基于管道模型理論的長白落葉松葉生物量與徑向生長關系研究
2022-03-25 08:47:00王軼夫喬晶晶邱思玉孫玉軍
林業科學研究 2022年2期
潘 磊,王軼夫,孫 釗,喬晶晶,邱思玉,孫玉軍
(北京林業大學森林資源和環境管理國家林業和草原局重點開放性實驗室,北京 100083)
葉生物量與光截獲、蒸騰作用、光合作用直接相關,是決定樹木生產力的最重要因素,為了間接估測葉生物量,Shinozaki 等提出了一種理論框架[1]:管道模型理論(pipe model theory),管道模型認為,在植物體內,單位數量的管道支持單位數量的葉子,管道不僅具有固定的橫截面積,而且通過所有的胞間層從葉子通過樹枝、樹干伸展到根尖部位,許多這樣的管道集合成束,形成一棵完整的樹形[2-4]。
根據管道模型理論,給定樹種的葉生物量應該與樹冠基部的樹干橫斷面積成一定比例[1,5],一些研究已經證明日本扁柏(Chamaecyparis obtusa(Siebold &Zucc.)Endl.)[6-7]、歐洲赤松(Pinus sylvestrisLinn.)[8]、白 樺(Betula platyphyllaSukaczev)[9]的葉生物量與枝下高樹干橫斷面積成比例。另外有研究表明,葉面積與樹干邊材面積成比例[10],然而這種比例并非是通用的,也有一些樹種存在例外[11-13],管道模型理論對于長白落葉松(Larix olgensisHenry)是否適用,是一個值得研究的問題。對于單株林木而言,關于樹葉在樹冠內的分布,先前的一些研究一般認為樹葉的分布在不同方向間沒有差異[14-16],然而在實地調查中發現,樹冠半徑和樹葉的分布在不同方向是有差異的。一些研究表明,分支大小和光合作用特征在不同水平方位是存在差異的,這種差異可能是由光環境的差異和不同方向競爭壓力大小的差異造成的[17-18]。
在樹干解析的過程中,發現樹干半徑和徑向生長量在不同方向上也存在差異,樹冠偏冠與樹干髓心偏心是生物量在樹冠和樹干上的不均勻分布,結合管道模型理論分析單木株內樹冠生物量在不同方位的不均勻分布與徑向生長周向變異的關系,有助于理解樹干生長和樹冠結構之間的功能關系,對于理解樹木生長發育等生態過程具有重要的意義。
1 研究方法與材料
1.1 葉生物量測定
1.1.1 單木葉生物量測定 將解析木伐倒后,使用分層標準枝法按1 m 分層對標準木的葉生物量進行了測定。測定時,首先按1 m 對樹干進行分段,使用手鋸和枝剪將每段的枝條沿基部截下,現場使用電子秤稱取每個枝條的帶葉鮮質量,結合每段的平均枝長、平均基徑和平均鮮質量,在每段選取2~3 個標準枝,稱量其帶葉鮮質量,然后將標準枝上的枝與葉分離,并分別稱取枝和葉鮮質量,而后均勻選取50 g 左右的葉樣本,記錄葉樣本的鮮質量,將葉樣本帶回至實驗室,置于烘箱中在105 ℃恒溫下烘干2 天左右至恒質量,稱取記錄每個葉樣本的干質量即生物量,計算葉樣本的含水量,根據現場測定的每層葉鮮質量和該層葉樣本的含水率,計算每層的葉生物量,累加后即為該株解析木的葉生物量。
本研究在黑龍江省伊春市朗鄉林業局東折棱河林場共調查獲取了31 株不同林齡的長白落葉松解析木的葉生物量數據和徑向生長量數據,其基本統計指標如表1 所示。
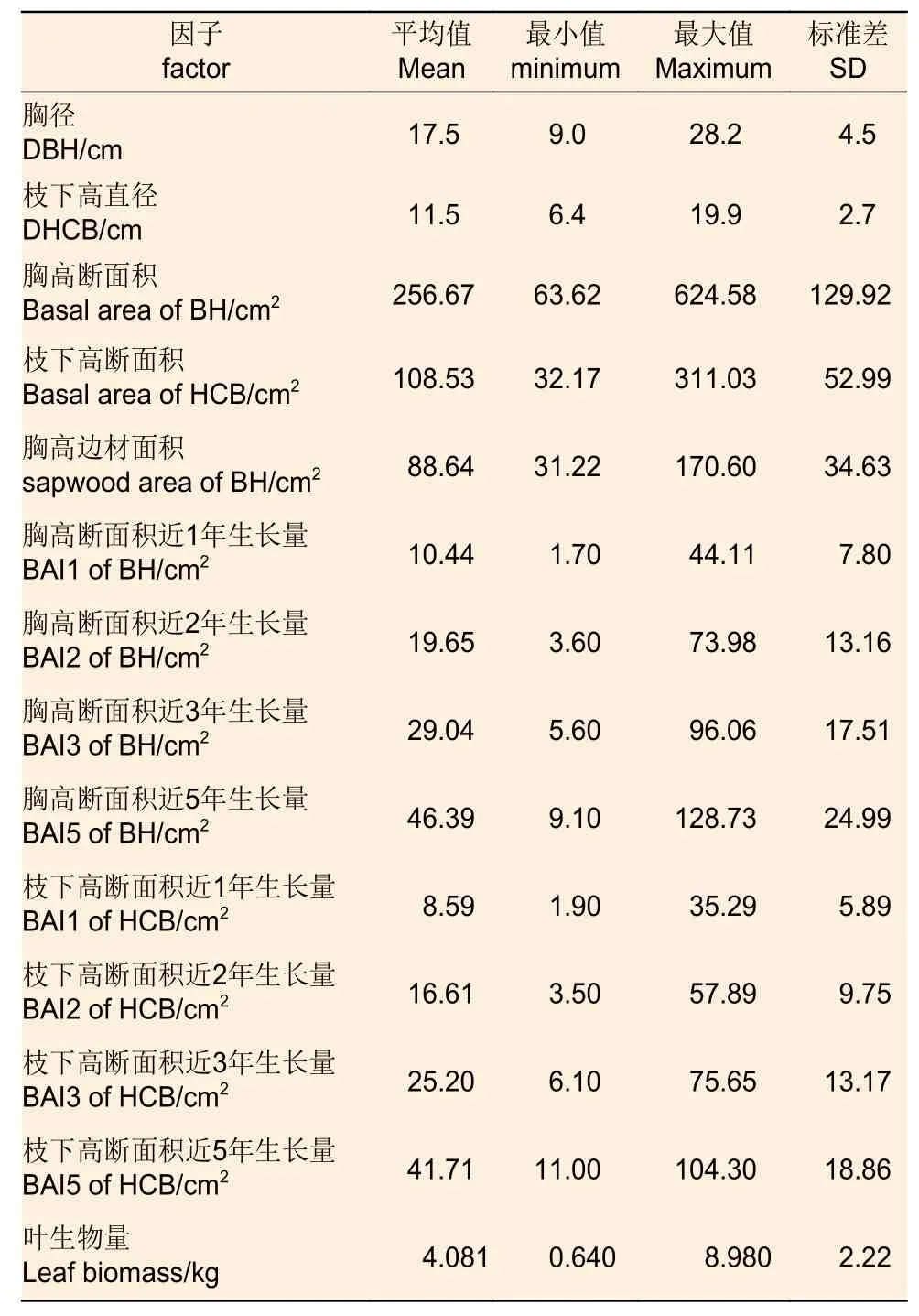
表1 建模生物量解析木的基本統計量Table 1 Descriptive statistics of sampled trees for biomass equations development
1.1.2 株內不同方位葉生物量測定 在單木株內水平上,將樹冠以45°間隔分為北(337.5°,22.5°)、東 北(22.5°,67.5°)、東(67.5°,112.5°)、東南(112.5°,157.5°)、南(157.5°,202.5°)、西南(202.5°,247.5°)、西 (247.5°,292.5°)、西 北(292.5°,337.5°) 8 個水平區間。
對標準木進行葉生物量測定時,使用數顯角度尺,以正北為0°,量測每個枝條的方位角,在各個區分段內,分別對8 個水平區間所有枝條進行稱質量。分別選取標準枝,將標準枝上的樹葉全部剝離稱質量,計算標準枝上樹葉的所占比例,而后收集50 g 左右的鮮葉樣品,將鮮葉樣本帶回至實驗室,置于烘箱中在105 ℃恒溫下烘干2 天左右至恒質量,稱取記錄每個葉樣本的干質量即生物量,計算葉樣本的含水量。根據各個區間的樹枝總質量、樹葉鮮質量占樹枝鮮質量比例、樹葉含水率,計算標準木各個方位區間的葉生物量。因為該項工作量較大,只對10 株解析木進行了株內不同方位的葉生物量調查,基本信息如表2 所示。
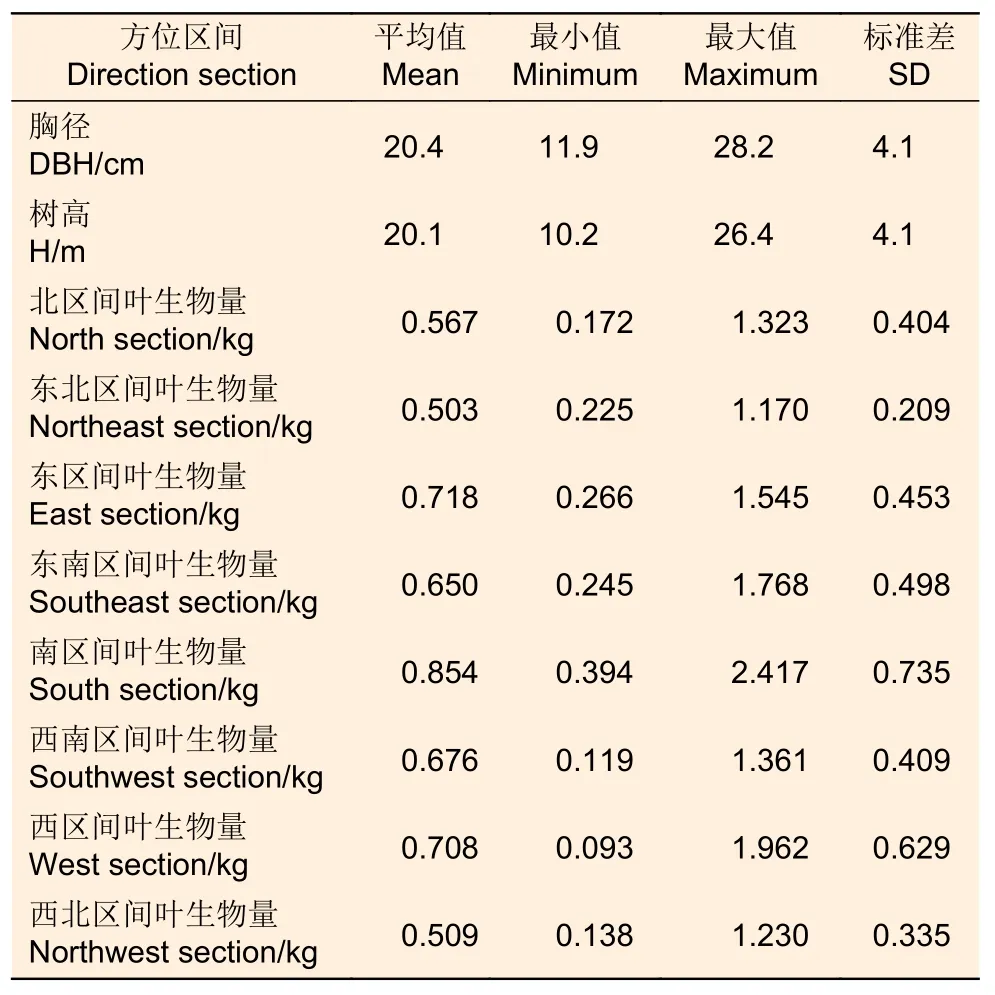
表2 株內不同方位區間的葉生物量Table 2 Descriptive statistics of sampled trees for leaf biomass in different direction
1.2 徑向生長測量
將標準木伐倒后,截取胸高處和枝下高處的圓盤,并在工作面上表明正北方向,將圓盤帶回實驗室,使用角磨機和砂紙將圓盤工作面打磨拋光至年輪清晰可見,并在工作面上使用鉛筆標明北、東北、東、東南、南、西南、西、西北8 個方向,而后將圓盤放在掃描儀上使用1200dpi 的分辨率掃描成jpg 格式的柵格圖像,利用WinDENDRO 年輪分析系統測定圓盤上8 個方向樹干半徑上的年輪寬度,測定精度為0.01 mm,以此獲得圓盤上8 個樹干半徑的長度和各個方向上的年輪寬度。
分別以8 個方向上的樹干半徑為半徑計算1/8的圓面積,將8 個樹干半徑計算的1/8 圓面積的和視為該高度的樹干斷面積。與上述10 株解析木不同方位區間葉生物量對應的不同方位區間樹干半徑、斷面積和斷面積生長量的基本信息如表3 所示。
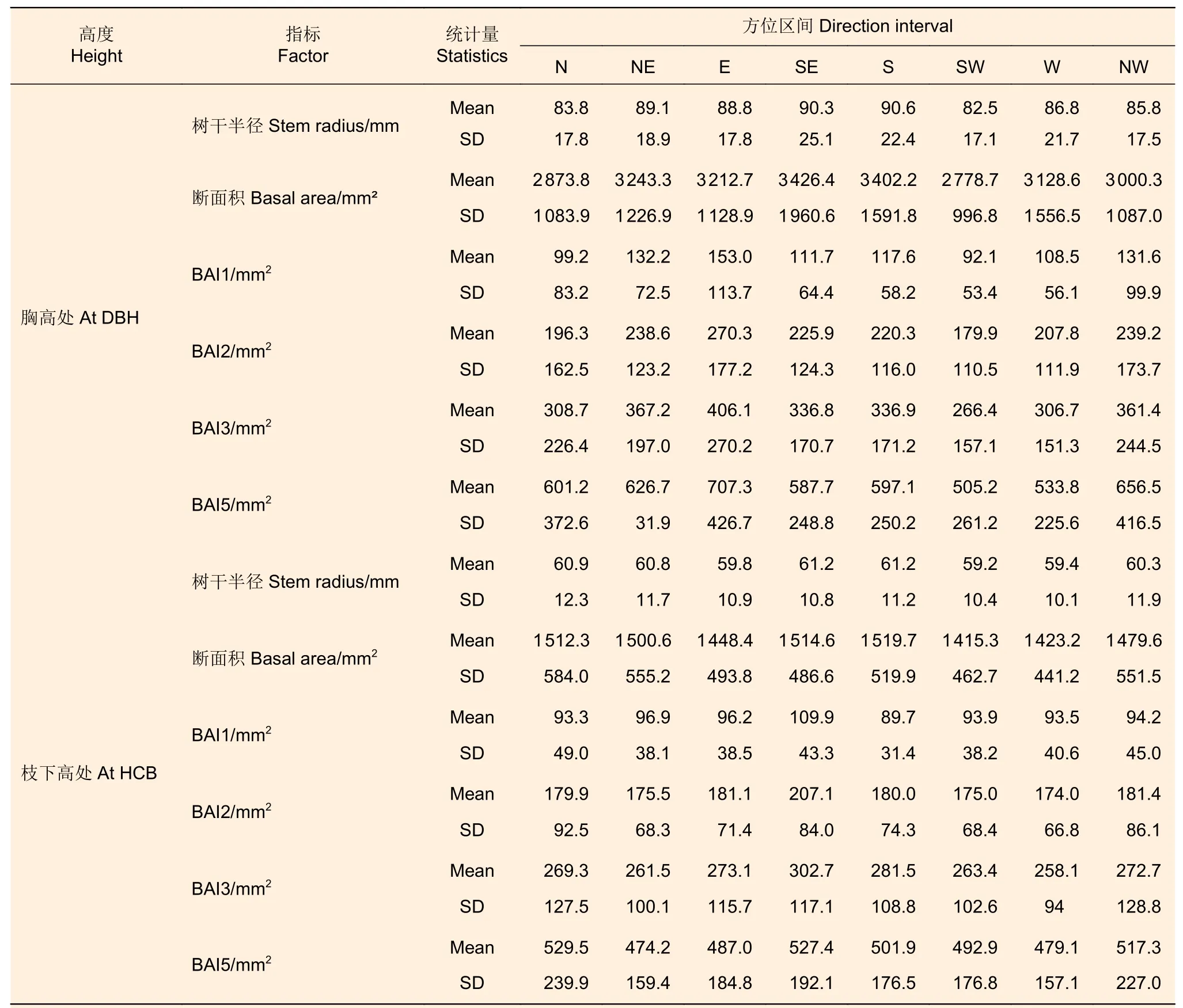
表3 株內不同方位區間的斷面積及生長量Table 3 Basal area and increment of different directions within tree
在胸高處圓盤上以顏色區分出心材和邊材的界限,分別量測8 個方向上的心材半徑,使用上述計算樹干斷面積的方法,分別計算得出8 個區間的心材面積,使用各個方向上對應的樹干斷面積減去心材面積,即得出各個方向上的邊材面積,將8 個方向上的邊材面積相加即為胸高處邊材面積,由于在枝下高處圓盤上心材和邊材的顏色界限不明顯,不能根據顏色進行辨別,故只對胸高處的邊材面積進行了測量計算。
1.3 統計分析
結合管道模型理論,使用一元線性回歸模型分析了胸高處和枝下高處的直徑、斷面積、胸高邊材面積和近n年斷面積生長量的關系;由于樹木存在樹冠偏冠和樹干髓心偏心現象,為了探討這兩種現象是否存在關系,結合管道模型理論,分別以長白落葉松株內不同方位區間的葉生物量和對應方向區間的胸高處和枝下高處的樹干半徑、斷面積和斷面積近n年生長量進行相關分析。數據處理分析、圖形繪制均在R 3.6.5 中進行。
調查的上海環城綠帶植物群落的喬木層的 Shannon-Wiener 指數在 0~1.572之間,平均為0.750。群落的物種多樣性指數整體偏小,喬灌草各層物種多樣性變化沒有明顯規律,不同群落間的物種多樣性有明顯差異。該結果反映出百米林帶植物群落在最初進行設計時,對群落的生態效應和穩定性未作充分考慮,從而表現出群落豐富度不夠。
2 結果與分析
2.1 單木葉生物量與徑向生長的關系
2.1.1 單木葉生物量與樹干直徑的關系 根據管道模型理論的基本原理,單株樹木的葉生物量與樹干斷面積成一定比例關系。本研究繪制了單木葉生物量與胸徑、胸高斷面積、枝下高直徑、枝下高斷面積的散點圖,并根據散點分布趨勢和生物量原理建立了y=ax的線性關系(圖1、圖2)。
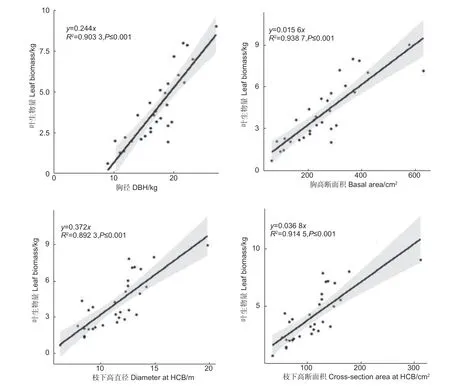
圖1 單株葉生物量與直徑、斷面積的關系Fig.1 The relationship between leaf biomass and diameter and basal area
由圖1 可知,單木胸徑和胸高斷面積與單木葉生物量均成明顯的線性關系,兩者的決定系數均達到了0.90 以上,其中胸高斷面積的決定系數略高于胸徑的決定系數。單木枝下高直徑和枝下高斷面積與單木葉生物量同樣均成明顯的線性關系,兩者的決定系數均達到了0.89 以上,其中枝下高斷面積的決定系數略高于枝下高直徑的決定系數。
由圖2 可知,胸高邊材面積與葉生物量也呈現明顯的線性關系,其決定系數達到0.863 5,說明胸高邊材與葉生物量存在較好的相關性,不過略低于胸徑、胸高斷面積、枝下高直徑、枝下高斷面積的決定系數。本研究中邊材是通過顏色差異和心材進行區別的,可能會存在測量誤差。根據管道模型理論,樹干中任何部位的邊材均應與其上方的葉生物量具有良好的相關關系,該結果符合管道模型理論。
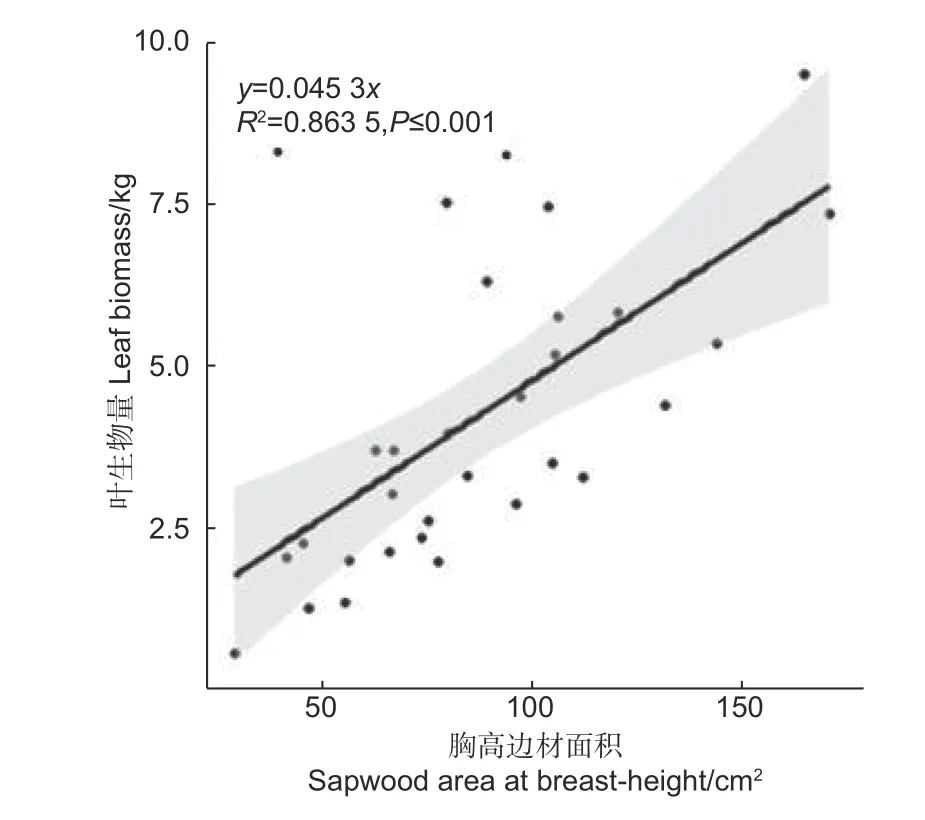
圖2 單株葉生物量與胸高邊材面積的關系Fig.2 The relationship between leaf biomass and sapwood basal area at breast height
2.1.2 單木葉生物量與樹干斷面積生長量的關系
對于單木而言,樹葉與樹干是相互依賴,相互制約的,葉量的多少受到樹干運輸水分和礦物質能力大小的制約,樹干的生長快慢又受到樹葉光合作用合成有機物多少的制約[19]。而在樹干中,只有邊材部分具有輸導能力,然而邊材與心材的分別一般以顏色來區分,在顏色不明顯時,在邊材與心材的交界處可能存在辨別誤差,并且有學者表明,對于針葉樹而言,邊材部分靠近心材的部分年輪已不再具備輸導能力,樹干最外圍的一兩個年輪起到的輸導作用最大[20]。為此,本研究建立了葉生物量與胸高處、枝下高處最近1,2,3,5 年斷面積生長量之間y=a+bx的線性關系(如圖3)。
由圖3 可知,胸高斷面積近1、2、3、5 年生長量均與葉生物量呈顯著正相關,決定系數在0.64和0.79 之間,其中近5 年生長量的決定系數最大為0.786 1,然后依次是近3 年生長量(0.744 5),近2 年生長量(0.704 5),近1 年生長量(0.646 2)。枝下高斷面積近1、2、3、5 年生長量也均與葉生物量呈顯著正相關,決定系數在0.72 和0.78 之間,其中近5 年生長量的決定系數最大為0.784 1,然后依次是近3 年生長量(0.755 3),近2 年生長量(0.733 7),近1 年生長量(0.720 1),略高于胸高處斷面積生長量的決定系數。
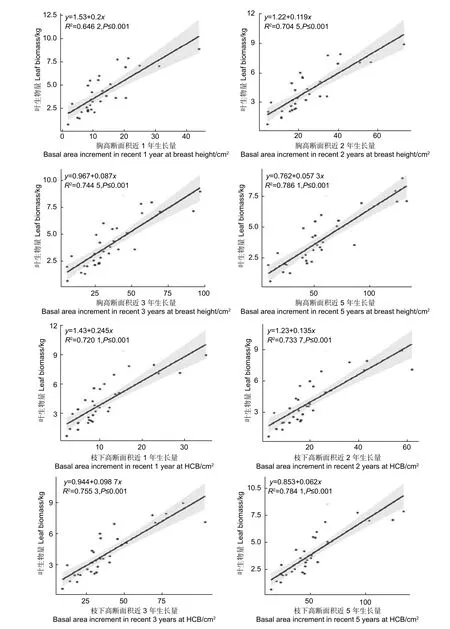
圖3 單株葉生物量與胸高、枝下高斷面積近1 年、2 年、3 年、5 年生長量的關系Fig.3 The relationship between leaf biomass and basal area increment in recent 1, 2, 3, 5 years at breast height and height to crown base
2.2 株內不同方位葉生物量與徑向生長的關系
由表4 可知,株內不同方位葉生物量與胸高處和枝下高處的半徑、斷面積和近1、2、3、5 年斷面積生長量均呈顯著正相關。總體而言,對應方位樹干半徑、斷面積的相關系數要高于近1、2、3、5 年斷面積生長量的相關系數。枝下高處樹干半徑、斷面積的相關系數要高于胸高處的,而對于斷面積生長量而言,胸高處的相關系數均高于枝下高處的相關系數。
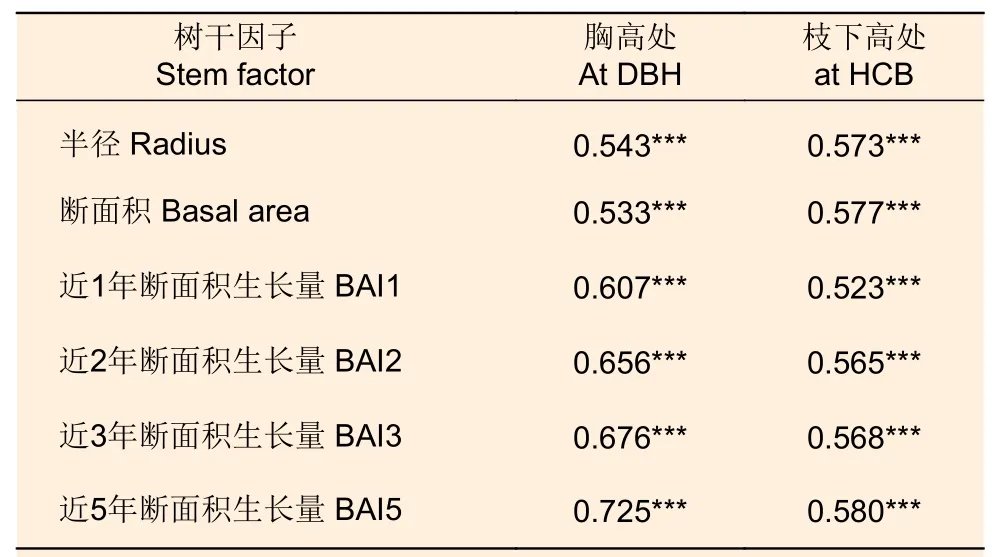
表4 株內不同方位葉生物量與對應方位樹干半徑、斷面積和斷面積生長量的相關系數Table 4 Correlation coefficients between leaf biomass in different directions and stem radius, basal area and basal area increment in corresponding directions
2.2.1 不同方位區間葉生物量與樹干半徑的關系
由圖4 可知,不同方位區間胸高半徑和胸高斷面積與對應方位區間的葉生物量均成明顯的線性關系,兩者的決定系數均達到了0.70 左右,其中胸高斷面積的決定系數略高于胸徑的決定系數。不同方位區間枝下高半徑徑和枝下高斷面積與對應方位區間葉生物量同樣均成明顯的線性關系,兩者的決定系數均達到了0.70 以上,其中枝下高斷面積的決定系數略高于枝下高半徑的決定系數。
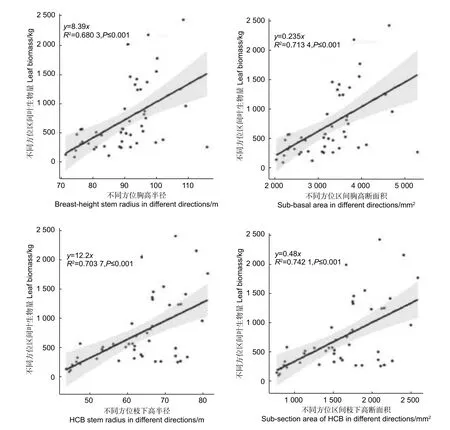
圖4 株內不同方位區間葉生物量與對應區間半徑、斷面積的關系Fig.4 The relationship between leaf biomass and radius and basal area of different orientations
2.2.2 不同方位區間葉生物量與樹干斷面積生長量的關系 為了探究樹冠的葉生物量分布與徑向生長量的關系,建立了不同方位區間葉生物量與對應方位區間胸高處、枝下高處最近1,2,3,5 年斷面積生長量之間y=a+bx的線性關系(圖5)。
由圖5 可知,不同方位區間胸高斷面積近1、2、3、5 年生長量均與對應區間葉生物量呈顯著正相關,決定系數在0.36 和0.52 之間,其中近5 年生長量的決定系數最大為0.525 5,然后依次是近3 年,近2 年,近1 年生長量。不同方位區間枝下高斷面積近1、2、3、5 年生長量也均與對應方位區間葉生物量呈顯著正相關,決定系數在0.27 和0.34 之間,其中近5 年生長量的決定系數最大為0.335 9,然后依次是近3 年,近2 年,近1 年生長量,低于不同方位區間胸高處斷面積生長量與葉生物量的相關關系。
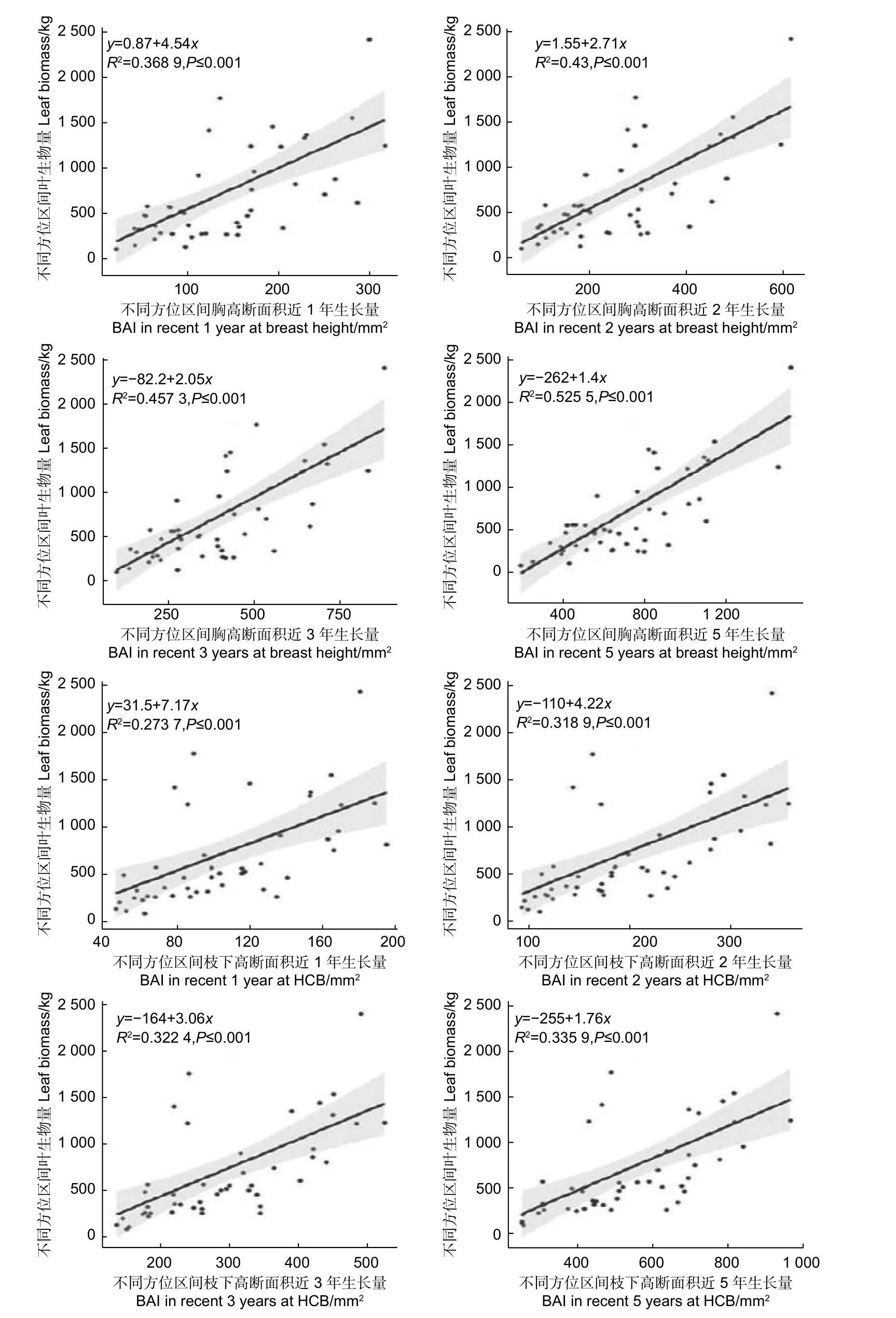
圖5 不同方位葉生物量與胸高、枝下高斷面積近1,2,3,5 年生長量的關系Fig.5 The relationship between leaf biomass and basal area increment in recent 1, 2, 3, 5 years at breast height and height to crown base
3 討論
劉盛等[21]使用生物染色劑對長白落葉松的水分輸導模式進行了研究,結果表明長白落葉松枝干中的水分輸導模式符合管道模型理論,水分在枝干的不同年輪間沒有明顯的橫向輸導現象,在同一年輪內,水分的向上傳輸呈現較好的定向傳輸性,并不是均衡向上傳輸,說明某一枝條上的樹葉僅與某些導管相連接,從樹葉基部通過樹干中的導管貫通至根系。
Shinozaki 等[22]在他們關于管道模型理論的第二篇論文中使用實驗數據進一步驗證了該理論,并得出結論“作為該理論在森林生態學中的應用,根據葉生物量和枝下高樹干橫截面積之間的比例關系,可以估算樹木和林分的葉生物量,”盡管該方法比Joseph[23]提出的使用胸徑的方法更精確,但是并未得到廣泛使用,有以下幾個原因,首先,該比例不僅取決于樹種,還取決于生長條件和林木發育階段,其次,次生生長是一個不可逆的累積過程,樹干直徑可以增長或停滯,但是不會減少,而樹木的葉生物量或葉面積可能會增加或減少,可能會出現季節性波動[24],或因為競爭、衰老、機械損傷而減少[25]。最后,基于管道模型理論的葉生物量估計需要測量到枝下高處,并在不同方向獲取樣芯,以推測計算枝下高處的邊材面積,而這些測量工作都比只測量胸徑要繁瑣復雜。一些研究者調查了胸高處邊材面積和枝下高處邊材面積與胸徑和枝下高直徑在估測葉生物量上的相關程度,并對結果進行了對比。以胸徑范圍在13~43 cm,年齡在15~55 a 之間的火炬松為例,使用胸徑估測葉生物量的效果等于或優于胸高處邊材面積和枝下高處邊材面積[26]。Snell and Brown[27]對7 個針葉樹種的研究表明,對于其中3 個樹種,邊材面積對于葉生物量和枝生物量的估測效果要優于胸徑,而對于其他4 個樹種,邊材面積和胸徑對于葉生物量的估測效果沒有顯著差異。因此,可能為了便于野外測量,在大多數研究工作中,樹干直徑仍然是用來估計葉生物量的首選方法。
樹冠形狀和葉生物量分布與樹干的徑向生長緊密相關,并且會影響光合產物在樹干中的分布[28-29],由于葉生物量在周向上的不均勻分布,樹干上常常會伴隨產生與木材力學性能相關的偏心徑向生長[30]。
導致樹冠偏冠和葉生物量不均勻分布的一個外因是鄰近競爭。在本研究中,長白落葉松不僅存在不同程度的樹冠偏冠,在樹干中也存在著一定程度的偏心徑向生長。通過分析長白落葉松株內不同方位葉生物量與對應方位胸高處和枝下高處半徑和徑向生長量的關系,結果表明,株內不同方位葉生物量分布與對應方位徑向生長量呈顯著正相關,也就意味著上方樹冠的分布越多,下方樹干的徑向生長也就越快。較大的冠形意味著更大的樹冠生物量和葉生物量,這可能是對應徑向生長相較更快的原因[31-32]。
4 結論
本研究基于管道模型理論,使用樹冠的葉生物量和樹干直徑、斷面積、斷面積生長量等實測數據,在單木水平上擬合了葉生物量估測模型,對比了使用胸高處和枝下高處樹干直徑、斷面積、邊材面積和近1、2、3、5 年斷面積生長量估測單木葉生物量的效果,結果表明胸高斷面積作為預測因子的擬合效果最好。由于樹冠偏冠現象和樹干髓心偏心現象的存在,基于管道模型理論的擴展,本研究將單木根據方位分為8 個均等區域,分別調查了各區域的葉生物量和對應的胸高處和枝下高處的樹干半徑、斷面積和近1、2、3、5 年斷面積生長量,通過與葉生物量的相關分析表明,單木株內不同方位區域的葉生物量與對應的胸徑處和枝下高處的樹干半徑、斷面積和近1、2、3、5 年斷面積生長量均呈顯著正相關。本研究以長白落葉松葉生物量和徑向生長相關因子為研究對象,結合管道模型理論,初步分析了長白落葉松偏冠現象與樹干髓心偏心現象之間的關系,不過本研究只是進行了初步的統計分析,而未更深層次的結合樹木生理學進行研究分析,對樹冠偏冠和髓心偏心關系的確定還需今后進一步的研究。
