中街山列島海洋牧場大黃魚時空分布與環境因子的關系
2022-02-24 14:22:02周永東蔣日進徐開達李振華徐國強孫浩奇王雅麗于春梅
浙江海洋大學學報(自然科學版) 2022年6期
關鍵詞:資源
印 瑞,周永東,梁 君,蔣日進,徐開達,李振華,徐國強,孫浩奇,王雅麗,于春梅
(浙江海洋大學海洋與漁業研究所,浙江省海洋水產研究所,農業農村部重點漁場漁業資源科學觀測試驗站,浙江省海洋漁業資源可持續利用技術研究重點實驗室,浙江舟山 316021)
大黃魚Larimichthys crocea 在我國海洋漁業中占有重要的經濟地位,是寶貴的海洋生物種質資源,也是浙江省最重要的底層經濟魚類之一[1],是我國著力恢復與重建的重要漁業對象。隨著資源保護和生態修復成為國家關注的重點,恢復與重建野生大黃魚資源愈加得到國家、地方政府以及科技工作者的關注。為了恢復東海大黃魚資源,多年來,政府曾采取了封港休漁、建立大黃魚幼魚保護區、開展大黃魚增殖放流等各種措施。而今,自然海區的大黃魚資源量開始逐漸增加,野生大黃魚重現東海的新聞屢見不鮮[2-3]。
中街山列島海洋牧場是浙江省首批國家級海洋牧場示范區,位于舟山漁場中心,是大黃魚、曼氏無針烏賊Sepiella maindroni 等海洋經濟生物重要的繁殖場所[4]。海洋牧場是一種典型的生產性生態系統,人工魚礁造成的水體營養物質的高可利用性能夠促進海洋牧場初級生產力的提高[5]。現代化海洋牧場的建設已經成為國家促進漁業產業轉型升級、緩解近海漁業資源衰退的重要舉措[6]。
自2006 年5 月中街山列島國家級海洋特別保護區成立以來,該海域逐漸成為漁業管理部門和相關科研單位關注的焦點。中街山列島海洋牧場區共投放超5 000 個人工魚礁單體,如今該海域表層水域逐漸清澈,魚類的資源密度也日漸增加[4,7]。
為了解現今中街山列島海洋牧場中大黃魚的資源時空分布,以及海洋牧場的建設對大黃魚分布的影響,本文應用廣義相加模型(generalized additive model,GAM)研究2021 年4 月—2022 年1 月中街山列島海洋牧場大黃魚資源的時空分布與環境因子的關系,以探究環境因子變化對大黃魚資源分布的影響,初步評估水環境狀況和大黃魚分布狀況,為探究海洋牧場的建設對大黃魚的資源恢復是否具有促進作用提供數據分析[5]。
1 材料與方法
1.1 數據來源
于2021 年4 月(春季)、7 月(夏季)、10 月(秋季)和2022 年1 月(冬季)在中街山列島海洋牧場海域及其鄰近海域進行4 個航次的底拖網漁業資源調查。調查海域范圍為30°~30.4° N,122.5°~122.9° E,共設置了18個調查站位(圖1),其中海洋牧場核心區10 個站位(H01~H10),海洋牧場外圍海域8 個站位(W01~W08)。
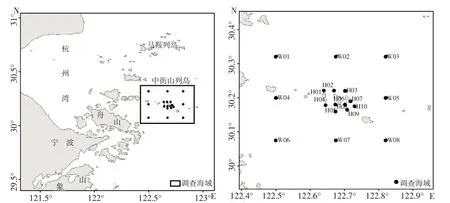
圖1 調查站位Fig.1 Map of survey stations
調查船為租用的漁業生產船,網具網口周長為25 m,網口高6 m,囊網網目尺寸為3.0 cm。平均拖速為2 kn,每個站位拖網時間約為30 min,所采集的樣品冷凍保存,帶回實驗室進行生物學測定。采用CTD水質儀同步測量調查海域各站位的溫度、鹽度和水深等環境因子。樣品的采集和分析均依照《海洋漁業資源調查規范》[8]進行。
1.2 數據分析
1.2.1 資源密度
根據掃海面積法計算大黃魚的資源密度,計算公式如下[9]:
式中:D 為資源密度(豐度,尾·km-2;生物量,kg·km-2);C 為實際調查漁獲量(數量,尾;質量,kg);q 為網具漁獲率(大黃魚屬于底層魚類,q 默認取0.5);A 為調查實際掃海面積,km2。
資源密度時空分布圖采用ArcGIS 軟件繪制。
1.2.2 GAM 模型構建
基于統計回歸分析的廣義相加模型(GAM)是一種分析漁業資源分布與環境因子間關系的模型方法[10-11]。可以反映響應變量與多個解釋變量之間的非線性關系。
本研究選取大黃魚的資源豐度作為響應變量,根據前期大黃魚分布習性的相關研究,選取底層海水溫度(SBT)、底層海水鹽度(SBS)、水深(Dep)、月份(month)這4 個因子作為初始解釋變量[12]。采用條件數和方差膨脹因子(VIF)度量多重共線性的強度,對選取的初始因子進行檢驗篩選,共線性強的2 個因子保留1 個,得到可以加入模型的因子[13]。
GAM 模型的表達式為:
式中:為防止零值的情況,對CPUE 值采取加1 后再對數化的處理;s 為自然立方樣條平滑函數;ε 為隨機變量[14]。
利用AIC(akaike information criterion)[10,15-16]準則檢驗中逐步回歸的方法,將因子逐步加入模型中擬合,AIC 值越小,表明模型的擬合效果越好。
通過F 檢驗評估因子的顯著性。通過R 軟件進行GAM 模型的構建及檢驗[17]。
2 結果
2.1 大黃魚資源的時空分布特征
2021 年中街山列島海域大黃魚的資源分布季節變化明顯,在海洋牧場的核心區與外圍區有明顯的資源分布差異。
秋季大黃魚資源量最高,平均豐度為91.83 尾·km-2,平均生物量為6.23 kg·km-2,在海洋牧場的核心區(H06、H09 站位)和外圍區(W01、W03、W06、W08 站位)均有分布,主要分布在外圍海域,其豐度與生物量分別是核心區的5.06 倍和3.9 倍。夏季其次,大黃魚的平均豐度為42.76 尾·km-2,平均生物量為2.50 kg·km-2,在海洋牧場的核心區(H05 站位)和外圍區(W06 站位)各出現1 次,外圍區的資源量稍高于核心區,核心區的大黃魚多為幼魚。冬季大黃魚資源量偏低,平均豐度為20.37 尾·km-2,平均生物量為1.62 kg·km-2,只出現在海洋牧場核心區(H02、H08、H09 站位)。春季最低,平均豐度為10.78 尾·km-2,平均生物量為1.40 kg·km-2,和冬季一樣,只出現在海洋牧場核心區(H08 站位)(表1、圖2)。
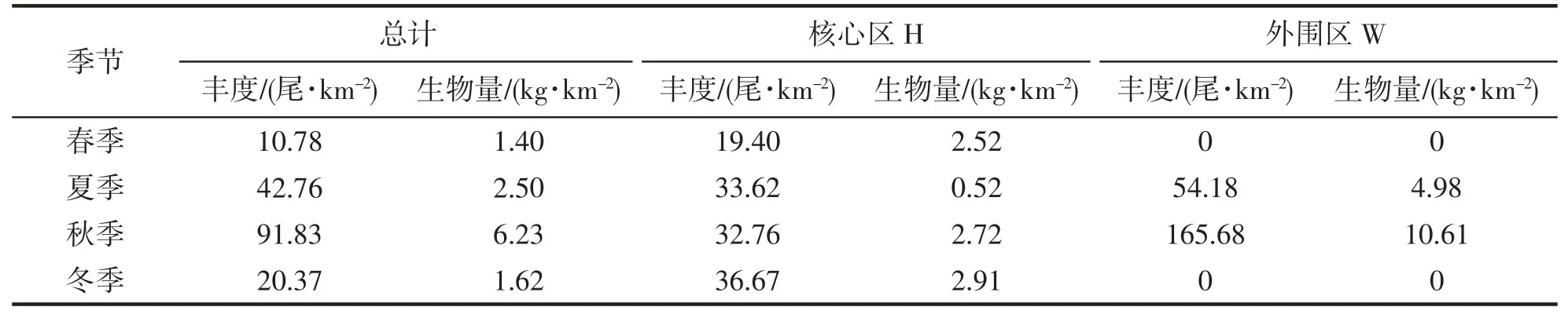
表1 大黃魚資源密度分布表Tab.1 Table of resource density distribution of L.crocea
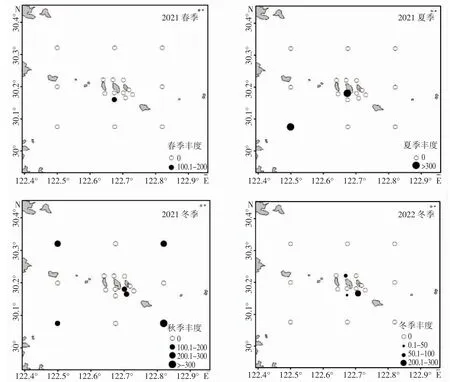
圖2 大黃魚資源密度分布圖Fig.2 Resource density distribution map of L.crocea
2.2 影響因子篩選
大黃魚為暖溫性近海洄游魚類,多生活在海水的中下層[18]。本研究初步選擇海域底層水溫、底層鹽度和水深作為環境因子,選擇月份作為時間因子。建模前的VIF 度量多重共線性強度分析,表明底層水溫與各環境因子具有極強的共線性,因此最終選擇月份、水深和底鹽加入GAM 模型進行模擬分析。最終擬合的模型表達式如下:
GAM 的累計偏差解釋率為28.84%,其中,月份、水深的累計偏差解釋率分別為8.47%和13.05%(表2)。
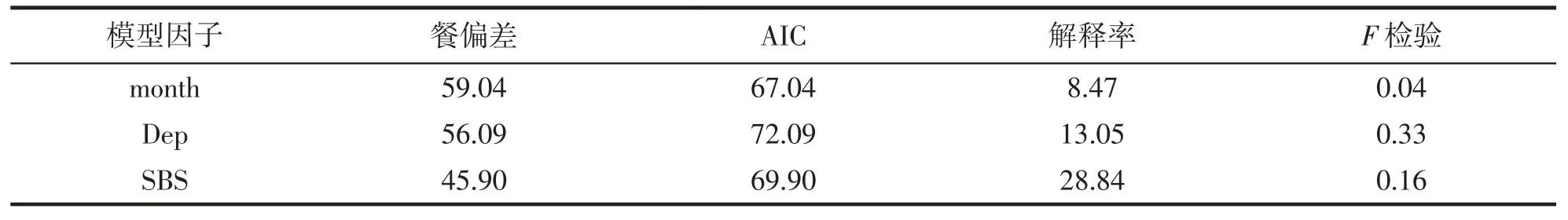
表2 GAM 模型擬合結果的偏差分析Tab.2 Analysis of deviance for GAM
2.3 影響因子分析
GAM 模型分析表明,水深和底鹽對大黃魚的資源分布影響顯著。本研究調查海域內,大黃魚出現在水深30~45 m、底鹽24~29 的海域中。由模型擬合結果來看,2021 年大黃魚出現率10 月(秋季)最大,4 月(春季)最低(圖3)。
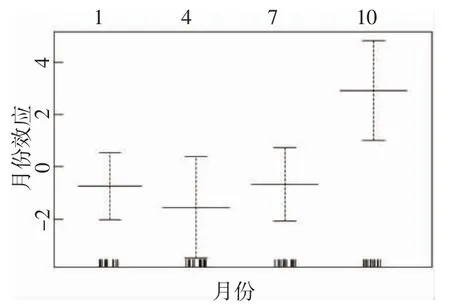
圖3 月份對大黃魚資源密度分布的影響圖Fig.3 Effect of month on density distribution of L.crocea
從環境因子對大黃魚資源分布的影響圖可知,水深和底鹽2 個因子均影響大黃魚資源密度分布。由水深與CPUE 的效應圖可知(圖4),在20~30 m 水深范圍內,大黃魚資源量隨水深的增加而急劇下降,在30~45 m 水深范圍內,資源量隨水深的增加而逐步上升,在大于45 m 水深的海域資源量又開始下降。
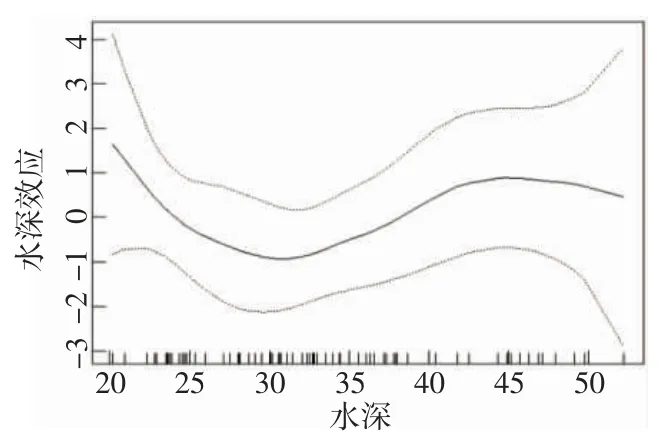
圖4 水深對大黃魚資源密度分布的影響圖Fig.4 Effect of water depth on density distribution of L.crocea
由底鹽與CPUE 的效應圖可知(圖5),當海水底層鹽度在22~24 范圍內時,大黃魚資源量呈下降的趨勢,在鹽度24~29 的范圍內,資源量隨底層鹽度的增加而略有上升,當底層鹽度大于29 時,資源量隨底鹽的增加而急劇下降。
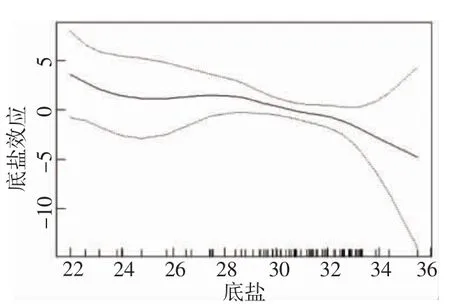
圖5 底鹽對大黃魚資源密度分布的影響圖Fig.5 Effect of bottom salt on density distribution of L.crocea
3 討論
3.1 大黃魚資源量的時空分布變化
棲息地是生物賴以生存和繁衍的空間,關系著生物的食物鏈和能量流[19]。棲息地包括越冬場、產卵場、索餌場和洄游通道。研究棲息地的環境條件是科學實施漁業資源保護與管理、恢復與重建的重要基礎[20]。
中街山列島海洋牧場及其鄰近海域中大黃魚資源分布具有顯著的時空分布差異和季節變化。本研究春、冬季大黃魚資源密度最低,且僅在海洋牧場核心區被采集到,多分布在廟子湖及其附近人工魚礁投放海域。夏季大黃魚資源密度高于春、冬季,在海洋牧場的核心區和外圍區各出現1 次,但核心區出現的多為大黃魚幼魚。秋季大黃魚在海洋牧場外圍區的資源密度遠高于核心區,在核心區大黃魚分布在人工魚礁投放海域,在外圍水域大黃魚的分布有明顯的種群外移現象。研究發現,在中街山列島海洋牧場核心區,大黃魚四季均有分布,且多分布在島嶼以南的人工魚礁投放海域;在外圍海域,僅在夏、秋季發現大黃魚。
根據以往研究,大黃魚在生殖期和越冬期具有較高的集群性[19-21]。大黃魚產卵期為5 月上中旬至6 月下旬,喜在沙泥和軟泥底質的區域產卵,完成產卵后,就近分散于產卵場外圍;10 月“秋宗”大黃魚開始產卵,并隨著水溫下降,大黃魚開始向越冬場進行越冬洄游,越冬期為當年12 月至翌年3 月,大黃魚的越冬場位于江外、舟外漁場等東海外海高鹽水和江浙沿岸水交匯的混合水區;4 月起,隨著臺灣暖流的上升增強,水溫升高,大黃魚進入產卵洄游階段,進入產卵場產卵[22]。
這與本研究中大黃魚資源的時空分布與季節分布幾乎一致,解釋了夏、秋季海洋牧場外圍區大黃魚資源量高于核心區的現象。在大黃魚的產卵期,其多聚集在人工魚礁投放海域,表明中街山列島海洋牧場的建設給大黃魚提供了一個適宜的棲息、繁衍海域。
3.2 環境因子對大黃魚資源分布的影響
棲息地環境的變化是影響生物時空分布的主要因素[23],水深、水溫、鹽度對魚類的生存、繁衍及洄游分布等有不同程度的影響[24]。魚類的生長發育具有適溫范圍和適鹽范圍,當棲息環境適宜時,會刺激魚類親體性腺發育成熟并產卵[25],當環境不適時,則不利于魚卵孵化和仔稚魚的生長發育,會導致魚類早期群體死亡率的上升。而水深的變化則會影響棲息海域的溫度、鹽度、壓強等因素,以此對魚類的資源分布產生影響。以往研究表明,大黃魚的最適水溫為15~22 ℃[22],最適鹽度在春季為17~28,在秋季為26~33[19~21]。
本研究中經模型擬合顯示水深和底鹽對于中街山列島海洋牧場大黃魚的資源分布具有顯著影響。中街山列島及其周圍調查海域的水深范圍為20~50 m,底鹽范圍為22~36。在調查海域,隨著水深的增加,大黃魚的資源量呈先下降再上升后又下降的趨勢,適宜水深為30~45 m,在45 m 深海域達到最大值,表明2021 年在中街山列島海域大黃魚的最適水深為45 m。在該海域,隨著底層海域鹽度的增加,大黃魚的資源量同樣呈現先下降再上升后又下降的趨勢,適宜底鹽為24~29,當底層鹽度為29 時達到最大值,表明2021 年在中街山列島海域大黃魚的最適底鹽為29。
大黃魚屬于集群性較強的魚類,特別是在產卵場和越冬場,具有較高的集群性。影響中街山列島海域大黃魚資源量的關鍵環境因子是水深和底鹽。
4 總結
大黃魚資源豐盛時期,其生活史隸屬于K選擇型,個體較大,生長緩慢,生命周期較長,產卵剩余群體多于補充群體,資源穩定性較強,但一旦遭受嚴重的捕撈破壞,就很難得到恢復[26]。如今,由于資源嚴重枯竭,目前大黃魚生活史類型已經從K 選擇型演變為r 選擇型,生命周期短,資源穩定性差[26]。
因此,要實現大黃魚資源的有效保護和恢復,需要充分掌握大黃魚的生物學特性和對棲息地的環境條件的選擇性,建立合適的海洋牧場,調整相應的休漁期、禁漁區以及可捕規格,進行合理的增殖放流,加強漁政管理,才能更好地、更科學地對其實施保護與恢復措施[22]。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
