無種質基因型限制結縷草組培快繁體系研究
2022-03-11 21:10:40許建民仇學文劉艷梁慧敏
江蘇農業科學 2022年4期
許建民 仇學文 劉艷 梁慧敏






摘要:結縷草從愈傷組織發育成胚性愈傷再進一步分化為完整植株一直是該領域的研究重點與難點。本研究通過試驗建立起一套完善的無種質基因型限制的結縷草組培快繁體系。結果表明,愈傷組織誘導培養基為MS+2 mg/L 2,4-D+0.2 mg/L 6-BA+40 g/L蔗糖最為合適,體胚細胞誘導培養基為改良MS+2 mg/L 2,4-D+0.2 mg/L NAA+0.1 mg/L 6-BA+40 g/L蔗糖最為合適,胚狀體增殖與生長培養基為MS+0.1~0.2 mg/L 2,4-D+1 mg/L NAA+01 mg/L 6-BA+30 g/L蔗糖最為合適,成苗快繁培養基為MS+0.02~0.05 mg/L NAA+1~2 mg/L 6-BA+30 g/L蔗糖最為合適,生根培養基MS最為合適。
關鍵詞:種質;基因型;結縷草;組培;快繁體系
中圖分類號:S317 文獻標志碼: A
文章編號:1002-1302(2022)04-0036-07
收稿日期:2021-05-11
基金項目:江蘇農林職業技術學院重點科技創新類項目(編號:2017kj09);江蘇高校“青藍工程”項目(編號:2019)。
作者簡介:許建民(1981—),男,甘肅張掖人,碩士,副教授,主要從事設施園藝環境控制研究。Tel:(0511)87290588;E-mail:jsnlxjm@vip.163.com。
通信作者:梁慧敏,博士,教授,主要從事林草生物技術育種研究。Tel:(0511)87290588;E-mail:278151187@qq.com。
結縷草屬(Zoysia)植物主要分布于亞洲東部廣大區域內,在我國主要分布于從東北遼寧到南方廣西等地的沿海狹長地帶。由于結縷草生態環境復雜多樣,具有典型嚴格的異花授粉特性,導致結縷草種間雜交及天然雜種大量存在,種內變異很大,也使得天然野生種群個體間保持了豐富的自然變異、基因型差異及遺傳多樣性。
利用生物技術結合常規育種方法進行結縷草新品種的選培育,是提高結縷草育種效率的有效途徑。采用生物技術進行品種或種質基因型改良及種質快繁最有效的途徑就是建立高頻體細胞胚再生技術體系,因為體細胞胚是單細胞起源的,體細胞胚再生途徑不但重演了合子胚形態發生的過程,實現了再生種子的來源,也是種質保存、誘導體細胞變異或基因轉化的理想受體。體細胞胚快繁不僅能保證優良種群個體基因遺傳一致性和穩定性,還能保持新種質的特異性。所以,體細胞胚再生技術是植物組培克隆中最具吸引力的研究力向。
結縷草是禾本科中較難培養的物種,大部分研究選擇的材料是以1~3個種或種質基因型為主,牽涉的種質基因型數量較少,且以多細胞的器官發生途徑為主[1-4],從愈傷組織變成胚性愈傷組織及再生植株都較困難,組培再生率低,再生過程長。有研究提出,結縷草體細胞胚的產量和質量依賴于對培養基組成和植物激素等條件的優化[5],但這方面的研究較少。因此,建立結縷草無種質基因型限制的高頻體胚再生培養技術是實現結縷草種質改良與高效利用的關鍵,具有重要的現實意義。
1 材料與方法
1.1 試驗材料
試驗于2018—2020年間在江蘇農林職業技術學院江蘇現代園藝工程技術中心開展,以分別來源于遼寧、青島半島、江蘇及長三角周邊地區、廣東及周邊地區的野生或育成的結縷草5個種8個種群的種質基因型為材料,包括結縷草[Zoysia japonica Steud.(Z1)]、中華結縷草[Zoysia sinica Hance(Zz)]、溝葉結縷草[Zoysia matrella (L.) Merr(Zg)]、大穗結縷草[Zoysia macrostachya Franch.et Saw.(Zd)]、細葉結縷草[Zoysia tenuifolia Willd. ex Trin.(Zx)]、青島結縷草[Zoysia japonica cv. Qingdao(Zq)]、蘭引Ⅲ號結縷草[Zoysia japonica cv. Lanyin3(Z3)]、華南半細葉結縷草[Zoysia matrella cv. Huanan(Zh)]等,分別用符號Z1、Zz、Zg、Zd、Zx、Zq、Z3、Zh表示。
1.2 試驗方法
1.2.1 外植體處理及無菌苗獲取
切取生長期的8種結縷草半嫩莖帶2~4節匍匐莖段,先用洗潔精水沖洗10 min左右,再用3%次氯酸鈉溶液消毒 15 min,在無菌室用75%乙醇消毒30 s,無菌水沖洗4次,0.1%氯化汞消毒7 min,無菌水沖洗7次,滅菌吸水紙吸干水分,剪取含2~3個節外植體接種到無菌芽苗培養基上進行無菌芽苗的萌動生長馴化培養與預篩選(培養基配方:1/4 MS+0.1 mg/L GA3+0.2 mg/L 6-BA+20 g/L蔗糖),每瓶接種5棵苗,共接種200瓶,在1 000~1 500 lx光照下 16 h/d,培養26~28 d。培養溫度為白天室溫(28±2) ℃,夜晚(20±2) ℃。
1.2.2 愈傷組織的獲取
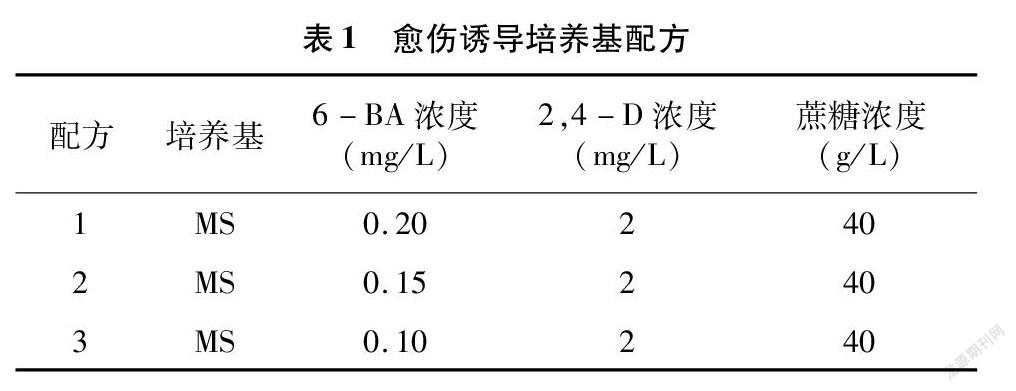
每種結縷草選擇開始萌動生長但未污染的芽苗原基600個,接種到3種不同配方的愈傷誘導培養基上,培養基配方如表1,每個處理每培養皿接種40個外植體,試驗重復5次,培養溫度同上,暗培養28~30 d后統計胚性愈傷組織的誘導率。
愈傷組織誘導率=產生愈傷的外植體數量接種外植體總數×100%。
1.2.3 體胚細胞的誘導
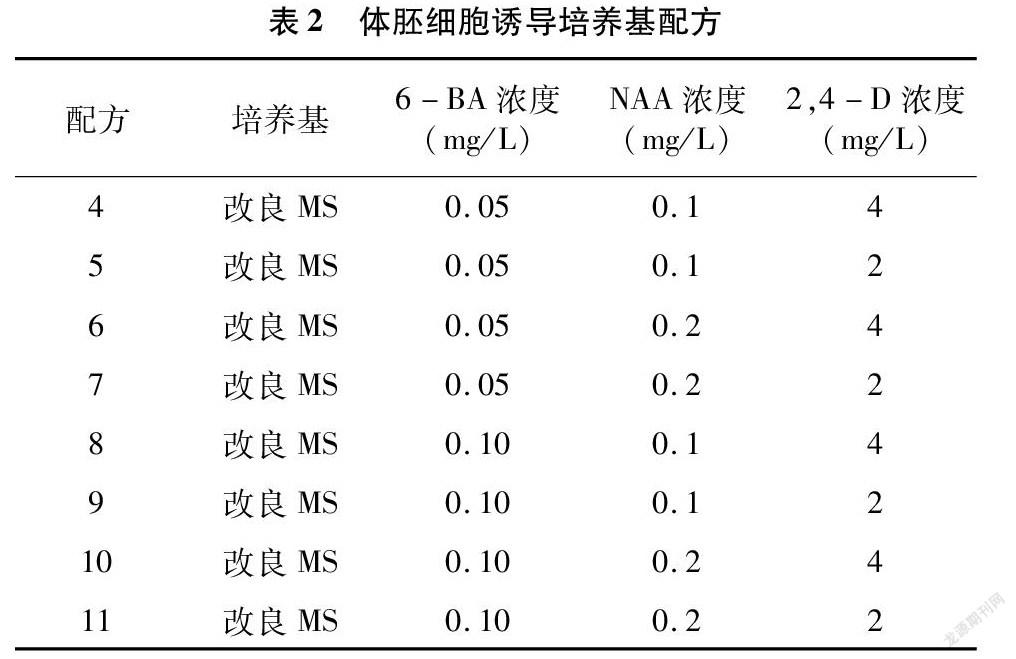
選擇愈傷分化最好的配方處理下的愈傷組織,繼續培養。培養基采用改良MS(用乙二胺四乙酸鐵鈉鹽替代硫酸亞鐵和乙二胺四乙酸二鈉,微量營養元素配方采用SH配方的濃度,有機試劑采用B5配方的濃度)+2,4-D+NAA+6-BA+40 g/L蔗糖+0.7%瓊脂的完全組合(表2)。每個處理每瓶接種12個愈傷,重復4次,暗培養下發育14 d,培養溫度同上,觀察胚狀體發育情況。
胚狀體發生率=發育成胚狀體的愈傷數愈傷組織總數×100%。
1.2.4 胚狀體的增殖與生長
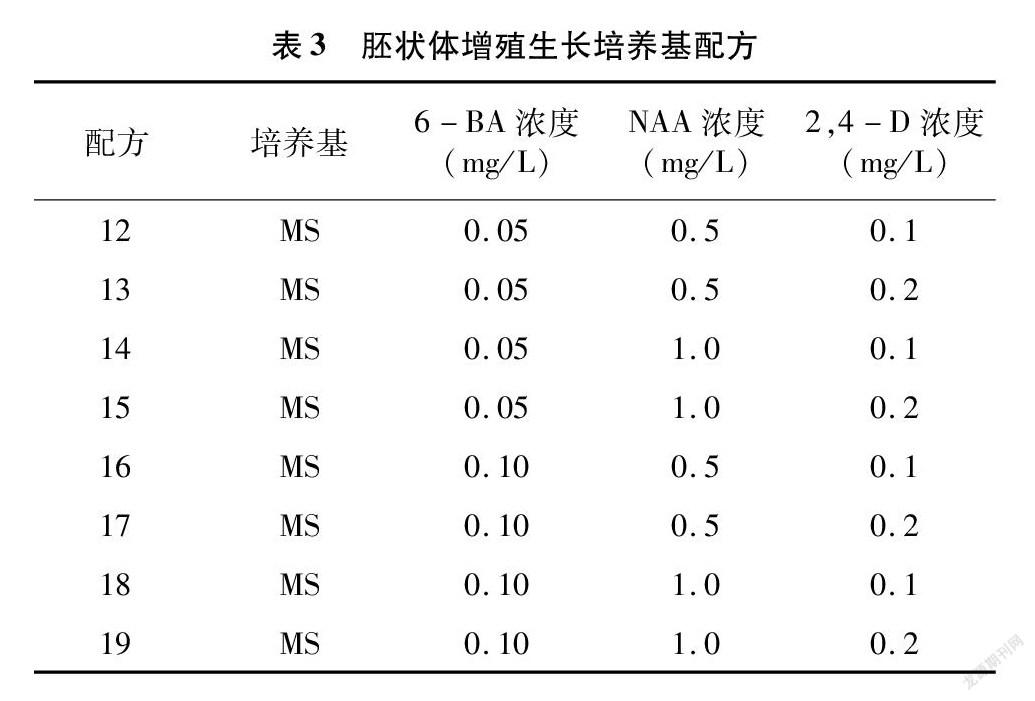
選取發育狀態良好、生長至魚雷期的胚狀體繼續進行增殖與生長培養。培養基采用MS+(0.1、0.2 mg/L)2,4-D+(0.5、1.0 mg/L)NAA+(0.05、0.10 mg/L)6-BA+30 g/L 蔗糖+0.7%瓊脂的完全組合(表3)。每瓶
接種9個胚狀體組織,重復4次,在1 000~1 500 lx光照下16 h/d,培養26~28 d。培養溫度同上。45 d 后觀察胚狀體的生長分化狀態,統計叢芽萌發率。
叢芽萌發率=萌發的叢芽數量接種的胚狀組織總數×100%。
1.2.5 結縷草的成苗快繁培養
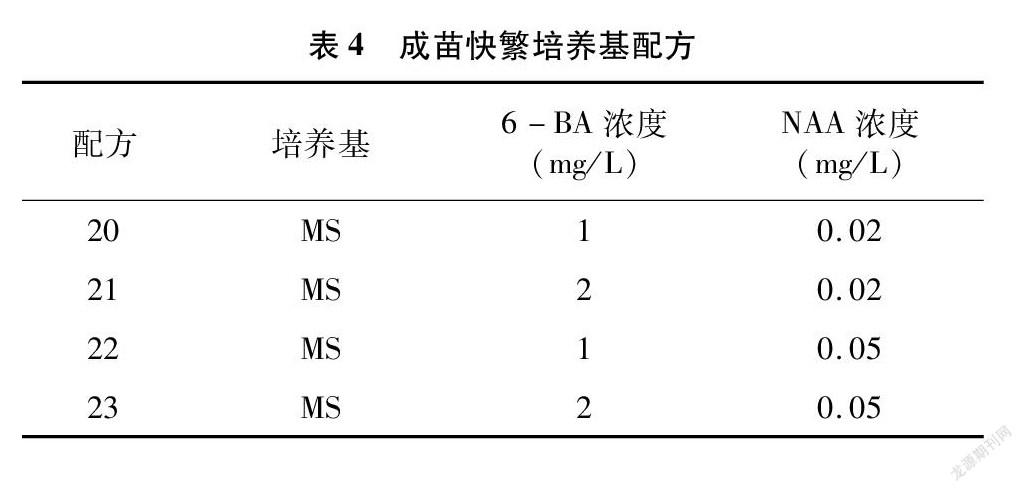
將分化好、生長狀態優的結縷草叢生芽接種到成苗快繁培養基上,進行繼代培養。培養基采用MS+(0.02、0.05 mg/L)NAA+(1、2 mg/L)6-BA+0 g/L蔗糖+0.7%瓊脂的完全組合(表4)。每個處理下每瓶接種6個叢生芽,重復4次,在1 000~1 500 lx光照下16 h/d,培養26~28 d。培養溫度同上。30 d后進行繼代培養,并統計不同處理下的叢生芽增殖系數。
叢生芽增殖系數=增殖芽數/接種芽數×100%。
1.2.6 叢生芽的壯苗生根培養
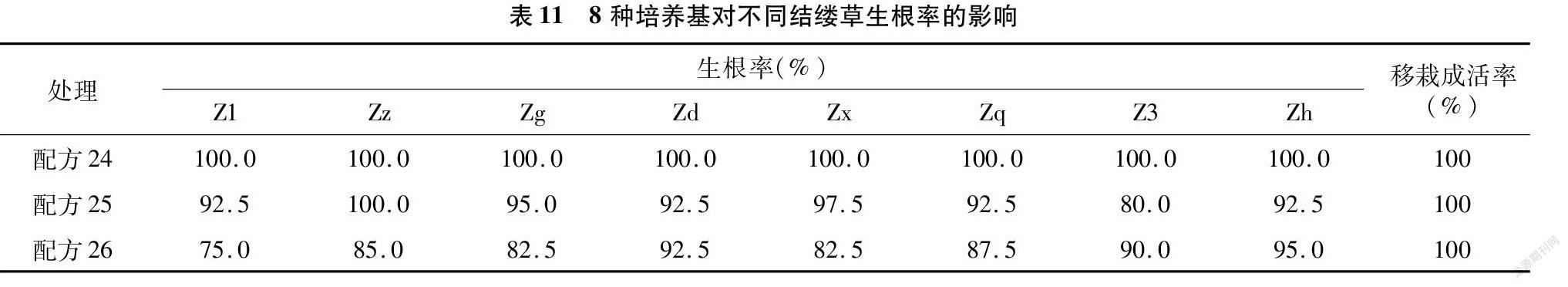
將快繁后獲得的結縷草成苗轉接到壯苗生根培養基上進行生根培養。培養基采用1/2 MS+(0、0.05、0.10 mg/L)IBA+20 g/L蔗糖+0.7%瓊脂的完全組合(表5)。每個處理下每瓶接種10個結縷草叢生芽,重復4次,2 000~2 500 lx光照強度下培養26~30 d,培養溫度同上,統計再生植株的生根率。
生根率=生根的叢生芽數接種數×100%。
1.2.7 再生植株的移栽
將壯苗生根培養后得到的生根的再生植株移栽到溫室小花盆中,盆土組分配比為蛭石 ∶泥炭土 ∶園土=2 ∶4 ∶4;移栽后遮陰處理,初期每天澆水1~2次,成活后逐漸減少澆水次數,15~20 d后統計成活率。
成活率=移栽后的成活數移栽總數×100%。
2 結果與分析
2.1 愈傷組織的誘導
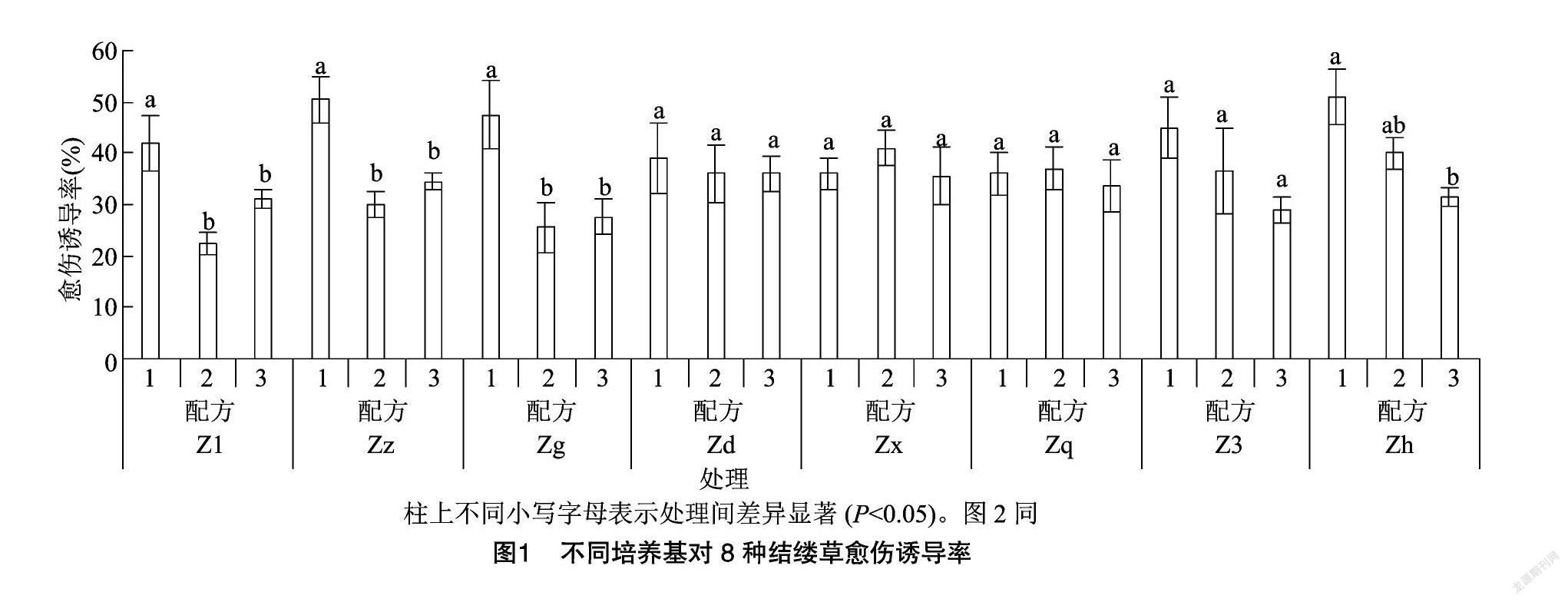
如圖1所示,在不同配方處理下的8種結縷草愈傷組織誘導率不盡相同。Z1、Zz、Zg、Zh這4種結縷草在不同配方處理后,愈傷誘導率均有顯著性差異,且都以配方1處理下產生的愈傷居多。而Zd、Zx、Zq、Z3這4種結縷草在不同的培養基配方處理后,愈傷誘導率未出現顯著性差異。從圖1也可以看出,除Zx與Zq是在配方2處理下的愈傷誘導率最高以外,其余的均是以配方1處理下的愈傷誘導率最高。
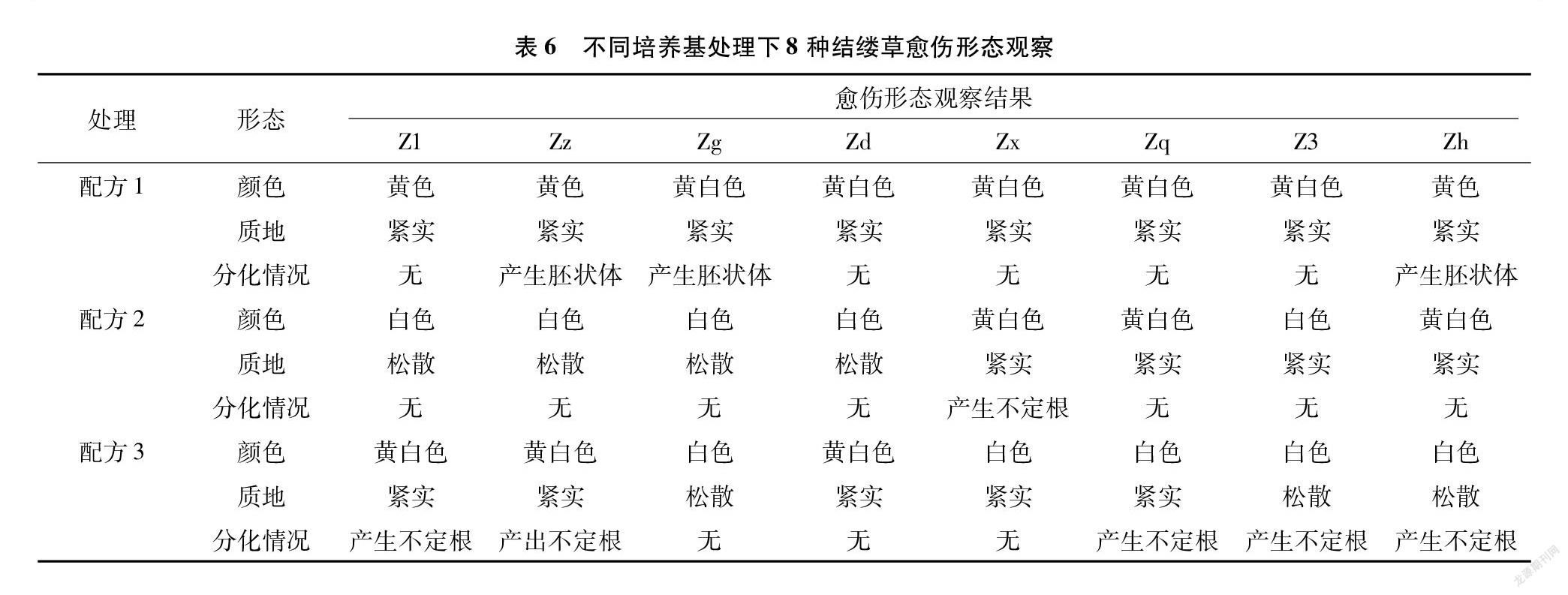
在對外植體進行脫分化培養時,產生的愈傷組織顏色與質地明顯存在差異且部分外植體出現了再分化的現象。本試驗中規定若超過2/5的外植體出現再分化的現象,則認為該培養基可導致植物再分化。從表6中可以發現,配方1處理下的8種結縷草外植體形成的愈傷組織顏色均為黃色或黃白色,質地緊實,Zz、Zg、Zh這3種結縷草則出現了球形胚、魚雷胚等典型的胚狀體組織,這表明,配方1在一定程度上可以直接誘導結縷草外植體分化成為胚性愈傷組織。在配方2、配方3處理下外植體產生的愈傷組織,均有部分存在松散、緊密性差的情況,而有些則再分化成了根,這表明配方2、配方3也可以誘導結縷草產生愈傷組織,但產生的愈傷組織多為普通的愈傷組織。
2.2 體胚細胞的誘導
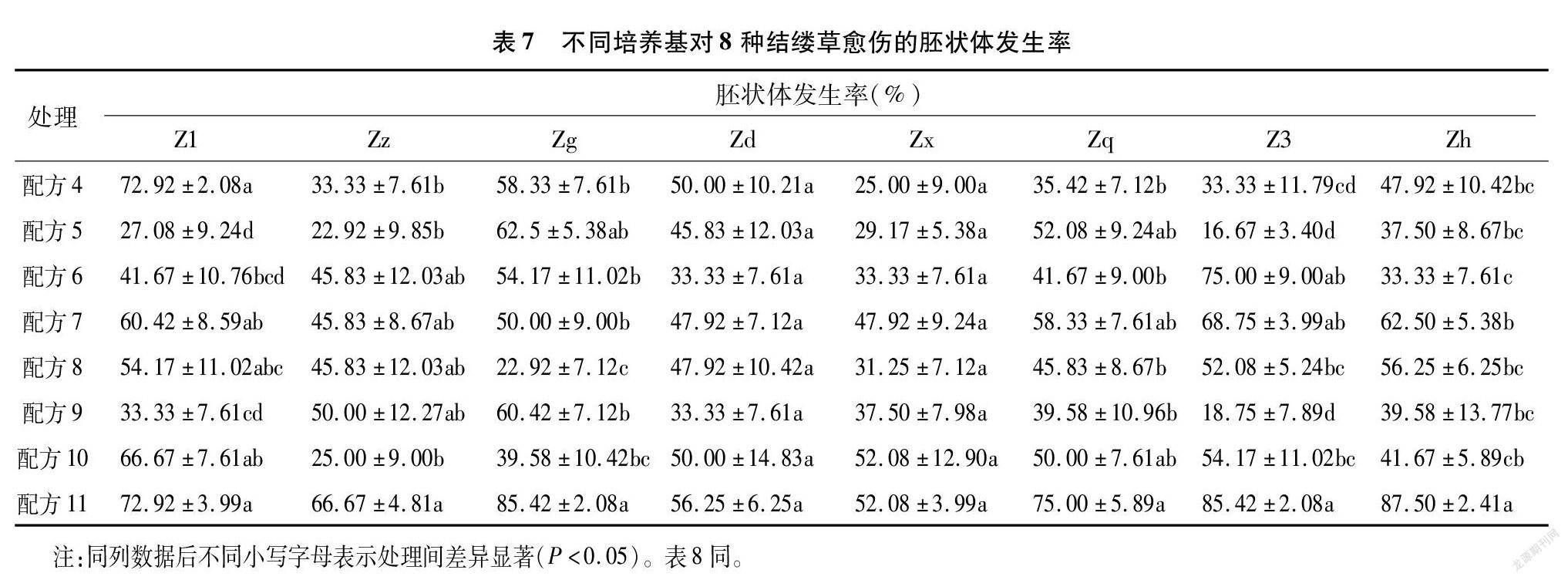
將愈傷組織轉接到體胚細胞誘導培養基上進行進一步培養,可獲取體細胞胚狀體。隨著時間推移,用肉眼即可觀察出胚狀體的發生情況與發育狀態。本試驗以出現肉眼可見的胚狀體(即球形胚、心形胚、魚雷形胚、子葉形胚)為判定標準。從表7可以看出,總體以配方11處理下的8種結縷草愈傷組織出現的胚狀體數量最多,胚狀體發生率最高。Zd、Zx這2種結縷草愈傷組織在不同配方處理下,胚狀體發生率無顯著性差異;而其余品種結縷草在不同配方處理下,存在顯著性差異。配方7、配方8也能適合大多數的結縷草愈傷組織發育成為胚狀體。
2.3 胚狀體的增殖與生長
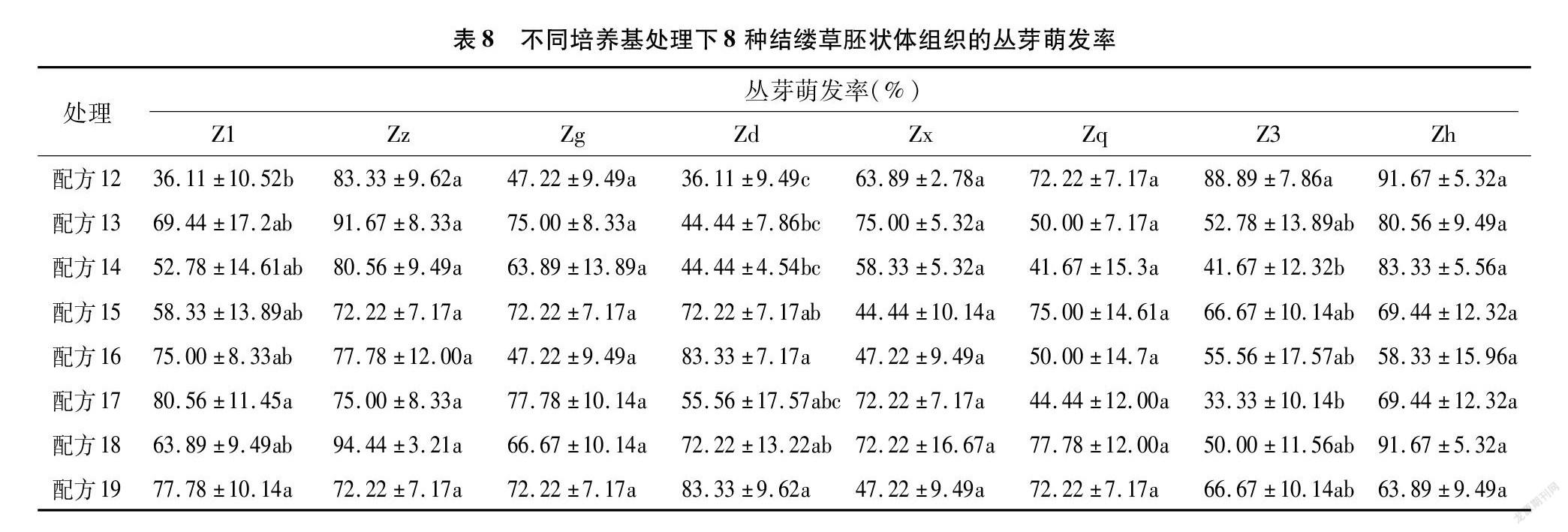
將通過體胚細胞誘導培養基處理過后產生的胚狀體接種到胚狀體增殖與生長培養基上,進行胚狀體的增殖與芽的誘導。從表8可以看出,8種配方處理下的結縷草胚狀體,幾乎每種都能誘導產生叢芽。Zz、Zg、Zx、Zq、Zh這5種結縷草胚狀體對不同的培養基配方敏感性并不顯著,在8種配方處理下都產生了不定芽且無顯著性差異。Z1、Zd、Z3則表現出明顯的差異性,Z1以配方17處理下的胚狀體芽萌發率最高,Zd則以配方16和19處理下的萌發率最高,Z3則是配方12處理下的最高。綜合來看,配方18與配方19可以有效地誘導大多數結縷草胚狀體產生芽,配方15、配方16、配方17也可以誘導大多數的結縷草胚狀體產生芽,不過對部分種質基因型結縷草胚狀體的效用會略低于配方18與配方19。
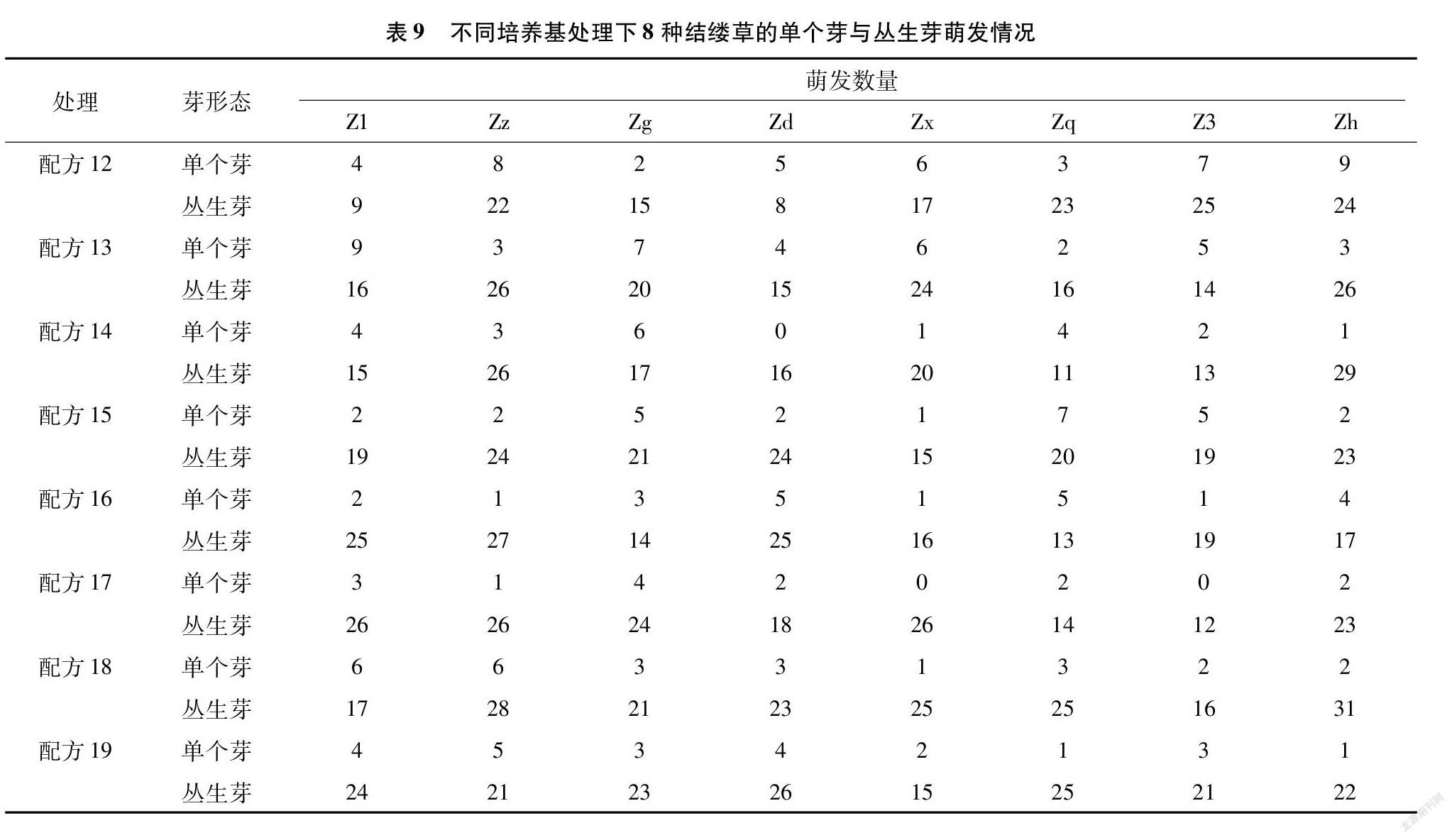
由胚狀體發育而來的芽可以細分為單個芽與叢生芽。從表9的數據可以看出,相對于單個芽而言,結縷草胚狀體更容易分化出叢生芽。配方14處理下的Zd與配方17處理下的Z3的芽萌發率很低,但萌發出來的芽均為叢生芽;而配方17處理下的Zx的芽萌發率為幾種配方里面最高的,但萌發出來的芽也均為叢生芽,這或許與結縷草之間不同的基因型有關。
2.4 結縷草的成苗快繁培養
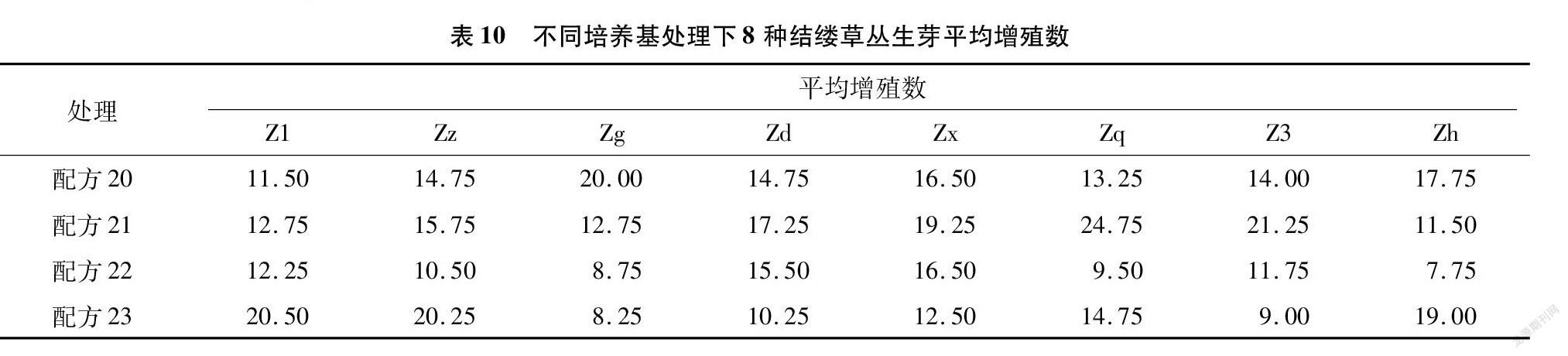
從表10可以看出,8種結縷草在不同配方處理下的增殖數都有很大的差異,極差最大的為配方21與配方22處理下的Zq,極差達到了15.25,極差最小為配方20與配方22處理下的Zx,極差為0。Z1、Zz、Zd、Zx這4種結縷草的增殖數在4種配方處理下都超過了10,表明這4種結縷草更容易快速繁育。
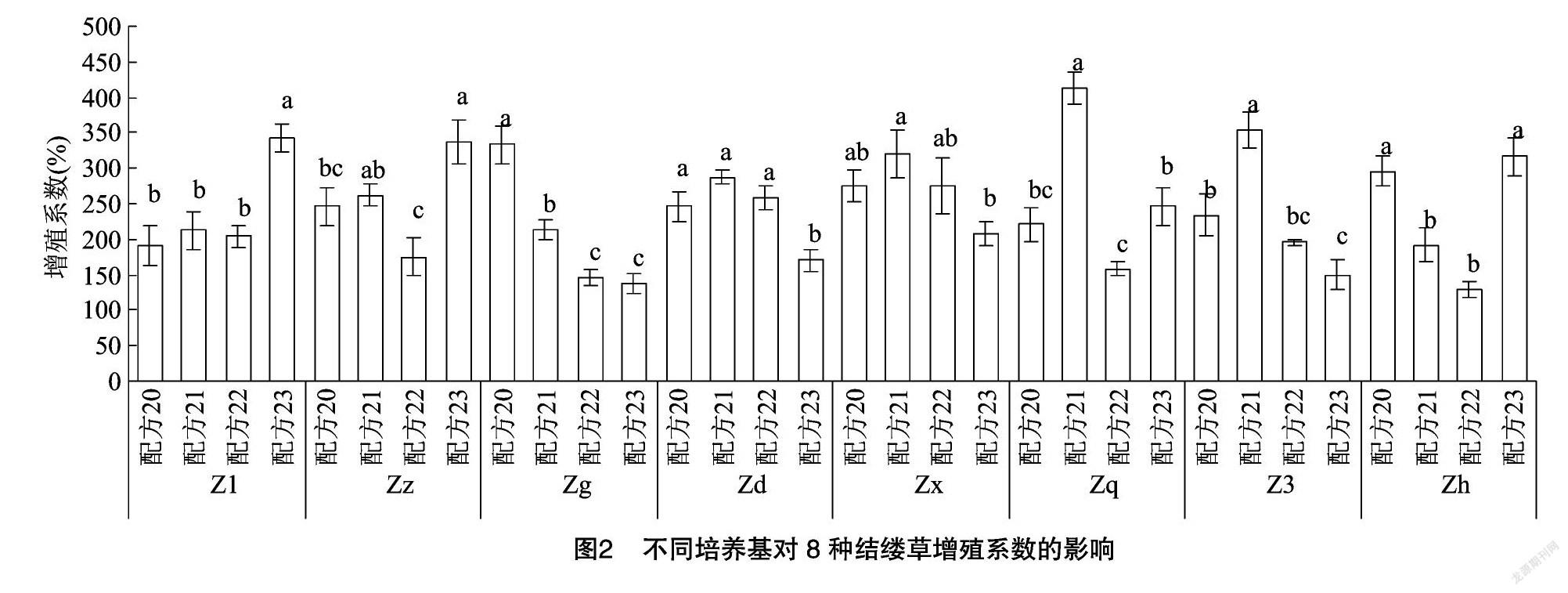
對圖2進行分析,可以發現在不同配方處理下的每種結縷草增殖系數都存在顯著性差異。但很難找到可以最大化促進8種結縷草快繁的配方。Z1、Zz、Zh在配方23處理下的增殖系數最高,與其他配方間存在顯著性差異;Zd、Zx、Zq、Z3在配方21處理下的增殖系數最高,與其他配方間存在顯著性差異;而Zg則在配方20處理下的增殖系數最高,與其他配方間存在顯著性差異。
2.5 壯苗生根培養與植株移栽
從表11可以看出,純MS培養基即可有效地促進結縷草根的分化,加入IBA后部分植株根的生長受到了影響,且濃度越高,根的分化就越受到抑制。根的良好生長也很大程度上促進移栽后成活率的提高,本試驗中根部生長良好的結縷草移栽成活率為100%。
3 討論與結論
結縷草愈傷組織的難獲取曾很長時間限制著結縷草組培快繁體系的建立。本試驗中通過添加高濃度的生長素2,4-D與低濃度的6-BA配合使用,成功獲得了8種結縷草的愈傷組織,這表明高濃度的2,4-D對結縷草愈傷的獲取起著至關重要的作用,這與前人的研究結果一致。柴明良等曾通過將萌發的結縷草種子接種到高濃度2,4-D的MS培養基上以獲得愈傷組織,但獲得的愈傷組織卻未能成功再分化[6]。Bhaskaren等的研究表明,2,4-D在大多數禾本科植物的愈傷誘導中都扮演著重要的角色[7]。張磊等將2,4-D、NAA、IAA、6-BA這4種激素單獨使用,發現以2,4-D處理的愈傷誘導率最高[8],本試驗結果與之也基本一致。但張磊等試驗獲得的愈傷誘導率高達64.6%[8],本試驗中愈傷誘導率最高也僅有51%,或許與這2個試驗的愈傷誘導配方不一樣有關。
一般外植體脫分化后獲得的愈傷可分為胚性愈傷與普通愈傷,胚性愈傷可進一步分化成完整植株,而體胚細胞的誘導主要經歷愈傷組織誘導階段和分化培養階段[9]。普通愈傷組織經誘導培養后一部分分化出胚性細胞,單個胚性細胞再分裂和分化形成一個由多細胞組成的原胚,即胚性愈傷組織[10]。賈玉芳等研究表明,MS+2.0 mg/L 2,4-D的組合效果最好,胚性愈傷組織比率為8583%[11],本試驗中,最適合胚性愈傷誘導的培養基配方為改良MS+0.1 mg/L 6-BA+0.2 mg/L NAA+2 mg/L 2,4-D,胚性愈傷組織的比率最高可達到88%,這與前人研究結果基本一致。Linacero等[12]和錢永強等[13]都證明了過高或過低的2,4-D濃度都不利于胚性愈傷組織的形成和分化作用。本試驗中部分結縷草在含4 mg/L 2,4-D配方處理下的體胚細胞誘導率極低,這表明高濃度的2,4-D會抑制體胚細胞的形成,但也有部分結縷草在含2 mg/L 2,4-D配方處理下的體胚誘導率較低,這表明低濃度的2,4-D也會抑制體胚細胞的形成,這與前人研究成果基本一致。但本試驗中幾種結縷草在高濃度或低濃度的2,4-D處理下的結縷草均有較高的體胚細胞誘導率,這或許與結縷草間的基因型不同或有其他激素參與進來共同作用有關,具體的機理還有待進一步探討。
有學者研究表明,在分化階段,暗培養條件后,愈傷組織通過分化成胚性愈傷組織,經體細胞發生途徑產生再生植株的數量大大高于光培養條件下愈傷組織的分化[14-18]。本試驗采取的即此途徑,胚狀體芽的分化率最高可達到94%,這與前人的研究成果基本一致。杜敏華等研究表明,0.5 mg/L NAA最適于結縷草芽的分化[19],本試驗中,配方12、13、16、17的NAA含量均為0.5 mg/L,且大多數能促進芽的分化,具有顯著性差異,這與前人的研究成果基本保持一致,至于部分結縷草的體胚細胞未能促進芽萌發,可能與其基因型有關。叢生芽的生命力與繁殖力明顯高于單個芽[20],本試驗的配方誘導出來的不定芽多數為叢生芽,為結縷草的成苗快繁建立了良好的基礎。
王棟的研究表明,培養基MS+0.1 mg/L IBA+0.1 mg/L 6-BA最有利于試管苗芽的增殖,芽數平均可增殖至9個以上[21],本試驗未能找出最適合各種結縷草快繁最佳培養基配方,但本試驗中使用到的配方處理后新增平均芽數均能超過9,較前人試驗結果較高,這可能與本試驗中激素濃度較高有關。宋俊芳等的研究結果表明,添加1 mg/L的IBA就能明顯促進根的分化[22],本試驗的結果與之不符,可能與種苗狀態有關,也有可能與結縷草間基因型有關,具體原因有待進一步探討。
本試驗旨在尋找出一種無種質基因型限制的結縷草組培快繁體系。通過試驗得出,愈傷組織誘導培養基為MS+2 mg/L 2,4-D+0.2 mg/L 6-BA+40 g/L 蔗糖最為合適,體胚細胞誘導培養基為改良MS+2 mg/L 2,4-D+0.2 mg/L NAA+0.1 mg/L 6-BA+40 g/L蔗糖最為合適,胚狀體增殖與生長培養基為MS+0.1~0.2 mg/L 2,4-D+1 mg/L NAA+0.1 mg/L 6-BA+30 g/L蔗糖最為合適,成苗快繁培養基為MS+0.02~0.05 mg/L NAA+1~2 mg/L 6-BA+30 g/L蔗糖最為合適。
參考文獻:
[1]閆曉玲,趙新爍. 結縷草愈傷組織誘導的影響因素[J]. 農村經濟與科技,2016,27(18):34-35.
[2]吳艷歌,畢 波,柴明良. 細胞分裂素對溝葉結縷草愈傷組織生長及其抗氧化系統的影響[J]. 核農學報,2016,30(7):1288-1295.
[3]馬龍雪,趙麗萍. 大穗結縷草愈傷組織誘導及植株再生研究[J]. 黑龍江農業科學,2016(9):15-18.
[4]路 佳,葛榮朝,孫振元,等. 日本結縷草莖節和愈傷組織再生體系的優化[J]. 河北師范大學學報(自然科學版),2011,35(5):519-526.
[5]王渭霞,胡張華,陳錦清,等. 松南結縷草成熟胚愈傷組織的誘導和再生[J]. 草業學報,2006,15(3):132-137.
[6]柴明良,郭達初,鈕友民,等. 三種結縷草試管繁殖研究[J]. 科技通報,1993,9(6):411-415.
[7]Bhaskaran S,Smith R H.Regeneration in cereal tissue culture:a review[J]. Crop Science,1990,30(6):1328-1337.
[8]張 磊,吳殿星,胡繁榮,等. 結縷草組織培養及農桿菌介導轉化的主要因子優化[J]. 草業學報,2004,13(4):100-105.
[9]魏文霞,劉苗苗,張婷婷,等. 黃瓜胚狀體途徑再生體系的建立[J]. 西北農林科技大學學報(自然科學版),2019,47(4):32-40.
[10]何業華,方少秋,馬 均,等. 菠蘿愈傷組織中體細胞胚起源過程的組織細胞學觀察[J]. 園藝學報,2010,37(5):689-696.
[11]賈玉芳,陳 曙,柴明良. 溝葉結縷草胚性愈傷組織繼代培養基對其生長和再生的影響[J]. 園藝學報,2010,37(8):1311-1316.
[12]Linacero R,Vazquez A M.Somatic embryogenesis from immature inflorescences of rye[J]. Plant Science,1990,72(2):253-258.
[13]錢永強,孫振元,韋善君,等. 中華結縷草成熟胚再生影響因素研究[J]. 核農學報,2005,19(6):436-440.
[14]史滟滪,劉艷軍,楊靜慧,等. 花椰菜松散型胚性愈傷組織的誘導[J]. 天津農業科學,2019,25(5):9-12.
[15]史滟滪,楊靜慧,劉 婷,等. 馬鈴薯松散型胚性愈傷組織的誘導[J]. 天津農業科學,2016,22(12):20-23.
[16]溫 琳,惠國強,楊海鵬,等. 半夏葉柄愈傷組織誘導與分化最適培養基篩選[J]. 山西農業科學,2017,45(6):905-908,977.
[17]張朝暉,張宗申,陳 陽,等. 雜交構樹體細胞胚誘導研究[J]. 河南農業科學,2016,45(12):127-131.
[18]李海霞,謝久鳳,孫金花. 玉米幼胚胚性愈傷組織誘導和繼代研究[J]. 江蘇農業科學,2020,48(7):74-77.
[19]杜敏華,柴春月. 日本結縷草離體培養及高頻率植株的再生[J]. 東北林業大學學報,2007,35(10):17-19.
[20]趙玉芬,儲博彥,尹新彥,等. 白砂糖對大花萱草‘紅運’試管苗生根的影響[J]. 林業科技開發,2013,27(4):30-32.
[21]王 棟. 溝葉結縷草組織培養及植株再生的研究[D]. 重慶:西南大學,2008.
[22]宋俊芳,金 芳,孫吉雄,等. 蘭引3號結縷草組織培養研究[J]. 草原與草坪,2007,27(5):29-31,36.
3262500338270
