納米金屬顆粒在土壤-植物系統中的遷移轉化及生物效應研究進展
2022-02-27 10:23:56湯岑鵬趙峰娃青1
環境科學研究 2022年2期
關鍵詞:植物
疏 茂,湯岑鵬,趙峰娃,趙 青1,2,*
1.廣東省科學院生態環境與土壤研究所,廣東省農業環境綜合治理重點實驗室,廣東 廣州 510650
2.華南土壤污染控制與修復國家地方聯合工程研究中心,廣東 廣州 510650
3.中國海螺創業控股有限公司,安徽 蕪湖 241070
4.中國科學院沈陽應用生態研究所,遼寧 沈陽 110016
人工納米材料被廣泛應用于生產和生活多個領域,如化妝品、防曬霜、紡織品、太陽能電池材料、催化劑以及納米診療[1]等,其在整個生命周期中將不可避免地進入環境.自然界存在的金屬離子可以在光、熱作用下轉變為納米顆粒.Ag+和Au+在太陽光照及/或DOM 存在環境中可轉化生成納米顆粒[2].同時,納米材料常被用于污染土壤的修復[3-6],如負載納米零價鐵的碳材料能有效去除重金屬Cr(Ⅵ)[5],改性納米零價鐵可提高稻田As 污染土壤的修復效能[6].這些人工及天然納米顆粒可能引發的生態環境問題是近年來研究關注的熱點.
由于尺寸小,納米顆粒可以突破血腦屏障,進入血液、淋巴循環,富集于肝臟、心臟、腎臟及中樞神經系統,誘發炎癥、細胞凋亡與組織壞死等毒性效應.它們不僅能引起DNA 損傷等遺傳毒性[7],還會產生活性氧物種(ROS)造成脂質過氧化,從而導致細胞損傷[8].不同于宏觀材料,納米顆粒(<100 nm)具有一些獨特的理化性質,如界面效應、小尺寸效應及宏觀量子隧道效應等[9].因此,納米顆粒誘發產生的ROS隨比表面積增加呈指數增長,將引發更嚴重的生物損傷.
納米金屬顆粒不僅能釋放同種金屬離子產生毒性,顆粒本身也可能具有更高的毒性效應.例如:穩定性較高的納米二氧化鈦對小麥的毒性主要來源于材料自身的毒性效應[10];穩定性較弱的納米氧化鋅則可溶出鋅離子對小麥[10]或水藻[11]產生毒性;α-Fe2O3中解離的Fe3+會影響藻類生長.但也有研究[8]發現,納米鋅對黑麥草的生物毒性主要由顆粒自身造成.納米金屬顆粒進入植物體內會產生ROS,引起植物細胞的損傷和死亡,也可能影響營養元素運輸和吸收,從而影響植物生長[12].對于納米銀、納米氧化鋅、納米氧化鈰、納米氧化鈦等金屬納米顆粒,無論能否釋放金屬離子,其自身的毒性都不容忽視.目前,納米金屬顆粒的毒性效應機制尚存爭議,一些關鍵問題有待于深入研究.我國對于納米金屬顆粒生物學效應的研究仍處于起步階段,揭示納米金屬顆粒的環境行為及生物學效應是發展納米產業迫切需要解決的課題.
土壤是納米金屬顆粒在環境中重要的源和匯,在其生物地球化學循環中起著關鍵作用.土壤中的納米金屬顆粒可下滲進入地下水,也可能被動植物吸收后經食物鏈傳遞富集.納米金屬顆粒在食物鏈上的傳遞存在生物放大效應,放大系數達5.32[9].植物是生態系統的初級生產者,作為食物鏈起始營養級,植物對納米金屬顆粒的吸收和轉化,不僅影響植物自身安全,而且可能影響食物鏈中各級消費者.因此,研究納米金屬顆粒在土壤-植物系統中的遷移轉化、在植物中的運輸過程和機制、在植物中的生物轉化及其對植物的生物學效應對于納米金屬顆粒的生態風險評價具有十分重要的理論和現實意義.
1 納米金屬顆粒在土壤中的遷移轉化
關于納米材料遷移的研究主要集中在土壤和地下水,如飽和條件下均質砂土中的遷移等[13].然而,納米材料在進入地下水之前必先通過一個非飽和區域(即包氣帶),在非飽和多孔介質中的保持和移動決定了其對地下水的污染程度[14].針對非飽和流的研究,多采用物理和化學性質較均勻的砂土在穩態流條件下進行[14].現有試驗結果和模擬模型在一定程度上有助于了解納米材料環境行為,但是尚缺乏對真實環境條件及多種因素同時作用的模擬.
土壤具有復雜的結構系統和化學異質性(如質地分層、黏粒團聚、金屬氧化物及DOM 在不同土壤組分中不均勻分布等),水分在土壤中的運動也以非穩態(瞬態)的滲透、排水和蒸發過程為主.土壤的異質性能夠影響納米膠體和土壤的相互作用.例如,土壤干濕交替過程中水的瞬態脈沖(水通量和速度同時變化)類似于“水力激活器”,能夠改變孔隙水的幾何布局及孔隙中液-氣界面的能量梯度和狀態,并使原來保持在液-固和液-氣界面處的納米膠體獲得動量,克服阻礙其運動的“能壘”,從而再移動和進一步擴散[15].因此,在非穩態不飽和流情況下,納米材料傳輸的程度可能會遠超過當前穩態流模型所預測的水平.此外,由于納米材料的大小和表面性能存在顯著差異,其對土壤異質性、孔隙水分飽和度和水流穩定性的響應可能會有很大不同.但這方面的研究尚鮮見報道.因此,納米材料在自然環境中非穩態不均勻流模式(如降雨)和空間異質吸附情況下的遷移是有待研究的重要科學問題,對其進行系統闡釋,有望對膠體不飽和非均勻遷移理論、試驗方法、數學模型和機制分析等諸多方面研究進行有益補充.
受各種環境因子制約,納米金屬顆粒在環境中的賦存形態復雜(見圖1).為滿足具體應用需求而設計生產的結構性質(表面官能團化、尺寸、晶型結構)千差萬別的納米金屬顆粒,在生產和應用過程中被排放進入環境.這些納米金屬顆粒又會隨著環境條件的變化而發生形態轉化,這些形態轉化過程主要包括但不局限于以下幾個方面.

圖1 納米金屬顆粒的形態轉化Fig.1 Morphological transformation of metal nanoparticles
a) 吸附/解吸.納米金屬顆粒可以與環境中的可溶性有機質(DOM,如腐殖酸等)發生吸附,形成有機結合態的納米金屬/金屬氧化物.影響DOM 在納米金屬顆粒上吸附的主要因素包括吸附劑性質(比表面積、表面電荷、晶型、表面官能團及團聚情況)、DOM 結構和性質(親疏水性、表面電荷、分子量、羥基和羧基含量和空間結構)以及環境條件(pH、離子強度、離子價態、溫度)[16].納米金屬顆粒與DOM 的吸附機理主要包括靜電作用、配位作用、疏水作用、熵變、氫鍵作用及陽離子的架橋作用[17].在這些作用中,配位作用即納米金屬顆粒表面與DOM 的羥基羧基官能團進行配位交換被認為是最主要的作用.
b) 分散/沉降.游離或有機結合態的納米金屬/納米金屬氧化物顆粒可以與其自身或者環境中的礦物發生團聚,形成不同水和離子半徑的團聚體,這些團聚體既有可能由于空間位阻、靜電斥力作用懸浮在溶液中,也可能發生沉降,主要取決于團聚體顆粒之間引力和斥力的相對大小.多價陽離子可顯著降低團聚體的穩定性,如納米銀的臨界絮凝濃度(CCC)根據單價和多價陽離子的不同可以產生2 個數量級的變化[18-19].DOM 也可以顯著影響團聚體的穩定性,在低濃度陽離子存在條件下,DOM 的加入可以顯著增強納米銀、納米二氧化鈦的團聚體穩定性[20].
c) 解離.納米金屬顆粒在環境中可以逐漸解離釋放出金屬離子.該過程受離子強度、pH、腐殖酸等環境條件的影響.例如,在水環境中,pH 的變化會導致離子強度發生改變,使得氧化鋅納米顆粒聚集尺寸發生改變,進而影響其解離過程[21].在等電點時,氧化鋅納米顆粒多呈現聚集狀態,但pH 降低將促進Zn+解離[22].
d) 氧化/還原.在環境中氧化還原條件影響下,納米金屬顆粒自身與溶液中游離的金屬離子之間可以發生相互轉變.DOM 也會對納米金屬顆粒產生影響,例如,將聚集的納米銀轉變為銀離子,或是在光的作用下產生自由基,從而影響納米金屬顆粒的各種性質.需要指出的是,溶液中一些游離的重金屬離子(銀離子、三價金離子等)可以在光照或者加熱條件下被DOM 還原成納米金屬顆粒[23].
綜上,納米金屬顆粒在環境中的形態轉化是一個復雜的動態平衡過程.體系中某一環境條件的改變,或者納米金屬顆粒某一賦存形態的變化都會導致該平衡被打破,使得體系中納米金屬顆粒各賦存形態的含量發生改變.以往研究大多關注某一形態轉化過程,而忽視了體系整體的平衡變化.因此,納米金屬顆粒在土壤中各形態轉變過程的耦合效應是有待研究的重要科學問題.納米金屬顆粒各形態轉化過程中各參數之間的聯系、環境條件(pH、離子強度、DOM)的改變對其形態轉變的影響等問題是研究納米金屬顆粒在土壤-植物系統中遷移轉化和毒性效應的前提和基礎.
2 納米金屬顆粒在植物中的運輸過程及機制
植物是納米金屬顆粒進入食物鏈的開端.研究[24]發現,在植物的根、芽等部位都觀察到了納米金屬顆粒的存在,表明它們能夠被植物吸收并在植物體內運輸.生物富集系數(BAF),即物質在生物體組織中的濃度與其環境濃度的比值,常被用來衡量植物吸收運輸納米金屬材料的能力.比較植物對納米金屬顆粒的BAF 值發現,不同植物對同一納米金屬顆粒,同種(品種、生態型)植物對不同納米金屬顆粒,甚至同一植物對同種納米金屬顆粒(尺寸或表面性質不同)的BAF 值均差異顯著[25].例如,納米銀(Ag NPs)、納米氧化鈰顆粒(CeO2NPs)和納米氧化鋅顆粒(ZnO NPs)在大豆根部的BAF 分別為0.008 25、0.210 00 和1.173 52,而ZnO NPs 在小麥、玉米、青豆根部的BAF 則為3.432 3、0.450 0 和1.380 0[25].造成這種差異的原因可能是環境中納米金屬顆粒各賦存形態在植物體內運輸機制不同.納米金屬顆粒在復雜環境條件下發生形態轉化,形成具有不同理化性質的多種賦存形態.由于納米材料的理化性質決定其與蛋白質的相互作用,因此,與植物體內金屬轉運蛋白結合較強的納米金屬顆粒更容易在植物體內運輸,從而可能具有較高的生物富集能力.
在土壤中,納米金屬顆粒首先吸附在植物根部,此過程會受到植物根系周圍環境條件與納米金屬顆粒自身性質的影響.首先,不同植物或植物在不同生長階段的根際分泌物會對納米金屬顆粒在植物根部的吸附起到促進或抑制作用[26].而吸收納米金屬顆粒(如納米銅)又能引起植物(黃瓜)根系分泌物代謝的顯著變化[27].其次,納米金屬氧化物顆粒大小會對吸附過程造成影響,但具體機制尚不清晰.吸附在根部的納米金屬顆粒會向植物體內轉運,這一過程有兩種途徑(見圖2):一是質外體途徑,納米金屬顆粒穿過一系列屏障,即角質層、表皮、皮質、內皮層、凱氏帶到達木質部向上運輸;二是共質體途徑,納米金屬顆粒通過胞間連絲進入細胞[28].研究[29]發現,氧化鈰納米顆粒主要通過質外體運輸途徑進入玉米.類似地,納米氧化鑭顆粒也通過質外體運輸進入黃瓜(Cucumis sativusL.)體內[30].納米氧化鐵和納米氧化鋅在大豆和玉米中的運輸是經木質部實現的[31].而金納米顆粒在毛白楊(Populus tomentosa)中的運輸則由木質部和韌皮部共同完成,且在根細胞間的韌皮部胞間連絲內積累(見圖2)[32].納米金屬顆粒的尺寸和表面電荷也會影響植物對其的吸收.例如,表面電荷不同的AuNPs 在水稻根部的吸收順序為AuNP1(+)>AuNP2(0)>AuNP3(?)[33].小尺寸ZnO NPs 能夠穿過細胞壁進入細胞質,而正電荷能夠增強ZnO NPs 在植物細胞壁上的吸附[34].這些已有報道大多基于電鏡觀察的結果,且主要集中于納米顆粒和納米顆粒團聚體.對于納米金屬顆粒的有機結合態或與金屬離子共存情況下,它們各自在植物體內的運輸途徑和機制并未開展系統研究.

圖2 金屬納米顆粒在植物體內的運輸途徑及其在韌皮部胞間連絲內的積累[32]Fig.2 Transport routes of metal nanoparticles in plant and accumulation of gold nanoparticles in the plasmodesma of the phloem complex in root cells[32]
一般認為,植物對金屬離子的長距離運輸(土壤?根?莖?葉)是通過木質部完成的,如氧化銅納米顆粒在南瓜中會通過木質部運輸[35].但有研究發現,對于一些低濃度重金屬離子在水培植物中的運輸,無法用單一的木質部運輸理論進行合理解釋,可能存在韌皮部運輸現象[36].某些重金屬元素(如Cd)在植物葉片中的累積過程表現出明顯的韌皮部運輸特征.Zhang 等[37]發現,CeO2NPs 以Ce(Ⅱ)和Ce(Ⅲ)的混合物形式通過木質部從根向地上部運輸,隨后以CeO2的形式通過韌皮部從地上部返回根.因此,納米金屬顆粒在植物體內的運輸是由木質部與韌皮部共同構成的維管系統實現的,木質部中流動方向從下到上,韌皮部中流動方向從上到下,整個流動過程是非循環的,納米金屬顆粒在木質部與韌皮部之間的運輸途徑與機制尚有待深入研究.
3 納米金屬顆粒在植物中的生物轉化
納米金屬顆粒在植物體內的生物轉化是其形態轉化的另一重要方式.首先,納米金屬顆粒在根際可能發生形態變化,然后被吸收至植物體內.有研究表明,植物根際分泌物的成分決定了氧化鈰納米顆粒的形態轉化以及被植物吸收后的轉運機制,但根系分泌物成分和氧化鈰納米顆粒發生形態轉化后的轉運機制需要進一步研究.其次,納米金屬顆粒的轉化可能發生在植物體內.納米金屬顆粒可以與植物體內的蛋白發生絡合,生成有機結合態的納米金屬/納米金屬氧化物(如巰基化合物)[38];植物體內的有機酸可以促進納米金屬顆粒的解離;植物體內的還原性物質(光合作用中一些還原性酸、鐵氧還原蛋白、葡萄糖和果糖等)可以還原納米金屬顆粒解離出的金屬離子[39].Hernandez-Viezcas 等[40]發現,納米氧化鋅顆粒在沙漠植物天鵝絨豆中全部轉化為二價鋅離子;Wang 等[39]通過分根試驗發現,納米氧化銅顆粒從玉米葉或莖中通過韌皮部運輸重新回到根上,在此過程中二價銅被還原為一價的氧化亞銅和硫化亞銅.研究[37]發現,納米氧化鈰顆粒(CeO2NPs)在黃瓜根系表皮和細胞間隙中被還原為三價鈰,并與磷酸根形成針狀的磷酸鈰沉淀.另外,有研究表明,植物會從外界吸收金屬離子,在體內將它們轉化為納米金屬顆粒.例如,培養基中減少的Au(Ⅲ)、Ag(Ⅰ)、Pt(Ⅱ)在紫花苜蓿幼苗中轉化形成了相應的納米金屬顆粒[41].盡管相關研究報道納米金屬顆粒在植物體內存在生物轉化,但到目前為止納米金屬顆粒在植物體內的轉化機理尚不清楚.
4 納米金屬顆粒對植物的生物效應
納米金屬顆粒對植物的生物效應機制復雜,可能造成植物細胞膜損傷、細胞器功能破壞、DNA 損傷等負面影響(見圖3).在這些過程中,細胞內ROS 水平起到了重要作用.納米金屬顆粒可以促進植物體內ROS(單線態氧、超氧陰離子、雙氧水和羥基自由基)的產生[25].100 mg/L 的納米金可以將薺菜體內的ROS 含量提高29%[42].納米銀和納米氧化鋅也可以在植物體內產生大量的雙氧水[43].Panda 等[43]研究發現,納米金不僅顯著增加了植物體內的雙氧水含量,同時還顯著增加了超氧陰離子的含量.ROS 的生成可以抑制葉綠素的合成[44],但也有研究發現納米金屬材料(如納米氧化鈰、納米氧化鋅、納米金、納米二氧化鈦)不會影響甚至會促進葉綠素的合成[42,45].ROS 還會引起植物細胞的脂質過氧化,進而導致離子滲漏和細胞死亡[25].研究[44,46]發現,納米氧化銅、納米氧化鋅和納米氧化鈰可以造成小麥和玉米的脂質過氧化;但Rico 等[47]發現,納米氧化鈰對水稻沒有明顯的脂質過氧化.此外,ROS 還會影響線粒體膜電位及抗氧化酶活性,這都可能對植物細胞造成損傷.由此可見,ROS 的產生是納米金屬顆粒對植物產生毒性效應的主要原因之一,但其機制還需要更深入的研究.除此之外,納米金屬顆粒還可以影響植物對營養元素的吸收和運輸.有學者認為,納米金屬顆粒對植物吸收和運輸營養元素的抑制比ROS 引起的毒害作用更強[25].納米氧化鈰可顯著降低大豆根瘤的固氮活性,進而抑制大豆的生長[48].納米氧化鈰可以與磷結合,從而顯著降低植物體內有效磷的含量[49].相反,納米二氧化鈦顯著增加了黃瓜體內有效磷和鉀的含量.Taylor 等[50]研究發現,納米金下調了擬南芥中重金屬轉運調控基因的表達.納米金屬顆粒對植物營養元素吸收和運輸的影響尚無一致結論,這可能由納米材料理化性質、植物種類以及培養條件等因素共同決定.
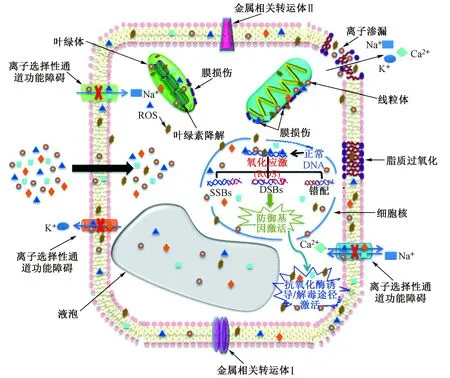
圖3 金屬納米顆粒對植物的生物效應[32]Fig.3 Biological effects of metal nanoparticles on plants[32]
5 結論與展望
5.1 結論
a) 納米金屬顆粒在土壤中遷移轉化過程中的形態轉化受到土壤環境(pH、離子強度、離子價態、溫度、DOM)的影響,會發生吸附/解吸、分散/沉降、解離和氧化/還原等過程.
b) 納米金屬顆粒在植物中的運輸過程:首先吸附在植物的根部,再通過質外體或共質體途經向植物內部轉移,由木質部和韌皮部組成的維管系統進行轉運.
c) 納米金屬顆粒在植物中的生物轉化過程主要包括在根際的轉化與在植物體內的轉化.根際分泌物以及植物體內的蛋白質與有機酸等都起到了至關重要的作用.
d) 納米金屬顆粒在植物中的生物效應主要包括植物氧化應激以及對植物吸收營養的抑制.
5.2 展望
a) 由于納米材料種類繁多,性質各異,納米金屬材料在植物-土壤系統中的遷移轉化過程及相關機制尚未明確.未來可從以下幾方面繼續深入研究:①納米金屬顆粒在土壤中形態轉變過程的耦合效應,各賦存形態在植物體內的運輸途徑和機制還有待確認;②在植物生長不同階段,納米金屬顆粒在植物體內各部位發生的形態轉化也亟需研究;③關于納米金屬顆粒對植物毒性的研究仍有許多問題有待解決,如各形態對應的植物毒性效應、毒性效應與植物對各形態的吸收運輸之間的聯系等.這些問題的研究可為今后如何調控納米金屬顆粒在植物可食部分的積累以及阻斷植物對納米金屬顆粒的吸收途徑等提供理論支持.
b) 試驗技術的限制使得目前無法采取有效措施以阻止土壤中納米金屬顆粒通過農產品進入食物鏈,也阻礙了生物修復技術的發展.基于同步輻射的分析技術在納米金屬顆粒的環境轉化機制研究中可以發揮不可替代的作用.同步輻射X-射線吸收(XAS)、微束X 射線吸收近邊結構分析(μ-XANES)及同步輻射掃描透射X-射線顯微成像技術(STXM)等技術是研究亞微米尺度下的結構與功能的有力工具.隨著同步輻射技術的發展,基于同步輻射的成像、形態和微區元素分布分析技術能夠為研究納米金屬在植物體內的分布、形態,揭示納米金屬顆粒在土壤-植物系統中各微界面上的遷移轉化規律及機制,明晰各形態納米金屬顆粒對植物的毒性效應提供關鍵性的試驗證據.
c) 納米金屬材料在植物-土壤系統中的遷移轉化及對植物的生物效應是納米金屬顆粒安全性研究的重要內容.今后應通過同步輻射的分析技術結合膠體化學、土壤化學、生物化學和分析化學等手段,認識納米金屬顆粒在土壤中各形態轉變過程的耦合效應,了解納米金屬顆粒在植物體內的生物轉化過程和機制,厘清納米金屬顆粒各賦存形態在植物體內的運輸途徑和機制,闡明納米金屬顆粒各形態對植物的毒性效應,進而為發展植物納米金屬顆粒的調控及阻斷技術,降低納米金屬顆粒的生物毒性,設計和開發環境友好型、生物相容性納米金屬材料提供科學依據.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
