萊州灣魚卵、仔稚魚的群落結構特征及其環境影響因素研究
2022-02-25 06:58:42于廣磊李燕宋秀凱黃華何健龍楊艷艷劉愛英
水產學雜志 2022年1期
關鍵詞:區域
于廣磊,李燕,宋秀凱,黃華,何健龍,楊艷艷,劉愛英
(1.山東省海洋資源與環境研究院,山東省海洋生態修復重點實驗室,山東 煙臺 264006;2.南水北調中線建管局北京分局,北京 100038;3.煙臺市海洋經濟研究院,山東 煙臺 264003)
萊州灣是典型的半封閉性內海和中國北部環渤海的三個主要灣之一。萊州灣海域及灘涂濕地(37°04'09″~37°36'00″N,118°57'47″~119°51'22″E)總面積為3 770 km2,是我國重要的漁業生產基地,注入萊州灣的內陸河流超過10 條,黃河和小清河是萊州灣陸源輸入的主要來源。近年來,在高強度捕撈壓力下萊州灣主要經濟魚類資源日趨衰退,捕撈對象日趨小型化、低齡化,生命周期短、低營養級種類偏多,對漁業資源的利用正在向低值、劣質轉化,漁業資源正朝著不利于利用的方向發展,漁業資源質量的總體狀況令人擔憂[1];黃河的徑流量和輸沙量劇烈減少,水流量和沉積物量的改變以及陸源污染輸入引發河口及沿海區域地質、地貌、生態和生物地球化學響應[2-4]。這些區域生境的改變嚴重影響了萊州灣魚卵仔稚魚群落結構及空間分布[5]。
本文于2012—2014 年選取設在萊州灣的20個站位的調查數據及相關資料,分析期間該海域的魚卵、仔稚魚數量分布和多樣性的年際變化,結合區域的水文和其他環境要素變量特征,刻畫生境與萊州灣春季產卵場形成、分布的特定聯系。
1 材料與方法
1.1 調查航次和站位分布
按照均勻分布、重點兼顧的原則,于2012—2014 年每年5 月在萊州灣海域布設20 個站位,進行3 個航次的調查。
1.2 樣品采集、處理及分析
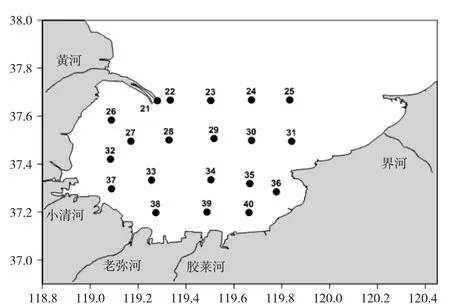
圖1 調查站位Fig.1 Survey stations
魚卵、仔稚魚樣品采集及分析均按照《海洋生物生態調查技術規程》[6]進行。樣品用口徑50 cm、長280 cm、38GG 篩絹制成的大型浮游生物網采集,于每個調查站表層水平拖網,每站拖網10 min,拖網速度為2.0 nmile/h。調查樣品以5%的福爾馬林溶液固定,帶回實驗室,單獨分揀魚卵、仔稚魚標本,在解剖鏡下鑒定各個站點樣品的種類,計數個體和發育階段。根據魚卵、仔稚魚的形態特征鑒定種類[7,8]。魚卵仔稚魚的密度以水平拖網采獲的總個體數除以濾水量計算,濾水量等于拖速乘以拖網時間,再乘以網口面積。由于海流、風浪等客觀因素,水平拖網速度難以嚴格控制,以每站位的實際數量(粒、尾)為指標進行比較。
溫度、鹽度、pH、溶解氧用YSI-professsional plus 進行現場測定。海水樣品采集和分析均按照《海洋監測規范》(GB 17378-2007)進行。浮游植物、浮游動物樣品的采集和鑒定均參照GB 17378.7《海洋監測規范》中的生態調查方法進行。
1.3 數據處理
用Primer 6.0 軟件對2012—2014 年間魚卵仔稚魚生物多樣性進行分析計算,用SPSS19.0 軟件分析數據的Person 相關性。采用Surfer11.0 軟件繪制魚卵和仔稚魚數量平面分布圖。
1.4 魚卵群落結構指數的處理方法
分析2012—2014 年萊州灣春季魚卵Margalrf種類豐富度指數(D)、Shannon-Wiener 多樣性指數(H)、Pielou 均勻度指數(J)和優勢度(Y)。
Shannon-weaver 多樣性指數(H'):
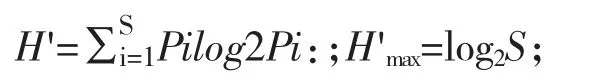
式中,Pi=ni/N,為第i 種在樣品中的細胞豐度概率;S 為總種數。
式中ni為第i 種的總個數;fi為該種在各樣品中出現的頻率;N 為全部樣品中的總個體數。
2 結果與分析
2.1 萊州灣魚卵、仔稚魚種類組成
3 個航次調查共采集到302 373 粒魚卵和2 912尾仔稚魚,計27 種魚類。其中,鑒定到種的有16種,隸屬于5 目11 科16 屬,還有6 個種僅鑒定到屬,5 種僅能鑒定到科。在已鑒定的16 種魚卵、仔稚魚中,鱸形目9 種,占56.3%;鯡形目3 種,占18.8%;鰈形目1 種,鲉形目1 種,鯔形目2 種;從科一級看,蝦虎魚科、鳀科和鯔科所占比例較高。
2012 年共采集到21 種魚卵、仔稚魚(含鑒定到種、屬),魚卵13 720 粒,仔稚魚956 尾;2013 年共采集到11 種魚卵、仔稚魚(含鑒定到種、屬),魚卵1 675 粒,死卵1 282 粒,占比高達76.5%,未采集到仔稚魚;2014 年共采集到12 種魚卵、仔稚魚(含鑒定到種、屬),魚卵286 978 個,仔稚魚1 956 尾,死卵33 512 粒,占比達11.7%。24 監測位點,采集到鳀卵223 880 粒,占到整個航次魚卵總量的78.0%。
2012 和2014 年春季萊州灣海域魚卵數量中,斑鰶(Konosirus punctatus)均作為優勢種;在2012年共采集到斑鰶卵6 078 粒,占當年調查采集魚卵總數的44.3%;2014 年斑鰶魚卵數量為11 458 粒,站位出現頻率達到85.0%;鳀卵2012 年為第3 優勢種,2014 年是第1 優勢種,共采集到鳀卵237 686粒,占當年調查采集魚卵總數的82.8%,站位出現頻率達到75.0%。
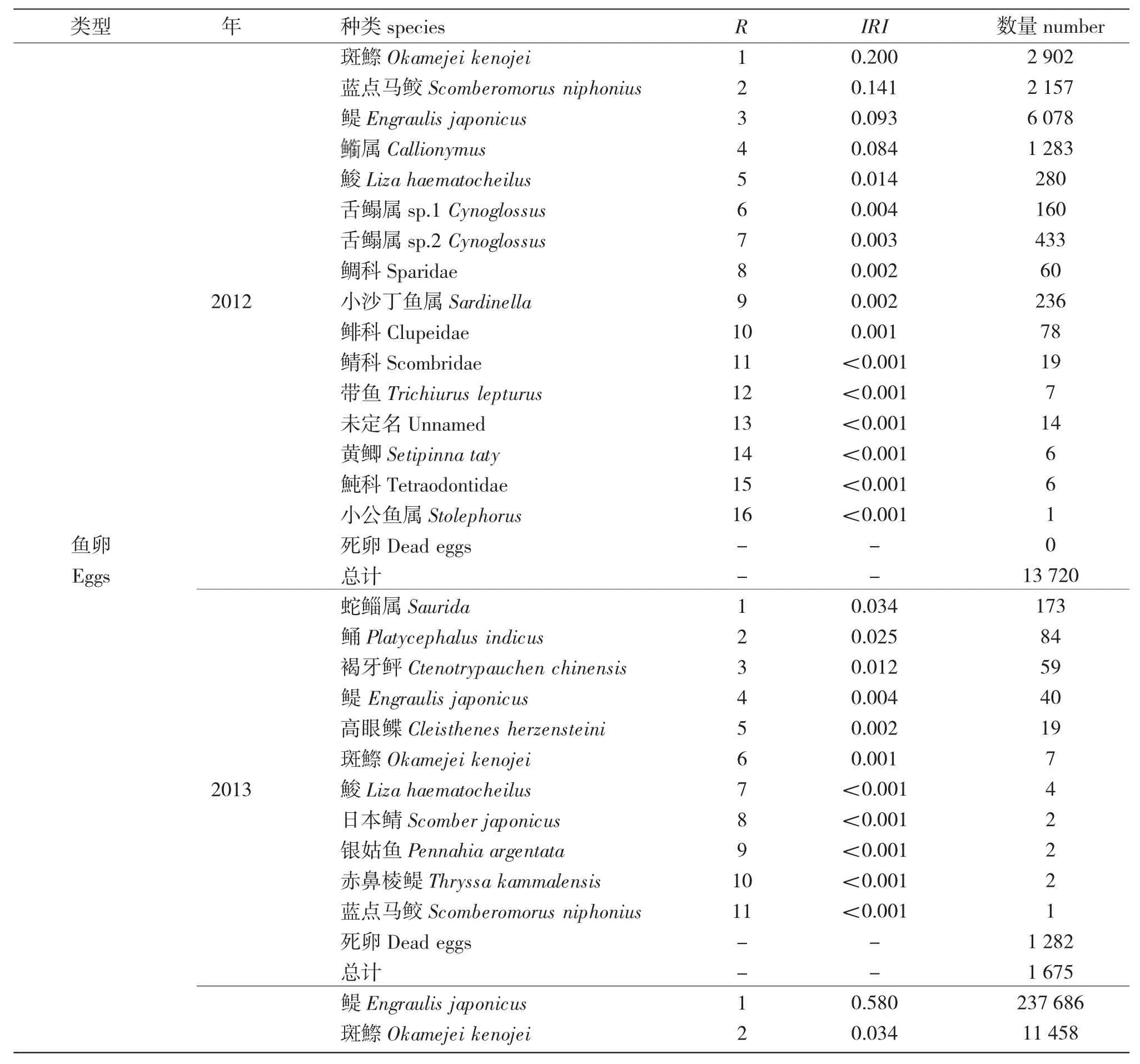
表1 2012—2014 年萊州灣魚卵、仔稚魚種群組成、數量及魚卵種類優勢度Tab.1 Population composition and quantity of fish eggs and larvae and species dominance of fish eggs in Laizhou Bay from 2012 to 2014
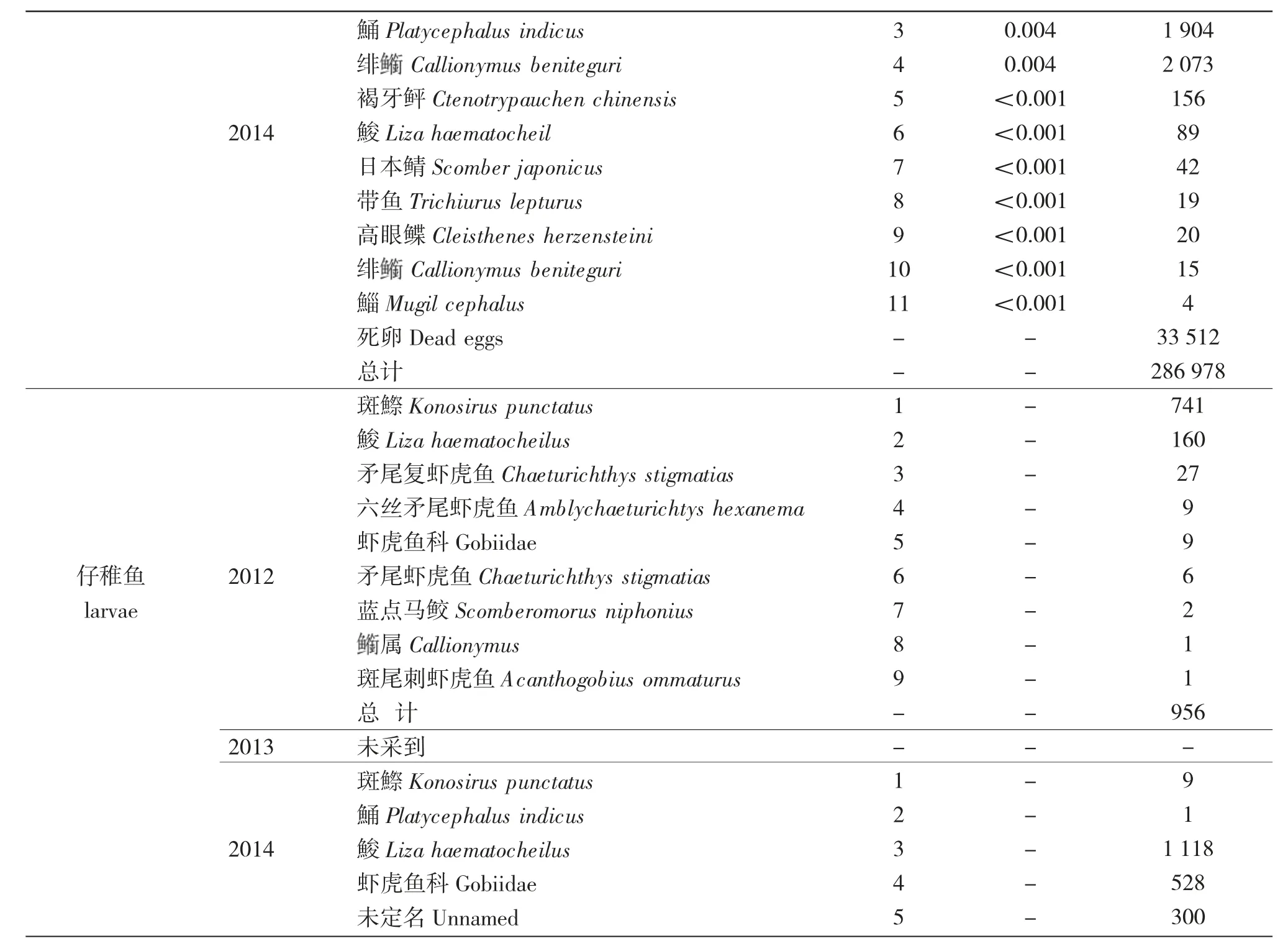
續表1
徐兆禮等[9]在研究東黃海浮游動物優勢種,將優勢度Y≥0.02 時的浮游動物確定為優勢種。本文按照此標準進行分析,得出2012—2014 年調查海域魚卵的優勢種共有6 種(屬),依次為鳀、斑鰶、藍點馬鮫、屬、蛇鯔屬、鲬。這些種類大多數為鯡形目、鱸形目和鲉形目,豐度占到魚卵總豐度的87.8%。
2.2 2012—2014 年萊州灣魚卵、仔稚魚數量的平面分布
由圖2 可以看出,魚卵、仔稚魚平面分布極不均勻,沿岸海域數量大于遠海。其中,黃河、小清河和膠萊河河口附近數量較多,萊州灣灣口海域數量較大,中部海域數量較少。
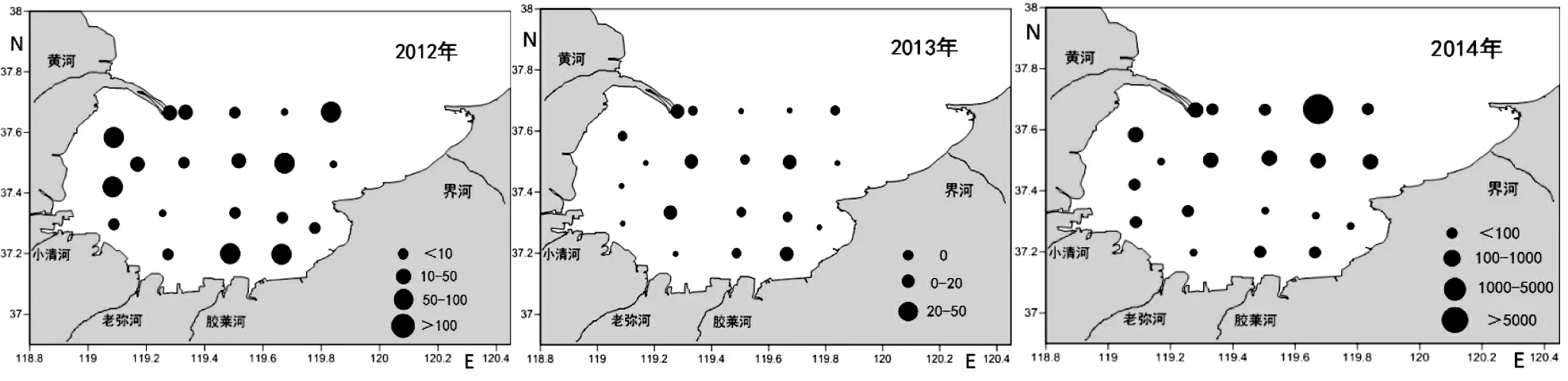
圖2 2012—2014 年萊州灣魚卵、仔稚魚數量的平面分布Fig.2 Horizontal distribution of fish eggs and larvae in Laizhou Bay from 2012 to 2014
2012 年,黃河入海口26 站位采集到魚卵、仔稚魚數量最大,達894.7 粒(尾)/100 m3,黃河口附近22、21、27、32 站位數量也都在50 粒(尾)/100 m3以上,區域整體豐度較高;膠萊河河口附近39、40 站位數量均在100 粒(尾)/100 m3以上;萊州灣東北灣口附近25、30 站位數量在200 粒(尾)/100 m3以上,整體豐度較高;小清河、老彌河及近岸遠海區域數量在50 粒(尾)/100 m3以下,豐度較低。2013 年,魚卵、仔稚魚數量整體豐度不高,除黃河口附近28 站達到53.3 粒(尾)/100 m3,其余各站數量都在50 粒(尾)/100 m3以下。此次航次沒有采集到仔稚魚樣品,水平分布上以黃河口、膠萊河河口、萊州灣東北灣口為魚卵數量較大區域。2014 年,萊州灣魚卵、仔稚魚監測站整體豐度較高,數量在34.8~56 160.0 粒(尾)/100 m3,其中24 站位高達56 160.0 粒(尾)/100 m3,位于萊州灣灣口區域,萊州灣灣口、黃河口、膠萊河河口、小清河口為魚卵、仔稚魚數量較大區域,數量均在100 粒(尾)/100 m3以上,老彌河及近岸遠海區域魚卵、仔稚魚數量較少。
2.3 魚卵、仔稚魚數量與主要環境因子相關關系
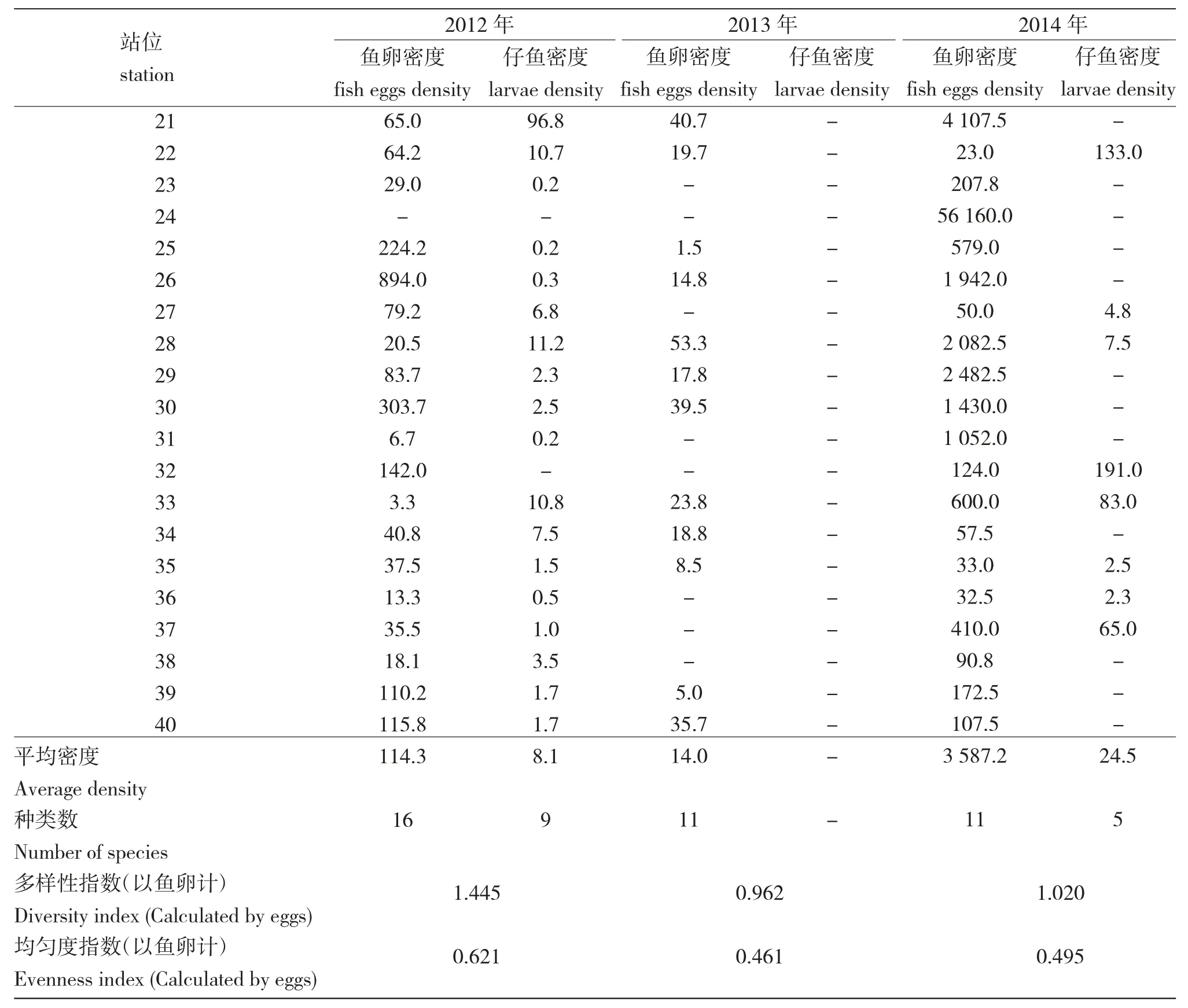
表2 2012—2014 年萊州灣魚卵、仔稚魚數量的平面分布及生態學特征Tab.2 Horizontal distribution and ecological characteristics of fish eggs and larvae in Laizhou Bay from 2012 to 2014
用SPSS 軟件分析了萊州灣調查海域的魚卵、仔稚魚數量同水溫、pH、鹽度、溶氧、營養鹽、浮游生物數量等各主要生境要素的相關關系(表3)。由表3 可知:2012 年仔稚魚數量與浮游植物含量呈明顯正相關(r=0.891,P<0.01),2013 年魚卵數量與化學需氧量呈明顯正相關(r=0.564,P<0.01)。由于調查區域所采集的魚卵、仔稚魚種類多數為廣鹽性的種類,鹽度的變化對仔稚魚影響不敏感(P>0.05)。2012年萊州灣區域漁產量及浮游植物對比2011/2013年,浮游植物2012 年平均密度達到20.9×104個/m3,2009 年5 月平均密度為8.05×104個/m3[11],2011 年5 月平均密度為19.6×104個/m[3][山東省海洋環境監測數據庫資料],2013 年5 月平均密度僅達到1.5×104個/m3。2012 年調查區域采集的仔稚魚中優勢種主要為斑鰶,達741 尾,占77.5%。斑鰶是暖水性魚類,主要棲息在內灣淺海區,是我國沿海常見的小型經濟魚種,主要以浮游植物為食,也食浮游動物和底棲生物[12],幼魚期攝取小粒徑的硅藻、甲藻等為[13]。2012 年化學需氧量反映了水中受還原性物質污染的程度,常作為有機物相對含量的綜合指標,而化學耗氧有機物是引起海水富營養化的重要因子,可為海洋浮游植物生長提供碳源,促進浮游植物生長[14]。大量文獻資料表明,魚卵、仔稚魚的分布不僅與浮游動物、浮游植物等餌源類生物單位水體的數量有關,也與水溫、pH、鹽度、溶氧、營養鹽等環境因素以及潮汐、洋流等物理海洋要素有相關性,但由于海水環境的復雜性、流通性及相關物理海洋數據的缺乏,還無法明確分析兩者的確切內在聯系。
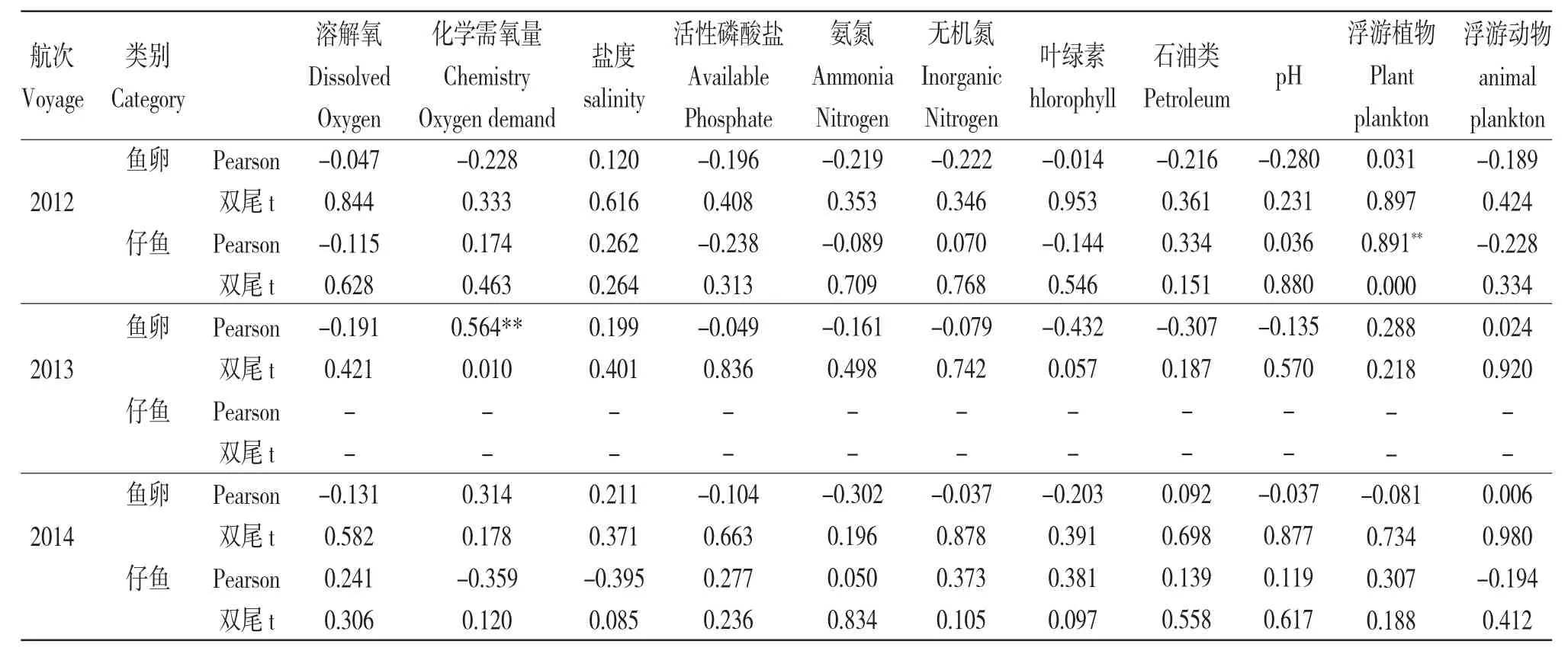
表3 2012—2014 年萊州灣海域魚卵、仔稚魚數量與各環境因素的積矩Pearson 相關關系Tab.3 Pearson correlation between the density of fish eggs and larvae in Laizhou Bay waters and environmental factors from 2012 to 2014
3 討論
3.1 生物種群及數量的變化
與2008 年同期調查數據相比[15],萊州灣海域5月魚卵、仔稚魚種類從2012 年的21 種下降到2013年的11 種和2014 年的12 種;與2008 年魚卵、仔稚魚種類的25 種相比總體呈下降趨勢,優勢種也由以斑鰶和鳳鯽為主變為以鳀、斑鰶為主。2012—2014 年,萊州灣魚卵、仔稚魚以鳀、斑鰶等暖水中上層以浮游生物為食的小型魚類為主,鲬、蛇鯔等雜食性魚類次之,這與萊州灣海域漁業資源結構正在向經濟價值低、個體小、營養層次低的小型中上層魚類類群演替的變化趨勢相吻合[16],浮游食物鏈漁業資源已取代底棲食物鏈漁業資源成為最主要的漁業資源;碎屑食物鏈漁業資源生物量百分比呈上升趨勢,近年來已成為繼浮游食物鏈漁業資源的第二大類漁業資源[17]。過度捕撈以及人為對海洋生態環境的改變是導致魚卵、仔稚魚種類及生態習性發生變化的重要原因[18]。
3.2 環境因子對魚卵、仔稚魚資源的影響
魚卵和仔稚魚基本上營隨波逐流的漂流生活方式,因此,魚卵和仔稚魚的分布與海流、潮汐以及水溫和鹽度分布等物理海洋學要素的關系密切[19];河流沖淡水最直接影響近岸鹽度時空分布。鹽度和溫度是影響魚卵新陳代謝的主要因素,其變動直接影響仔稚魚的生產和形態發育,近海低鹽水系對魚卵的繁殖與孵化有重要影響[20-24]。本研究中春、夏季魚卵、仔稚魚優勢種分布與表層鹽度均無顯著相關性,這說明魚卵、仔稚魚的分布可能受到海流等其他因素的影響。對照歷史調查數據及近期研究數據,顯示漁業資源對產卵場溫鹽適應范圍在擴大:產卵場受污染、圍填等破壞和長期捕撈擾動,很多種類被迫由原來的最適產卵場擴展到周邊區域產卵,同時,優良產卵場的衰減,產卵密集區的范圍有逐漸縮小趨勢[18]。
與20 世紀90 年代相比,萊州灣產卵密集區萎縮為萊州灣西南部和萊州灣東北部兩個產卵密集區[25]。魚卵、仔稚魚自主游泳能力弱,海流對其分布的影響較為顯著[26]。黃河口北部的魚卵、仔稚魚受逆時針環流系統的影響,在余流尾部形成高密度中心;黃河口南部區域魚卵、仔稚魚在靠近萊州灣一側聚集,可能受該區域逆時針環流系統的影響[27]。
仔稚魚具備了主動追索食餌的習性,其分布除了與物理海洋學要素密切相關外,也與浮游生物分布有一定關系[28]。河流攜帶大量營養鹽入海,為海洋生物繁育提供了豐富營養物質,孕育了較高的初級生產力,為海洋生物幼體的生長發育提供了豐足的食物。許多魚類幼體階段生活在該生態系統中,使河口海域成為大量海洋魚類重要的產卵場、育幼場,孕育著種類不斷更替的魚類資源[29]。黃河實施調水調沙管理措施后,增加了黃河入海水量和3~6月營養鹽的入海通量,為浮游植物的生長繁殖提供了豐富的營養物質,黃河口附近的魚卵和仔魚數量也逐年增加,河口及近海區域生物群落結構略有改善[3]。小清河流域的治理,使河口區域海洋生態環境好轉,污染狀況有所緩解[30]。水質的改善提高了小清河口臨近海域初級生產力,對恢復魚類產卵場起到重要作用[31]。
猜你喜歡
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經大學學報(2015年3期)2015-12-10 03:49:15
