能是由不同地區環境異質性間接通過凍土溫度、植被覆蓋度、土壤理化性質等因素對土壤微生物生長繁殖產生影響。
土壤深度和海拔高度也對可培養細菌數量影響明顯。本研究表明,同一海拔0~10 cm和20~30 cm深度土壤可培養細菌數量隨著土壤深度的增加逐漸變小。這與李定瑤(2014)[24]和李越(2012)[25]等研究者的成果一致,其研究均發現可培養微生物的數量隨著土壤深度的增加呈現下降趨勢。表層土壤物理環境變化較為劇烈,底物輸入波動較大、植物根系與土壤微生物對養分的競爭較為激烈[26],適宜的水分、高含量的有機質和養分狀況,使得微生物所受脅迫小,有利于微生物群落的發展[27],而深層土壤水分長期處于飽和狀態且土壤溫度更低[28],制約了微生物群落的發展,造成不同深度土壤微生物數量變化的因素。
本研究還發現,不同海拔0~10 cm和20~30 cm深度可培養細菌數量均隨著海拔的升高呈現下降趨勢。這與毛文梁等(2013)[29]研究結果一致,其研究發現不同海拔下祁連山土壤中可培養細菌數量隨著海拔的升高而呈現降低的趨勢,且兩者之間呈現負相關。顯然,海拔高度能間接導致溫度、水分、光照、土壤微環境及植物群落等環境因子的變化,最終影響到土壤微生物的生長和活性[30]。
相關分析表明,限制研究區內亞高山草甸細菌數量的關鍵環境因子包括STC、ST與土壤pH值。類似的,相關學者發現北麓河北高寒草原區土壤全碳、含水率與細菌群落結構極顯著相關[31];甘南亞高寒草甸不同坡向土壤微生物群落分布的顯著差異是受土壤pH值、全磷和含水量等多種環境因子共同作用的結果[32];亞甲貢山土壤溫度和總氮是影響細菌生物量的主要因素[33]。也有學者發現,土壤溫度是對土壤微生物多樣性變化貢獻最大的因素[34],這可能是因為細菌適合生存在溫度適宜、營養豐富、有機物易于分解的土壤中,而隨著海拔不斷升高,溫度成為環境的主導性因素,因而使得細菌數量呈現隨海拔升高而遞減的趨勢。綜上所述,不同區域限制微生物數量的關鍵環境因子不盡相同,需要區別對待。
4 結論
云頂山亞高山草甸不同海拔、不同土壤深度細菌數量具有明顯差異,且與環境因子具有密切關系。本研究結論如下:研究區不同海拔、不同深度可培養細菌數量介于3.13×109~10.4×109 CFU/g之間;不同海拔、不同土壤深度下可培養細菌數量明顯不同,具體表現為同一海拔0~10 cm深度土壤細菌數量顯著高于20~30 cm深度(P<0.05),不同海拔0~10 cm與20~30 cm深度土壤細菌數量均隨著海拔高度的升高而顯著下降(P<0.05)。此外,本研究還發現,在環境因素對土壤細菌數量的影響方面,細菌數量受STC和ST極顯著影響,受土壤pH值顯著影響。
參考文獻:
[1]VERESOGLOU S D, HALLEY J M, RILLIG M C. Extinction risk of soil biota[J].Nature Communications,2015(6):8862-8871.
[2]許光輝,鄭洪元,張德生,等.長白山北坡自然保護區森林土壤微生物生態分布及其生化特性的研究[J].生態學報,1984,4(3):207-223.
[3]ROY S, SINGH J. Consequences of habitat heterogeneity foravailability of nuterients in a dry tropical forest[J].Journal of Ecology,1994,82(3):503-509.
[4]DILLY O, MUNCH J C. Ratios between estimates of microbialbiomass content and microbial activity in soils[J].Biology and Fertility of Soils,1998,27(4):374-379.
[5]宋長青,吳金水,陸雅海,等.中國土壤微生物學研究10年回顧[J].地球科學進展,2013,28(10):1087-1105.
[6]金凱.中國植被覆蓋時空變化及其與氣候和人類活動的關系[D].咸陽:西北農林科技大學,2019.
[7]André Pornon, Marion Boutin, Thierry Lamaze. Contribution of plant species to the high N retention capacity of a subalpine meadow undergoing elevated N deposition and warming[J].Environmental Pollution,2018,245(2):235-242.
[8]歐陽青,任健,尹俊,等.短期增溫對亞高山草甸土壤養分和脲酶的影響[J].草業科學,2018,35(12):2794-2800.
[9]王萍.亞高山草甸棄耕地恢復演替過程中土壤微生物多樣性研究[D].蘭州:蘭州大學,2011.
[10]宋三多.不同雪被厚度下亞高山草甸土細菌群落結構變化研究[D].雅安:四川農業大學,2011.
[11]謝勇,尹海燕,初曉輝,等.放牧和封育對滇西北亞高山草甸土壤養分及微生物特性的影響[J].云南農業大學學報(自然科學),2017,32(6):1121-1128.
[12]朱雪梅.若爾蓋高原亞高山草甸土壤細菌遺傳多樣性分析[D].雅安:四川農業大學,2009.
[13]李曉麗,徐滿厚,孟萬忠,等.模擬增溫對云頂山亞高山草甸水熱因子及群落結構的影響[J].生態學報,2020,40(19):6885-6896.
[14]吳然,康峰峰,韓海榮,等.山西太岳山不同林齡華北落葉松林土壤微生物特性[J].生態學雜志,2016,35(12):3183-3190.
[15]張世雄,楊曉艷,溫靜,徐滿厚.山西呂梁山亞高山草甸物種多樣性的時空變化格局[J].生態學報,2018,38(18):685-6693.
[16]馬麗,徐滿厚,周華坤,等.山西亞高山草甸植被生物量的地理空間分布[J].生態學雜志,2018,37(8):2244-2253.
[17]王譽陶,畢玉婷,王倩,等.山西亞高山草甸植物群落物種多樣性的空間分異[J].中國農學通報,2018,34(18):77-83.
[18]上官鐵梁,張峰.云頂山虎榛子灌叢群落學特性及生物量[J].山西大學學報(自然科學版),1989,12(3):347-352.
[19]上官鐵梁,張峰.云頂山植被及其垂直分布研究[J].山地研究,1991,9(1):26-29.
[20]劉明光,劉瑩,張峰,等.云頂山自然保護區植物群落的分類與排序[J].林業資源管理,2011(4):82-88.
[21]李素清,張金屯.山西云頂山亞高山草甸群落生態分析[J].地理研究,2007,26(1):83-90.
[22]李素清,楊斌勝,張金屯.山西云頂山亞高山草甸優勢種群和群落的格局分析[J].應用與環境生物學報,2007,13(1):9-13.
[23]Wang K Y, YangW Q, Hu T X. Processes of subalpine forest ecosystem s in the West of Sichuan[M].Chengdu: Sichuan Science& Technologv Press,2004.
[24]李定瑤.青藏高原多年凍土區可培養微生物多樣性及生長特性研究[D].蘭州:蘭州大學,2014.
[25]李越.土壤可培養微生物在黑土碳氮轉化中的作用研究[D].長春:吉林農業大學,2012.
[26]Zhu S Y, Zhou B, Mao Z J,et al. Space-time dynamics of fine root biomass of six forests in Maoershan forest region[J].Scientia Silvae Sinicae,2006,42(6):13-19.
[27]李香真,曲秋皓.蒙古高原韻原土境微生物量碩氧特征[J].土壤學報,2002,39(1):97-104.
[28]王長庭,龍瑞軍,王啟基.不同類型高寒草地群落物種特征和均勻度的重要性[J].草地學報,2005,13(4):320-323.
[29]毛文梁,臺喜生,伍修錕,等.黑河上游祁連山區土壤可培養細菌群落生境的垂直分異特征[J].冰川凍土,2013,35(2):447-456.
[30]都耀庭.高寒草甸土壤微生物數量沿海拔梯度的變異[J].黑龍江畜牧獸醫,2015(9):130-133.
[31]張曉蘭.熱融滑塌對青藏高原中部多年凍土區土壤細菌群落的影響[D].蘭州:蘭州交通大學,2019.
[32]李瑞,劉旻霞,張燦,等.甘南亞高寒草甸不同坡向土壤微生物群落分布特征[J].生態環境學報,2017,26(11):1884-1891.
[33]CUI H J, WANG G X, YANG Y, et al. Soil microbial community composition and its driving factors in alpine grasslands along a mountain elevational gradient[J].Journal of Mountain Science,2016,13(6):1013-1023.
[34]金裕華.武夷山不同海拔土壤微生物多樣性的變化特征[D].南京:南京林業大學,2012.
