不同年限毛竹-白及復合系統(tǒng)土壤微生物群落多樣性特點
2022-03-05 06:12:00曹小青孫孟瑤王晶晶李培璽儲炳銀徐小牛
中國土壤與肥料 2022年1期
關鍵詞:利用
曹小青,王 亮,孫孟瑤,王晶晶,李培璽,儲炳銀,徐小牛
(安徽農業(yè)大學林學與園林學院,安徽 合肥 230036)
森林土壤微生物是土壤生態(tài)系統(tǒng)中最活躍的組分之一,在促進土壤養(yǎng)分循環(huán)、物質轉化和能量交換中起著重要作用[1],具有分解土壤礦物質、促進腐殖質的分解和合成、保護植物根系免受病原菌的侵染和其他特性的能力[2]。同時,微生物本身還含有一定的N、P、K等,為地上植物生長提供必要的養(yǎng)分,直接或間接影響著植物的生長[3]。土壤微生物不僅受地上植物和土壤特性的影響,還通過自身特性與植物相互作用,形成與植物的交互反饋機制[4]。大量研究表明,土壤微生物群落功能多樣性不僅能敏感地指示氣候和土壤環(huán)境狀況的變化,還可以反映土壤質量和森林健康狀況[5-6],被認為是重要的生態(tài)指標。因此,研究森林土壤微生物群落多樣性可以揭示森林生態(tài)系統(tǒng)“植物-土壤-微生物”三者之間的相互作用機制[7]。
毛竹(Phyllostachys pubescens)是禾本科(Gramineae)常綠喬木狀單子葉植物。毛竹作為我國亞熱帶地區(qū)廣泛分布的大徑級竹種,栽培歷史悠久,其經營包括材用、筍用和筍材兼用,也可生態(tài)林經營,具有重要的經濟和生態(tài)價值,在脫貧致富、鄉(xiāng)村振興中發(fā)揮著重要作用。由于毛竹特殊的繁殖方式,長期以來多以單一經營為主,易受自然干擾及市場風險的沖擊,嚴重影響其經營效益[8]。同時,毛竹林結構單一,物種多樣性低,存在一定的生態(tài)風險。因此,為提高竹林的經營效益及其生態(tài)穩(wěn)定性,近年來竹木混交以及竹藥、竹草復合經營備受關注[9],已成為生態(tài)研究的熱點。相關研究表明,實施竹木混合經營有利于改善土壤養(yǎng)分狀況和提高森林生產力,有利于毛竹林的可持續(xù)發(fā)展和管理[10-11]。但是,關于毛竹與藥用植物復合經營的研究相對較少[9,12]。
白及(Bletilla striata)屬蘭科(Orchidaceae)多年生宿根草本、國家二級保護植物,其假鱗莖是我國傳統(tǒng)、珍稀名貴中藥材,具有收斂止血、消腫生肌的功能,主要用于治療咯血、吐血、外傷出血、瘡瘍腫毒、皮膚皸裂等[13]。白及種植3~4年后即可收獲假鱗莖,生長5年以上假鱗莖藥用價值更高。由于需求增加,白及市場價格不斷攀升,野生資源因亂采濫挖以及天然生境的破壞而瀕臨滅絕。因此,急需開發(fā)高效、綠色的白及林下仿野生栽培技術,促進白及中藥農業(yè)可持續(xù)發(fā)展[9,14]。
目前,有關林下套種白及的試驗研究已有一些報道[9,15],但是毛竹-白及復合經營的研究甚少。為此,基于長期定位試驗,對不同套種年限的毛竹-白及復合林分土壤微生物群落功能多樣性進行分析,旨在探討土壤微生物、植被與土壤質量之間的相互作用關系,為毛竹-白及復合模式的可持續(xù)經營提供依據。
1 材料與方法
1.1 樣地概況與樣品采集
試驗樣地位于安徽廣德市邱村鎮(zhèn)(119°23′37″E,31°01′49″N),屬皖南山地和沿江平原的過渡帶,以丘陵崗地為主,地勢較平坦,竹資源豐富。屬北亞熱帶濕潤季風氣候,四季分明,溫和濕潤,年平均氣溫為16.3℃,無霜期近300 d;多年平均降水量達1294 mm,主要分布在夏秋季節(jié)。地帶性植被為亞熱帶常綠闊葉林,并分布有大面積毛竹林、杉木(Cunninghamia lanceolata)、馬 尾 松(Pinus massoniana)和濕地松(P. elliottii)等人工林。地帶性土壤為紅黃壤,質地較黏,土層深厚,肥力一般。
該地自2010年開始毛竹林下種植中藥材,以白及、黃精為主,目前種植面積約150 hm2。套種前,通過疏伐降低竹林密度,使郁閉度在0.5~0.6,再進行林地墾復,深度約30 cm,種植前每公頃施用腐熟有機肥22.5 t。選用1年生白及組培苗,按株行距20 cm×30 cm栽植,栽后每年除草2~3次,沒有追肥和使用農藥。于2019年5月白及生長前期在試驗區(qū)內進行樣地調查與樣品采集,不同復合林分的栽培管理措施一致,其中毛竹林為天然林,除適度采伐竹材外無任何經營措施。選取立地條件相近、套種白及年限分別為1、3、4年的毛竹復合經營林分,分別設置面積為20 m×20 m的標準樣地各3個,樣地間距不小于100 m,采用“S”形5點混合取樣法,用土鉆(內徑3 cm)采取白及行間0~10 cm的土樣,過篩去除凋落物、細根和石礫等,混合均勻后分為兩份,一份裝入無菌袋,置于便攜式冰盒冷藏帶回實驗室,在4℃冰箱保存,用于土壤微生物功能多樣性的測定;另一份風干后過2 mm篩,用于土壤理化性質測定。
1.2 土壤理化性質和土壤微生物群落功能多樣性的測定
土壤理化性質指標采用常規(guī)方法測定[16]。土壤含水量采用烘干法測定;土壤pH按照水浸提電位法(水土比為2.5∶1),采用Extech Ⅱ型pH計測定;土壤有機碳和全氮使用元素分析儀(EA3000,Vector,Italy)測定;銨態(tài)氮、硝態(tài)氮使用2.0 mol·L-1KCl浸提、全磷經濕式消煮,隨后采用自動流動分析儀(Alliance-Futura,France)測定。
土壤微生物群落功能多樣性采用31種碳源的Biolog-ECO微平板法測定[17-18],即稱約10 g新鮮土樣放入250 mL的三角瓶中,加入100 mL已滅菌的0.85% NaCl溶液,搖床振蕩30 min(轉速180 r·min-1),取上清液稀釋1000倍,再置于4℃冰箱內靜置2~3 min,取其上清液倒入無菌培養(yǎng)皿,利用移液槍向ECO微平板每孔內加入150 μL稀釋液,接種好的Biolog-ECO板于28℃生化培養(yǎng)箱中黑暗培養(yǎng)7 d,每隔24 h在Biolog微平板自動讀取儀上測其吸光度。Biolog-ECO板上的31種碳源依其化學基團的性質,分成6大類,即糖類(10種)、羧酸類(7種)、氨基酸類(6種)、聚合物類(4種)、多胺類(2種)、多酚酸類(2種)。
1.3 數據處理與統(tǒng)計分析
通過Biolog-ECO微平板測定的平均每孔顏色變化率(Average well color development,AWCD)可表示微生物群落的整體代謝活性[18],是反映土壤微生物碳源利用能力及其活性和多樣性大小的一個重要指標:
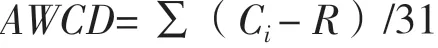
式中: Ci為第i個孔590 和750 nm的吸光度值,R為對照孔吸光度值,若Ci- R ≤ 0,其吸光度值計為0;31為ECO板供試碳源的種類數。
根據培養(yǎng)72 h的AWCD值變化,土壤微生物群落多樣性指數按下式計算[18]:

式中:Pi=(Ci-R)/∑(Ci-R),表示第i孔的相對光密度值與整個微板相對光密度值總和的比值,ni表示第i孔的相對吸光度(Ci-R)。
利用Excel 2016整理數據,SPSS 19.0進行統(tǒng)計分析,采用單因素方差分析和LSD多重比較法對不同套種年限毛竹-白及復合林分土壤微生物碳源利用特征的差異顯著性進行分析(P<0.05),Pearson相關系數檢驗各因子之間的相關性;采用Origin 2018 64bit繪圖。
2 結果與分析
2.1 不同套種年限毛竹-白及復合林分的土壤理化性質
不同套種年限對復合林分土壤理化性質具有顯著影響(表1)。套種4年的林分土壤含水量、全氮、全磷含量均顯著高于套種1和3年的復合林分(P<0.05);套種3年的土壤有機碳、銨態(tài)氮顯著低于套種1和4年的復合林分(P<0.05);套種1年的土壤pH顯著低于套種3和4年的復合林分(P<0.01)。
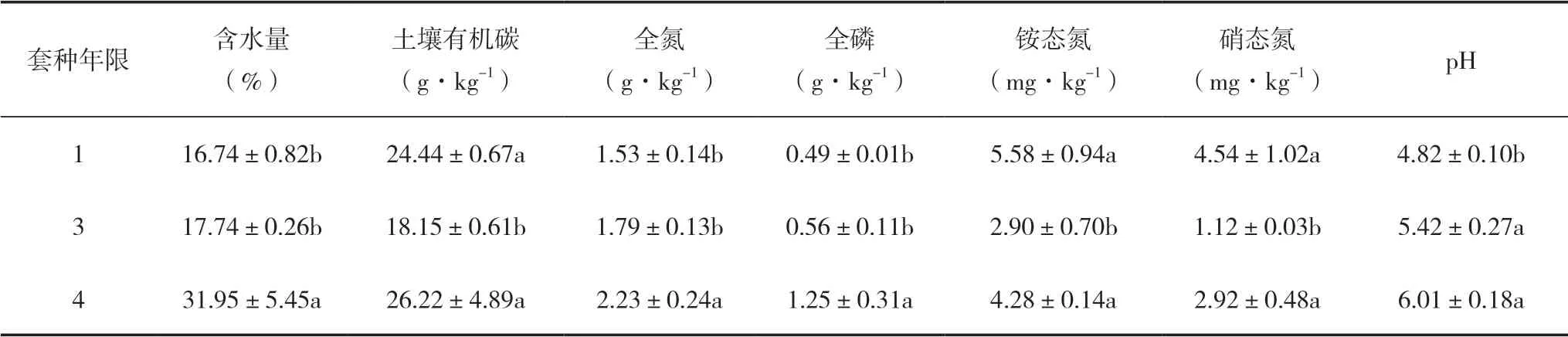
表1 毛竹-白及復合系統(tǒng)土壤理化性質
2.2 不同套種年限毛竹-白及復合林分土壤微生物群落碳源利用特征
試驗結果顯示,不同林分土壤微生物群落AWCD值隨著培養(yǎng)時間的增加呈逐漸升高的趨勢(圖1)。在培養(yǎng)0~24 h內沒有顯著變化,隨后呈直線上升,表明微生物代謝活性旺盛,碳源被大量利用。不同套種年限的土壤微生物利用不同碳源的AWCD值呈現4年>3年>1年的變化趨勢。與套種1年的林分相比,套種3和4年的AWCD值均有顯著差異(P<0.01);而套種1年的AWCD值無顯著差異(P>0.05)。

圖1 不同套種年限毛竹-白及復合林分土壤微生物群落平均顏色變化率
土壤微生物群落利用6大類碳源的AWCD值均隨著培養(yǎng)時間的延長逐漸升高(圖2),對6大類碳源的代謝能力差異顯著(P<0.01)。在不同碳源利用水平上,不同套種年限林分差異較大(表2)。套種1年的土壤微生物群落碳源利用水平從高到低依次是羧酸類、氨基酸類、聚合物類、糖類、多胺類、多酚酸類,除糖類之外,均顯著低于套種3和4年林分(P<0.05)。套種3和4年林分土壤微生物群落對碳源利用僅在多胺類存在顯著差異(P<0.05),在其他碳源利用上兩林分間無顯著差異。

表2 培養(yǎng)72 h土壤微生物對碳源的利用程度
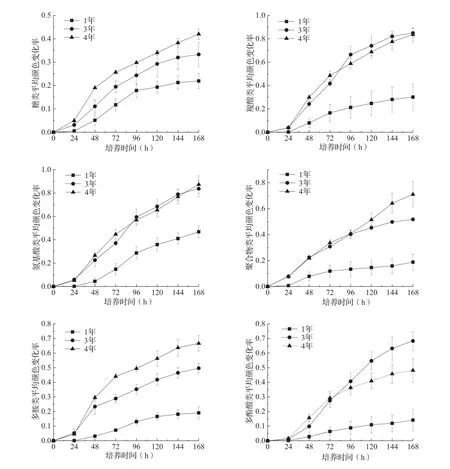
圖2 不同套種年限毛竹-白及復合林分土壤微生物利用不同碳源的AWCD值
2.3 不同套種年限毛竹-白及復合林分的土壤微生物群落功能多樣性
隨著套種年限的增加,土壤微生物多樣性、豐富度和優(yōu)勢度指數均表現為逐漸升高的趨勢(表3)。不同套種年限的多樣性指數和72 h AWCD值無顯著差異(P>0.05);套種3和4年的土壤微生物豐富度和優(yōu)勢度指數顯著高于套種1年林分(P<0.01)。可見,套種年限對土壤微生物群落物種豐富度和均勻度指數影響較大,但對微生物群落優(yōu)勢度指數影響較小。

表3 不同套種年限毛竹-白及復合模式土壤微生物群落多樣性指數
主成分分析(PCA)結果顯示,第一主成分(PC1)貢獻率為52.39%,第二主成分(PC2)貢獻率為14.94%,累計貢獻率達67.33%。PC1和PC2作為微生物群落碳源利用變化的主要成分,可反映不同套種年限土壤微生物群落代謝能力的基本情況。不同套種年限的復合林分土壤微生物群落具有明顯的分異(圖3)。套種1年林分主要分布在PC1軸負方向,套種3年林分分布在PC2軸正方向,套種4年林分分布在PC1軸正方向;且不同套種年限林分的土壤樣本分布較為分散,說明隨著套種年限的增加,土壤微生物群落代謝功能變化明顯。

圖3 不同套種年限毛竹-白及復合模式土壤微生物群落多樣性的主成分分析
由表4可知,與PC1具有較高相關性的碳源(| r | > 0.6)有18個,包括糖類3個、羧酸類6個、氨基酸類4個、聚合物類2個、多胺類2個和多酚酸類1個;與PC2具有較高相關性的碳源有5個,包括糖類2個、羧酸類1個和氨基酸類2個。表明6大類碳源對PC1和PC2均起分異作用,其中對PC1影響大。

表4 31種碳源在PC1和PC2上的載荷值(|r|>0.6)
2.4 土壤微生物多樣性和土壤理化性質的相關性
從表5可以看出,土壤微生物McIntosh指數與土壤全磷、pH呈顯著正相關(P<0.05);土壤微生物Shannon指數與土壤全磷呈顯著正相關(P<0.05),與pH值呈極顯著正相關(P<0.01);土壤微生物Simpson指數與土壤全磷呈顯著正相關(P<0.05),與銨態(tài)氮和硝態(tài)氮呈顯著負相關(P<0.05),與pH值呈極顯著正相關(P<0.01)。這說明土壤微生物群落多樣性指數與土壤理化性質密切相關、相互影響。

表5 土壤微生物群落多樣性指數與土壤理化性質的相關系數
從表6中可以看出,6大類碳源中的糖類和多酚酸類與pH呈顯著正相關(P<0.05);羧酸類和聚合物類與全磷呈顯著正相關(P<0.05),與pH呈極顯著正相關(P<0.01);氨基酸類與pH呈極顯著正相關(P<0.01);多胺類與含水量呈顯著正相關(P<0.05),與土壤有機碳、全氮、全磷以及pH均呈極顯著正相關(P<0.01)。這說明土壤微生物群落與土壤理化性質關系緊密,尤其是多胺類與理化性質之間的關系顯著。
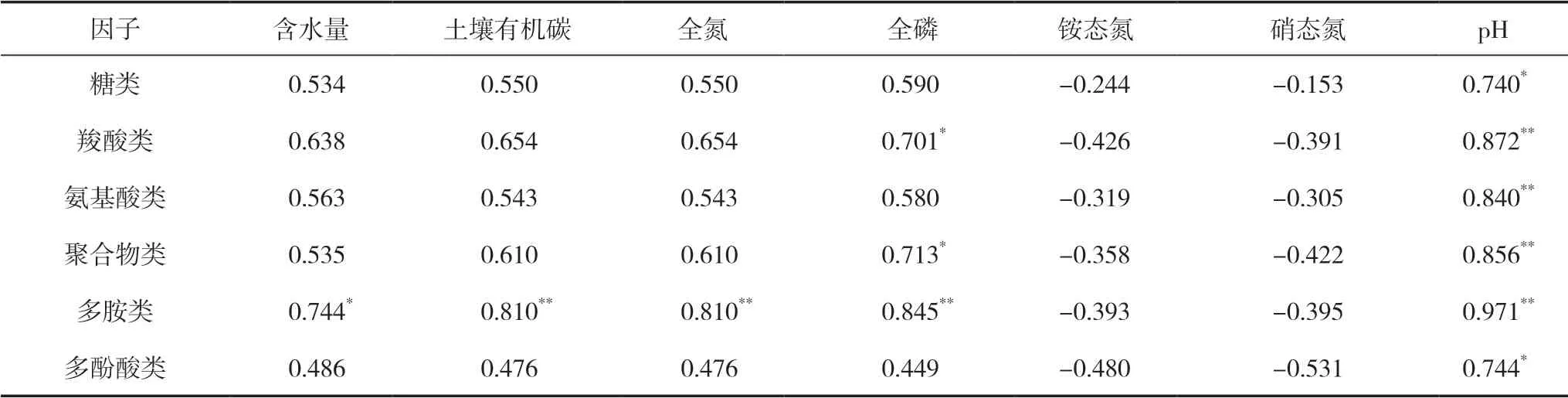
表6 土壤微生物碳源利用強度與土壤理化性質的相關系數
3 討論與結論
3.1 套種年限對土壤微生物群落碳源利用特征和功能多樣性的影響
土壤環(huán)境是土壤微生物組成和差異的主要因素[19],而森林凋落物、根系分泌物和土壤有機質是土壤微生物可利用碳源的主要來源[20-22]。有研究表明通過合理的間作套種以及不同樹種混交,可提高土地利用率,改善土壤環(huán)境,增加土壤微生物多樣性[23]。本研究發(fā)現,不同套種年限的毛竹-白及復合林分土壤微生物群落AWCD值變化存在明顯差異,隨著套種年限增加,土壤微生物碳源利用強度顯著提高,套種4年林分土壤微生物代謝活性最強,而套種1年林分土壤相對較弱。這可能與白及生長發(fā)育有關,套種初期(1年)白及個體小,覆蓋度低,隨著年限增加,白及覆蓋度不斷提高,套種4年時達85%以上,土壤生態(tài)系統(tǒng)逐漸趨于穩(wěn)定,為微生物群落發(fā)育提供了適宜條件;同時,白及地上部生物量隨生長年限增加逐漸增大,凋落物量以及根系分泌物增多,為微生物群落提供了充裕的可利用碳源種類和數量,促進了微生物的生長發(fā)育[24-25]。因此,微生物群落多樣性顯著提高,代謝功能增強。王超群等[26]研究顯示,隨著林齡的增加,杉木林土壤微生物群落碳源利用率逐漸提高,這與本研究結果一致。
多樣性指數反映了土壤微生物群落功能多樣性變化,不同指數反映多樣性不同側面,Shannon指數和McIntosh指數主要反映群落物種個體數和分布均勻度,而Simpson指數反映群落中最常見類群的優(yōu)勢度[27-28]。本研究中Shannon指數、McIntosh指數和AWCD值均隨套種年限增加顯著提高,表明土壤微生物功能多樣性隨年限增加而提高。但Simpson指數在不同套種年限之間差異不顯著,說明土壤微生物群落的優(yōu)勢種群并沒有隨套種年限增加而顯著變化。由于土壤微生物群落結構及其多樣性不僅受到立地環(huán)境的影響,而且與植被類型及其組成結構緊密相關[29],在立地環(huán)境基本一致的條件下,則主要受到植物群落結構的影響。Meril?等[30]研究得出,土壤微生物群落多樣性與林下物種豐富度及多樣性呈正相關關系。由于本研究的試驗地立地條件較一致,因此,不同套種年限林分結構變化可能是導致土壤微生物群落功能多樣性差異的主要因素。
3.2 套種年限對土壤理化性質的影響
根據不同類型碳源利用微生物隨套種年限變化的差異(圖2)可知,糖類碳源利用微生物受套種年限影響程度比較小,可能是毛竹-白及復合林分土壤的優(yōu)勢微生物類群。但是,隨著套種年限增加,在套種3年后氨基酸類、羧酸類、糖類、聚合物類等碳源利用微生物成為優(yōu)勢類群。主成分分析結果直觀反映了不同套種年限毛竹-白及復合林分土壤微生物群落利用碳源的種類和強度的整體差異[31]。套種年限的變化使得植被覆蓋度發(fā)生變化,由此引發(fā)土壤環(huán)境改變,從而導致土壤微生物群落結構變化。本研究結果表明,表層土壤中有機碳、全磷、pH隨著復合年限增加逐漸增加,與Chatterjee等[32]研究得出的土壤pH隨著林齡的增加而降低的結果不一致。這可能是由于復合林分中白及生長逐年加快,對土壤養(yǎng)分需求較大,導致土壤中氮素的轉化加速,有效氮增加,根系在吸收硝態(tài)氮時分泌出OH-來維系細胞膜內外電荷平衡,最終通過土壤中的OH-來協(xié)調土壤的酸堿度[33]。同時,白及生長加快,地上部生物量逐年增加,大量吸收土壤下層養(yǎng)分,地上部枯死后歸還土壤增多,在一定程度上加速了土壤養(yǎng)分循環(huán)速率。相關分析結果表明,土壤微生物多樣性指數與土壤全磷、pH、銨態(tài)氮和硝態(tài)氮具有顯著相關性,土壤微生物碳源利用強度與土壤pH也具有顯著相關性,與王淼等[34]研究結果相似。可見,土壤微生物碳源利用特征及多樣性指數受到諸多因素的影響,加之Biolog-ECO微平板法的局限性[18],如要全面認識植物-土壤-微生物之間的互作機制,今后須結合高通量測序等技術,深入探討竹藥復合模式土壤微生物群落多樣性的演變規(guī)律。
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
中學生數理化(高中版.高考數學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業(yè)設計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
