茶小綠葉蟬侵害對茶樹生長和茶葉品質影響的研究進展
2022-03-18 09:11:50楊子銀吳淑華辜大川
茶葉通訊 2022年1期
楊子銀,吳淑華,辜大川
1. 中國科學院 華南植物園,廣東 廣州 510650;2. 中國科學院大學,北京 100049
茶小綠葉蟬是我國各大茶區均有分布的昆蟲,長期以來茶小綠葉蟬優勢種的分類歸屬問題一直存在爭議,直到近些年才形成較為統一的意見。茶小綠葉蟬侵害會造成茶葉減產,導致茶產業經濟損失嚴重。茶小綠葉蟬侵害對于茶鮮葉的危害特點主要包括外形和內含物質變化方面。適度的茶小綠葉蟬侵害會誘導一些有利茶葉品質的物質形成,但由于茶小綠葉蟬繁殖速度快且世代重疊,在實際生產中蟲口密度難以控制,這不僅限制了茶小綠葉蟬在茶葉品質提升方面的應用,同時防控難度也較大。本文主要總結了茶小綠葉蟬侵害對茶樹生長、茶葉品質代謝物影響的研究進展,并提出未來的主要研究方向包括利用預警措施和蟲口密度控制技術提升茶葉品質、挖掘茶小綠葉蟬關鍵激發子、闡明茶小綠葉蟬侵害對茶葉品質和產量影響的內在機制與品質-產量調控的重要節點。
1 茶小綠葉蟬簡介
1.1 優勢種及遺傳變異
茶小綠葉蟬,又稱葉跳蟲、浮塵子,屬小綠葉蟬屬半翅目葉蟬科,廣泛分布于中國、日本、越南、印度及其他亞洲國家的產茶區,是茶葉生產中危害最嚴重的茶樹害蟲之一[1-2]。我國茶園小綠葉蟬共發現60多種,包括小貫小綠葉蟬(Empoasca onukii Matsuda)、煙翅小綠葉蟬(Empoasca limbifera)、擬小莖小綠葉蟬(Empoasca paraparvipenis)等[3]。不同種的小綠葉蟬形態特征及體色十分接近,且蟲體較小,因而中國茶園小綠葉蟬的優勢種分類歸屬問題一直存在爭議,且須根據雄性外生殖器特征才能將其區分。早年葛鐘麟等[4]和趙冬香等[5]通過鑒定認定茶小綠葉蟬的優勢種為假眼小綠葉蟬;隨后秦道正等[6]、施龍清等[7]、于曉飛等[8]和孟召娜等[9]通過顯微照相機、解剖鏡、電動體視顯微鏡等技術鑒定后,認定小貫小綠葉蟬為我國主要產茶區域茶園的優勢種。小貫小綠葉蟬早在1971年首次被記錄在中國分布,但未指明是否侵害茶樹[10]。在日本,小貫小綠葉蟬在1952年被認定為茶樹的為害種[11]。在印度、韓國等亞洲國家,小貫小綠葉蟬也被報道為害茶樹[12-13]。在中國臺灣地區所報道的臺灣小綠葉蟬(Jacobiasca formosana Paoli)在分子水平上被認為與小貫小綠葉蟬是同一個種,但有待進一步研究確定[8]。
小貫小綠葉蟬的親緣關系與地理距離相關[14],但是不同地理種群的遺傳距離與地理分布不一定對應[15]。在中國,茶樹種植區分布在不同的氣候區,小貫小綠葉蟬對不同棲息地的適應可能導致了種群間的遺傳分化。并且,這些地區內部和地區之間的基因流動被山脈和大河等地理屏障所限制,因而在已確認的區域內和區域間,小貫小綠葉蟬種群之間可能發生遺傳分化[6]。此外,先前的形態學研究表明該蟲群體內部和群體之間的雄性生殖器結構存在變化,這也表明其可能存在顯著的群體水平遺傳變異[6]。然而,隨機引物擴增多態性技術(random amplifiedpolymorphic DNA,RAPD)對小貫小綠葉蟬的歸屬研究結果表明,雖然小貫小綠葉蟬具有較豐富的遺傳多樣性,但其遺傳基礎較為一致,異種的可能性幾乎沒有[16]。隨后,12個小貫小綠葉蟬種群和5個近緣種外群的mtCOI基因序列分析結果也顯示,不同種群間無顯著遺傳分化,種群個體在分子系統樹中呈平行分布[17]。然而,微衛星分子標記的5個不同小貫小綠葉蟬種群和2個近緣種外群的遺傳物質結果卻表明,四川和云南群體存在顯著的遺傳分化,與陜西、山東和河南群體的遺傳多樣性相似[18]。以往研究結果的不同,可能是由于研究中使用的遺傳標記較少,變異程度較低,達不到研究群體結構的要求。因此,人們對茶小綠葉蟬的種群結構所知甚少。近期通過對小貫小綠葉蟬基因組染色體水平的組裝和注釋,發現關鍵途徑的基因組變化,包括與新陳代謝、晝夜節律和免疫系統功能相關的途徑,這可能是小貫小綠葉蟬成功傳播和適應的基礎[19]。此外,該研究通過對54份小貫小綠葉蟬的遺傳變異分析,揭示了中國不同茶區茶小綠葉蟬的種群結構和進化歷史,為昆蟲對寄主植物的適應提供了研究基礎,進而促進可持續的害蟲管理[19]。
1.2 形態特征及種群動態
茶小綠葉蟬屬不完全變態昆蟲,完成一個世代需經歷卵、若蟲和成蟲三個階段,共40 d左右[20]。該蟲繁殖能力強,繁殖速度快,世代重疊。若蟲體長約2.5 ~ 3.5 mm,體色淡綠至淡黃,體型較成蟲略小,無翅,其他特征與成蟲相似[20]。成蟲體長約3.1 ~ 3.8 mm,體色黃綠至淺黃。前翅為白色、淡黃色、半透明色,后翅為透明膜狀,爪呈棕色,每只足脛骨神經節末端為綠色[21];頭頂有兩個綠色斑點,頭部有較大灰褐色復眼,位于頭部兩側,占整個頭部的一半;在復眼內部密集排列了蜂窩狀正六邊形小眼[22]。雌蟲和雄蟲觸角均由鞭節、梗節和柄節構成,均呈剛毛狀且有小球形網粒體分布,感受器為鱗形和毛形[22]。小球形網粒體的存在有助于成蟲逃離蜘蛛網[23]。雌雄異型,雌性成蟲體型比雄性成蟲大,體色相對更深,壽命更長[24];雄性外生殖器的下生殖板呈三角形,尾節突弧狀彎曲,陽莖位于陽莖干端部且無突起和背腔,肛突端部弧形彎曲且末端不尖銳,這些特征為分類的主要依據[9]。茶小綠葉蟬生長季在茶樹嫩莖上成批產卵,在茶樹或雜草中越冬,越冬的成蟲可產卵1個月,每只雌性成蟲有10 ~ 130個卵,呈新月形,長約0.8 mm,最初為乳白色,后期為淡綠色[21]。
茶小綠葉蟬發生隱蔽,通過吸食茶嫩葉(背面)或嫩莖(韌皮部)的汁液造成傷害[2]。成蟲和若蟲均畏強光怕濕,主要依賴視覺和嗅覺遠程定位宿主,其中視覺占主導作用,因此光和環境顏色會干擾茶小綠葉蟬的準確定位[22]。成蟲白天吸食葉背面的汁液,晚上棲息于茶樹枝條下,喜歡乘風飛行和傳播[21]。茶小綠葉蟬的主要活動時間為4 ~ 11月,一年內有兩個爆發高峰,分別是 5 ~ 6 月和 9 ~ 10 月,常年發生蟲害面積占茶葉總種植面積的75%以上[3,21]。茶小綠葉蟬分布廣泛,南至海南、廣東和廣西等省,北至安徽,均有不同程度的為害[25],發生代數受地理位置及氣候環境的差異而不同,華中地區 9 ~ 11 代、華南地區 11 ~ 13 代、海南島13代以上[3,20]。茶小綠葉蟬的種群動態和活動規律受溫度、濕度、光照及風速的影響[3,26]。其中,溫度對茶小綠葉蟬的影響最大,主要影響其存活率、著卵量、保護酶活性變化、總抗氧化能力和總蛋白含量,最適溫度為17℃ ~28℃,平均溫度低于10℃或高于29℃,則蟲口數量急劇下降[3,27,28]。此外,不同的光周期和波長會影響它們行為活動及繁殖和發育[29]。
2 茶小綠葉蟬侵害對茶樹生長的影響
茶小綠葉蟬是廣泛分布于我國各大茶區的主要害蟲,通常會導致夏、秋茶損失10% ~30%,個別為害嚴重年份達到50%以上,居茶樹病蟲害之首[30-31]。據推算,每百梢茶梢被140頭茶小綠葉蟬侵害10 d后,經濟損失率達到20%以上[32]。每年茶樹病蟲害總防治費用的40% ~ 60%用于茶小綠葉蟬的防治[30]。
茶小綠葉蟬侵害會顯著抑制茶樹的生長。茶小綠葉蟬若蟲與成蟲均通過刺吸茶樹嫩梢芽葉攝取汁液。此外,雌性成蟲還會在茶樹嫩梢內產卵,導致嫩梢輸導組織受損,水分與營養供應不足[33-34]。一般而言,被若蟲和成蟲刺吸后的茶葉嫩梢,呈現典型的葉蟬燒癥狀[35],即受害茶樹芽葉葉緣變黃、葉脈變紅、葉片粗老和卷曲,受害嚴重時葉尖和葉緣呈現紅褐焦枯,芽葉生長停滯,嚴重時全葉焦枯易脫落,葉片質地變脆,莖節節間變短[33-35]。2000年,日本學者對茶小綠葉蟬侵害影響茶樹葉脈褐變和新芽生長進行了更為細致的研究[36]。結果表明,經一只中期若蟲侵害1 d,葉脈顏色變成棕色。侵害4 d后,葉脈全部變為褐色。然而,一只雌蟲侵害導致的棕脈芽數比一只若蟲侵害導致的少。中齡期若蟲或雌蟲侵害后,莖上至莖下均出現褐紋。中齡期若蟲或雌蟲侵害1 d,中脈處出現點狀斑點。第二、三葉上的斑點比更上部位葉上的斑點多。褐脈芽的比例與新芽的長度有極顯著的相關性,但與新芽的重量無顯著的相關性。隨著技術的發展,直流刺探電位圖譜系統被用于茶小綠葉蟬的口器刺吸行為研究。結果表明,茶小綠葉蟬可能主要從茶樹的韌皮部攝取汁液,部分從葉肉攝取[2]。
近年來,對普通綠茶品種“福鼎大白茶”和葉綠素缺乏的白化品種“黃金芽”進行茶小綠葉蟬侵害后的形態、生理和生化變化等方面的研究,結果表明,茶小綠葉蟬對于茶樹生長的影響是全方面的,并且存在著品種差異[37]。茶小綠葉蟬侵害均降低了“福鼎大白茶”和“黃金芽”茶樹芽長、葉面積、葉厚和莖粗。此外,茶小綠葉蟬侵害還降低了兩個茶樹品種葉片上表皮、柵欄組織和海綿組織的厚度,降低了莖節間的薄壁組織厚度和髓徑。茶小綠葉蟬侵害降低了“黃金芽”葉片中葉綠素a、葉綠素b和類胡蘿卜素的含量,進一步影響了光合速率,但對“福鼎大白茶”無顯著影響。在茶小綠葉蟬侵害下,光系統II的最大量子產率和實際光化學效率以及“黃金芽”的非光化學猝滅受到抑制。以上結果表明,茶小綠葉蟬對于茶樹芽長、葉面積、葉厚和莖粗等生長相關指標的影響在不同品種中是保守存在的,即茶小綠葉蟬會顯著抑制不同品種茶樹的生長。另一項關于茶小綠葉蟬侵害茶樹的研究也得出類似的結果,茶小綠葉蟬侵害后的茶樹鮮葉主側脈變紅、葉片萎縮、葉面積減少、葉片重量下降、葉尖和葉緣呈現紅褐焦枯、莖節節間縮短等[38]。
盡管不同茶樹品種的生長均受到茶小綠葉蟬的抑制,但不同茶樹品種仍表現出了對于茶小綠葉蟬抗性的差異,并且抗性差異可能與茶樹形態、組織或者細胞差異相關。此前研究表明,茶梢上茶小綠葉蟬蟲口密度與茶樹葉片主脈下方表皮和下方厚角組織厚度、柵欄和海綿組織厚度呈極顯著負相關[39]。隨后的另一項關于30份茶樹種質對茶小綠葉蟬的抗性研究也表明,茶樹抗性與葉片上表皮細胞數、葉肉厚度、海綿和柵欄組織厚度皆呈負相關[33]。然而最近通過茶小綠葉蟬蜜露排泄量反證茶樹品種(系)抗蟲性的研究則表明,不同品種(系)茶樹的葉片總厚度、上表皮和下表皮厚度、葉面積以及色差等指標與雌成蟲的蜜露排泄量之間無顯著相關性;葉片茸毛的長度與蜜露排泄量呈現顯著正相關性,而葉片茸毛密度則與雌成蟲蜜露排泄量呈顯著負相關性[40]。換言之,葉片茸毛密度較大的茶樹可能抗葉蟬的能力較強,葉片茸毛長度較長的茶樹可能抗葉蟬能力較弱。
綜上所述,茶小綠葉蟬的若蟲和成蟲均會刺吸茶樹嫩梢取食,使茶樹嫩梢呈現典型的葉蟬燒表型。茶小綠葉蟬的侵害在不同茶樹品種中均抑制了茶樹的生長,導致一系列茶樹生長相關指標的改變,最終導致茶葉的減產;并且不同茶樹品種對于茶小綠葉蟬的抗性存在差異,可能與茶樹形態、組織或者細胞差異相關。然而目前關于茶小綠葉蟬侵害影響茶樹生長的研究僅停留在侵害后對茶樹表型變化的描述以及一些生長相關指標的測量,缺乏更為深入的分子機制解析,這也是今后研究需要加強探究的方向。
3 茶小綠葉蟬侵害對茶葉品質的影響
盡管茶小綠葉蟬侵害抑制茶樹生長,導致茶葉產量顯著降低。然而,適度的茶小綠葉蟬侵害有利于茶葉品質提升,如著名的烏龍茶“東方美人茶”就是采用茶小綠葉蟬侵害后的茶鮮葉為原料制作加工而成。茶樹在遭遇茶小綠葉蟬侵害后,茶樹葉片會產生次生代謝產物[41]。其中一些代謝物一方面幫助茶樹抵御昆蟲侵害,另一方面可作為茶葉品質成分影響茶葉品質。
3.1 揮發性代謝物
茶小綠葉蟬侵害會誘導茶葉中反式-β-羅勒烯、正辛醇、芳樟醇、(E)-β-法尼烯、水楊酸甲酯、苯甲醛和吲哚含量上升[42]。采用茶小綠葉蟬和茉莉酸甲酯分別處理茶葉后,(E)-β-羅勒烯、(E)-4, 8-二甲基 -1, 3, 7-壬三烯和 (E,E)-α-法尼烯這三種化合物分別占總誘導揮發性混合物的近90%和74.2%[43],暗示茶小綠葉蟬可能是通過茉莉酸途徑誘導了這三種揮發性物質的產生。相比于健康茶梢中的揮發物,茶小綠葉蟬侵害后的茶樹新梢新出現12種組分,其中紫蘇烯含量較高[44]。
茶葉中的揮發性物質,盡管大多數含量很低,但具有特殊的氣味,在茶葉香氣品質中發揮了重要作用。茶小綠葉蟬為害程度不同,誘導的茶葉樣品中香氣物質的組成種類和比例也會不同。檢測茶小綠葉蟬不同為害時間的茶葉樣品,烷類物質總含量呈現減少,而醇類、醛類物質總含量呈現增加,并且隨著為害程度的增加,出現對二甲苯、四氫薰衣草醇、2-乙基己醇、鯨蠟醇、角鯊烯、茶香螺烷、2-丁基-1-辛醇和柏木腦等新的香氣物質[45]。在對廣東地區茶小綠葉蟬侵害“金萱”品種茶葉揮發性成分的研究表明,茶小綠葉蟬為害鮮葉后,萜醇含量減少,脂肪醇、芳香醇、萜酮、脂肪酮、脂類和烴類增加,而制作的紅茶揮發性成分中萜醇、脂肪醇、萜酮、脂肪酮、烴類和酸類含量降低,脂類增加[46]。研究者進一步通過主成分和聚類分析結果表明,萜醇和酯類物質是區分不同為害程度鮮葉及其紅茶的主要揮發性成分[46]。金珊等[47]以茶小綠葉蟬為害蟲口密度最大和最小的烏龍茶茶樹品種肉桂和鐵觀音為研究材料,分析了蟲害誘導型揮發代謝物,結果表明,鐵觀音和肉桂在蟲害后都釋放大量的β-月桂烯、β-羅勒烯、反式-β-羅勒烯、α-法呢烯和芳樟醇;此外,茶小綠葉蟬侵害還誘導了(Z)-丁酸-3-己烯酯、癸醛、吲哚、己酸-3-己烯酯、苯乙醇這些新的揮發物產生。隨后一項關于茶小綠葉蟬蟲口密度和葉片損害面積與代謝物關系的研究則表明,茶小綠葉蟬的蟲口密度對于揮發物的產生具有非常顯著的影響,而葉片損傷的影響僅略微顯著[41]。對茶小綠葉蟬密度最敏感的揮發物主要是濃度隨蟲口密度增加而增加的萜類化合物,而對葉片損傷最敏感的揮發物主要是脂肪酸衍生物和揮發性苯丙烷/苯類[41]。最近關于茶小綠葉蟬侵害茶梢的揮發性物質釋放曲線研究結果發現,偏最小二乘判別分析模型可以區分健康和受茶小綠葉蟬侵害茶梢之間的揮發性物質特征,預測的準確率達91.8%,并且確定了八種關鍵揮發物是重要的預測變量[48]。表1整理了近年來不同茶樹品種受茶小綠葉蟬侵害后在茶鮮葉以及干茶茶湯中產生或含量增加的釋放和內在揮發性代謝物。
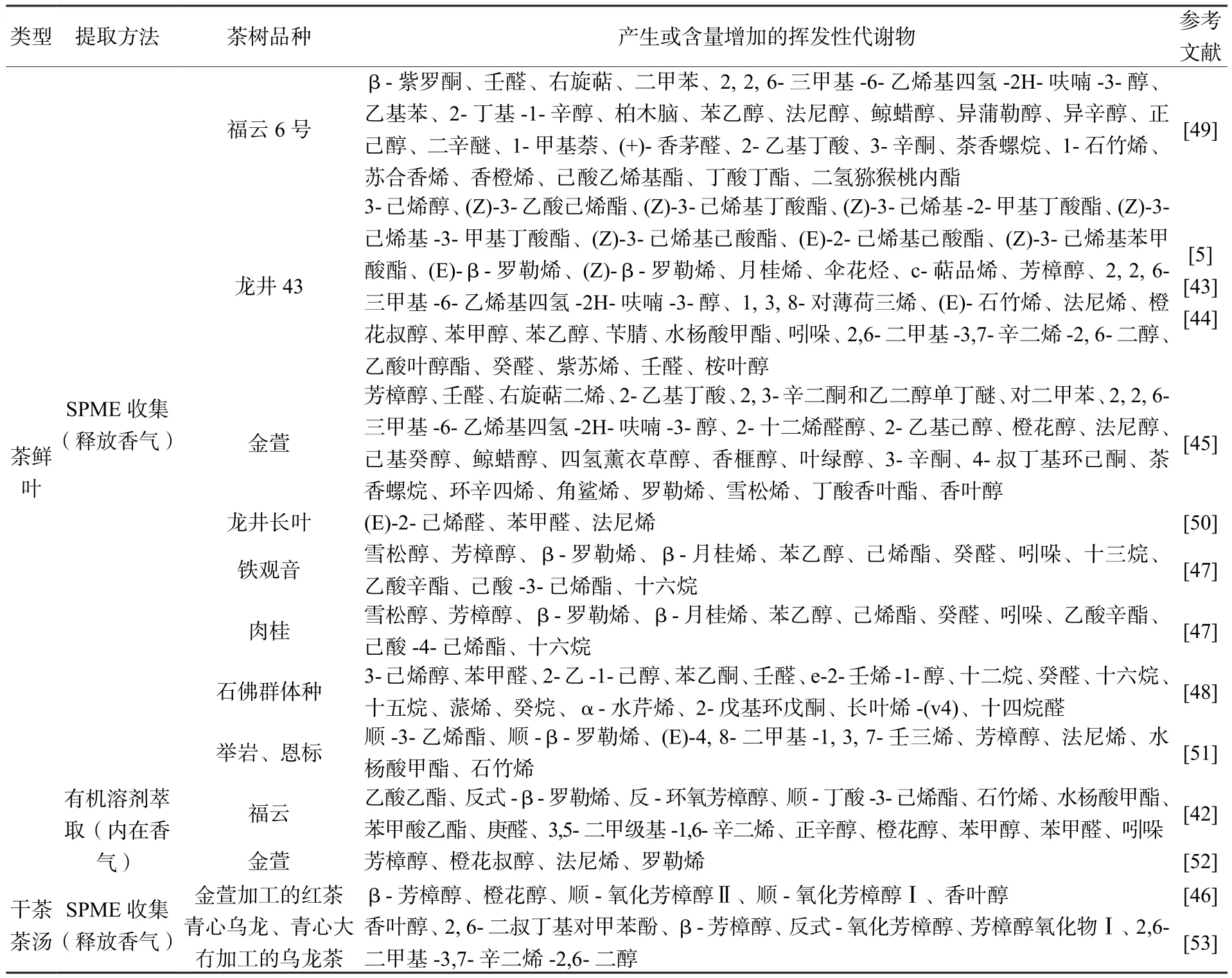
表1 茶小綠葉蟬侵害后茶鮮葉和干茶茶湯中發生變化的揮發性代謝物Table 1 Changes of volatile metabolites in tea green leafhopper infested fresh tea leaves and its tea product infusion
近年來,隨著技術的發展,轉錄組、蛋白組和代謝組等多組學技術相繼被應用于茶學研究,推動了茶學研究的發展。表2總結了已發表的茶小綠葉蟬侵害茶樹相關組學數據信息,從基因、蛋白和代謝物水平更為全面地解析了茶小綠葉蟬侵害對于茶樹的影響。盡管這些組學數據的茶小綠葉蟬處理條件存在差異,但是我們仍可從共性特征中得到一些將來工作的線索。例如,轉錄組的數據基本都包含了植物激素信號轉導通路,暗示茶小綠葉蟬侵害影響的激素信號轉導通路可能是將來研究工作的重點。然而目前對于組學數據的利用尚不充分,今后對于組學數據的深入挖掘以及多組學數據的聯合分析,也將是未來研究需加強的地方。此外,伴隨分子生物學的發展,一些蟲害誘導的揮發性代謝物合成基因相繼在茶樹中被克隆與鑒定,如萜類合成酶基因[54]、芳樟醇合成酶基因CsLIS1和CsLIS2[52]、α-法呢烯合成酶基因CsAFS[55]和CsAFR[56]、橙花叔醇合成酶基因CsNES[57]和CsNES2[56]、β-羅勒烯合成酶基因CsOS2[58]和CsBOS1[59]、吲哚合成酶基因CsTSA和CsTSB2[60]等。此外,研究者還從茶小綠葉蟬中功能鑒定了萜烯合成酶基因EoTPS[61]。今后對于這些基因的研究將進一步加深人們對于茶小綠葉蟬侵害如何影響茶葉揮發性代謝物的理解。

表2 茶小綠葉蟬侵害茶樹的相關組學數據Table 2 The related omics data of tea plants exposed to tea green leafhopper infestation
3.2 非揮發性代謝物
揮發性代謝物主要貢獻了茶葉的香氣品質,而非揮發性代謝物則主要貢獻了茶葉的滋味品質。非揮發性物質的組成復雜,主要包括多酚類、咖啡堿、氨基酸和可溶性糖等,茶多酚貢獻了茶湯滋味中的苦澀味;咖啡堿貢獻了茶湯滋味中的苦味;氨基酸貢獻了茶湯滋味中的鮮爽醇味;可溶性糖貢獻了茶湯滋味中的甜味[69]。除對揮發性物質有影響外,茶小綠葉蟬侵害對非揮發性物質同樣影響顯著。隨著茶小綠葉蟬為害時間的增加,鮮葉的水浸出物、茶多酚、黃酮含量發生變化,咖啡堿含量降低,氨基酸含量呈現平緩增加趨勢,可溶性糖含量呈現先增加后緩慢減少的趨勢;不同為害程度鮮葉的氨基酸和咖啡堿含量均下降,而可溶性糖含量增加[45]。另一項研究表明,茶小綠葉蟬取食使茶葉中葉綠素a、葉綠素b、總葉綠素和可溶性碳水化合物含量明顯下降,茶多酚含量增加,并且可溶性蛋白含量與茶小綠葉蟬密度的增加和侵害時間的延長呈正相關[70]。然而,也有研究表明,茶小綠葉蟬侵害茶葉后,鮮葉中氨基酸、茶多酚、兒茶素含量下降,咖啡堿總量略微上升,可溶性糖總量沒有明顯變化[38]。近期通過代謝組學手段,發現黃烷醇類物質、黃酮類化合物、黃酮、黃酮苷、黃酮醇苷和可水解單寧在茶小綠葉蟬侵害后顯著增加,谷胱甘肽和茶氨酸含量在茶小綠葉蟬侵害后呈現下降趨勢,并且植物激素水楊酸和茉莉酸在茶小綠葉蟬侵害后呈現上升趨勢[63-64]。除水楊酸和茉莉酸外,脫落酸也被報道在茶小綠葉蟬侵害后增加,茶黃素被茶小綠葉蟬侵害所誘導[71]。最新的研究發現,葉片損傷(受損葉面積百分比)而非茶小綠葉蟬蟲口密度,以劑量依賴方式顯著降低總多酚、表沒食子兒茶素沒食子酸酯和可可堿濃度[41]。茶小綠葉蟬侵害金萱、福云6號、金觀音、青心大冇、青心烏龍、鐵觀音、梅占、黃金桂等茶樹的鮮葉加工成紅茶或烏龍茶,其水浸出物、茶多酚、氨基酸、咖啡堿、水浸出物、可溶性糖、茶黃素、茶紅素和茶褐素的含量也發生了變化[45,72]。表3整理了近年來不同茶樹品種受茶小綠葉蟬侵害后在茶鮮葉以及干茶茶湯中變化的非揮發性代謝物。關于茶小綠葉蟬侵害誘導茶葉中非揮發性物質生成和變化的研究相對較少,一些研究結果甚至呈現相反的趨勢,這很可能與茶小綠葉蟬處理的條件、方式、時間、蟲口密度和茶樹品種等一系列的因子相關。因此,未來關于闡明特定處理條件下茶小綠葉蟬侵害調控茶葉中非揮發性代謝物生成的工作將十分有意義。同時,盡管部分非揮發性代謝物在不同的茶小綠葉蟬侵害研究中呈現的趨勢可能不一致,但是含量在茶小綠葉蟬侵害后幾乎都發生了顯著性的改變,表明了茶小綠葉蟬侵害對非揮發物含量具有顯著的影響。

表3 茶小綠葉蟬侵害后鮮葉及茶湯中變化的非揮發性代謝物Table 3 Changes of non-volatile metabolites in tea green leafhopper infested fresh tea leaves and its tea product infusion
4 展望
4.1 利用茶小綠葉蟬預警措施與蟲口密度控制技術提升茶葉品質
茶葉采摘具有較強的季節性,以春茶的品質最好,而占全年總產量約60%的夏秋茶因其品質較差常常未得到有效利用,從而造成茶葉資源的極大浪費[73]。如前所述,茶小綠葉蟬的爆發期也具有極強的季節性,每年有兩個爆發期,并且恰好處于夏秋茶階段。適度的茶小綠葉蟬侵害對于茶葉的品質具有提升作用,因此是否可能通過利用茶小綠葉蟬的爆發期從而提升夏秋茶的品質。很早以前研究者就通過數學模擬的手段揭示茶小綠葉蟬種群數量的動態規律[74]。隨后進一步探索茶小綠葉蟬發生與氣象條件的關系,發現氣溫對茶小綠葉蟬影響最大,并且采用日平均溫度預測的卵盛孵期與實際發生期基本符合,可以提早10 ~ 15 d發布蟲情預報[75]。當前對于茶小綠葉蟬的綠色防控手段多樣,主要有生態防控、生物防治、農業防治和化學防治等[76]。因此,未來利用精準的預警手段預測茶小綠葉蟬的爆發期,從而提前做好綠色防控手段,將茶小綠葉蟬蟲口密度控制在合適的范圍內,為精準提升夏秋茶品質提供有效途徑。
4.2 茶小綠葉蟬激發子發掘與應用
與其他昆蟲侵害不同,茶小綠葉蟬侵害后的茶葉加工成的茶具有獨特的蜜果香。除茶小綠葉蟬侵害造成的損傷所誘導的普遍性代謝物對于品質提升的貢獻外,茶小綠葉蟬特有的激發子在成茶品質提升中起關鍵性作用。然而目前茶小綠葉蟬激發子尚未被發現,但我們仍可從近期的一些相關研究中獲得有價值的線索。已有文獻綜述,昆蟲源激發子大致可分為四類:脂肪酸-氨基酸軛合物、植食性昆蟲唾液中的水解酶、昆蟲識別素inceptin和caeliferins[77]。近期研究表明,茶小綠葉蟬唾液腺勻漿液的上清液所誘導的茶樹防御反應與茶小綠葉蟬取食所誘導的茶樹防御反應類似[78]。最近有學者也通過酶活性測定以及對轉錄組和蛋白組數據的綜合分析,鑒定并分析了在茶小綠葉蟬唾液腺和腸道中特異性表達的蛋白酶[79]。以上這些工作的開展,為將來挖掘茶小綠葉蟬激發子提供了重要參考。相比于蟲口密度的難以精確控制,茶小綠葉蟬激發子具有安全有效和精確控制的特點,未來對茶葉品質提升應用具有巨大的潛力。
4.3 闡明茶小綠葉蟬侵害對茶葉品質和產量影響的內在機制與品質-產量調控的重要節點
品質與產量是衡量作物優良的兩個重要性狀。如前所述,茶小綠葉蟬侵害茶樹會誘導其產生豐富的次生代謝產物,這些代謝產物一方面幫助茶樹抵御昆蟲侵害,另一方面使茶葉品質得以提升。然而,和其他昆蟲侵害類似,茶小綠葉蟬侵害也會造成產量的大幅減產,嚴重時甚至導致茶葉絕收。因此,產量與品質的平衡,就是茶樹生長與代謝的平衡。目前茶小綠葉蟬侵害對茶葉品質和產量影響的研究大多在表型層面,其內在機制尚不清楚,未來的工作需要深入解析茶小綠葉蟬侵害對茶葉品質和產量影響的調控網絡,發掘關鍵基因。通過分析目前已報道的茶小綠葉蟬侵害茶樹轉錄組學數據,可發現植物激素信號轉導通路是不同茶小綠葉蟬處理下差異基因均富集的通路,因此是否有可能將來在該通路上尋找到茶小綠葉蟬調控品質與產量的關鍵節點。通過對該節點的精確調控,從而實現茶小綠葉蟬侵害下茶葉品質與產量的顯著性提升,實現“提質增產或者保產”的目標。
