轉錄因子GATA結合蛋白3對哮喘易感基因ADAM33表達的影響
2022-03-28 02:49:22芮菲菲杜娟郝忠分馬世奇王雯束進
江蘇大學學報(醫(yī)學版) 2022年2期
芮菲菲, 杜娟, 郝忠分, 馬世奇, 王雯, 束進
(江蘇大學第四附屬醫(yī)院兒科, 江蘇 鎮(zhèn)江 212001)
隨著分子生物學和分子遺傳學發(fā)展,近來證實哮喘是一種多基因遺傳病[1]。在眾多哮喘易感基因中,第一個通過定位克隆鑒定發(fā)現(xiàn)與哮喘有關的易感基因為解整合素-金屬蛋白酶33(a disintegrin and metalloproteinase 33,ADAM33)[2],其位于人類染色體20p13,含有22個外顯子和21個內(nèi)含子,編碼膜錨定酶去整合素和金屬蛋白酶結構域蛋白33,影響參與氣道重塑的生長因子和細胞因子活性[3],從而誘導哮喘發(fā)作。
研究發(fā)現(xiàn),GATA結合蛋白3(GATA binding protein 3,GATA3)在哮喘患者的支氣管肺泡灌洗液及外周血單個核細胞中呈高表達[4]。全基因組關聯(lián)分析發(fā)現(xiàn),GATA3單核苷酸多態(tài)性位點rs10905284是哮喘發(fā)病的重要危險因素[5]。由此可見,GATA3表達與哮喘的發(fā)生發(fā)展密切相關。GATA3可作為調節(jié)T細胞分化為Th2細胞的關鍵轉錄因子,其表達上調激活氣道和小氣道黏膜Th2細胞,引起一系列細胞炎性因子(IL-4、IL-5、IL-9及IL-13)釋放[6],從而導致哮喘發(fā)作。ADAM33與血清IL-4、IL-1含量呈正相關,其表達可能受Th2型細胞因子調控[7],進而誘導哮喘的發(fā)生發(fā)展。由此推測,轉錄因子GATA3可能通過與ADAM33啟動子結合影響ADAM33表達從而引起哮喘發(fā)作。因此本研究以人HEK293T、BEAS-2B細胞為研究對象,探討GATA3對ADAM33轉錄調控的影響。
1 材料與方法
1.1 細胞和主要試劑
人腎臟HEK293T細胞和人正常肺上皮BEAS-2B細胞購自美國典型培養(yǎng)物保藏中心;DMEM高糖培養(yǎng)基、胎牛血清(美國Gibco公司);KpnⅠ和BglⅡ酶(美國NEB公司);雙熒光素酶試劑盒(美國Promega公司);DNA高保真酶、DNA DL5000及DL2000標準參照物、實時熒光定量PCR試劑盒(日本TaKaRa公司);T4DNA連接酶、GenGreen核酸染料(美國Thermo Fisher公司);感受態(tài)大腸埃希菌DH5α、PCR引物、轉染試劑Lipofectamine 3000、pcDNA-GATA3質粒及pcDNA3.1質粒、RNA 提 取 試 劑Trizol均為美國Invitrogen公司產(chǎn)品;瓊脂糖粉(西班牙Biowest公司);膠回收試劑盒、質粒提取試劑盒(美國Omega公司)。
1.2 細胞培養(yǎng)
將人HEK293T、BEAS-2B細胞用含10%胎牛血清、100 U/mL青霉素、100 μg/mL鏈霉素的高糖DMEM,于37 ℃、5% CO2培養(yǎng)箱進行培養(yǎng)。
1.3 重組報告質粒的構建與鑒定
以GenBank數(shù)據(jù)庫中的人ADAM33基因(轉錄本2,NM-153202)mRNA序列為模板,用Primer 5.0設計特異性引物,在上、下游引物5′端分別加入KpnⅠ和BglⅡ酶切位點,酶切位點前加保護堿基,上游引物分別為P1:5′-CGGGGTACCGCCTGGCCAAG-ACCAGGAGTTTGAG-3′;P2:5′-CGGGGTACCAGA-ATCGCTTGAACCAGGGAGTCGG-3′;P3:5′-CGGGGTACCCGGCCTTTGCCCACCGCCTCATCCC-3′;P4:5′-CGGGGTACCGGAGTGGCGACCCGGCTCCCCCATG-3′;P5:5′-CGGGGTACCGGAGGCGGGAGCGAGGGCGG-GCGGT-3′;下劃線部分為KpnⅠ酶切位點;共用下游引物:5′-GGAAGATCTGCTGGATTGCGGCCGACCCGTGCC-3′(下劃線部分為BglⅡ酶切位點)。以BEAS-2B細胞的DNA為模板,行PCR擴增,反應體系:DNA聚合酶 0.5 μL,5×緩沖液5 μL,dNTP混合物2 μL,cDNA 0.5 μL,上游引物0.5 μL,下游引物0.5 μL,雙蒸水16 μL,共25 μL。擴增條件:98 ℃ 10 s,60 ℃ 15 s,68 ℃ 1 min,30個循環(huán);4 ℃保存。擴增產(chǎn)物進行電泳;回收目的片段條帶并進行純化;將純化產(chǎn)物及報告基因質粒pGL3-Basic行KpnⅠ和BglⅡ雙酶切、電泳、純化;用T4DNA連接酶將純化的目的基因與pGL3-Basic載體連接,并轉化至感受態(tài)大腸埃希菌DH5α,培養(yǎng)過夜;選取若干個菌落加到培養(yǎng)基中混勻,取適量菌液為模板,PCR擴增后電泳,將出現(xiàn)目的條帶的菌液送至測序公司進行測序,再將測序正確的菌液取適量振蕩培養(yǎng)過夜后進行質粒的小量制備,構建P1、P2、P3、P4、P5等不同長度的ADAM33質粒。
1.4 重組質粒中ADAM33基因啟動子活性鑒定
將人HEK293T、BEAS-2B細胞按構建的ADAM33質粒分為P1、P2、P3、P4、P5組及pGL3-Basic組。將細胞按每孔1.5×104個細胞密度接種于96孔板,每組3個復孔,培養(yǎng)24 h,用轉染試劑Lipofectamine 3000對其進行轉染。先配制a液:5 μL無血清培養(yǎng)基+0.2 μL Lipofectamine 3000;再配制b液:5 μL無血清培養(yǎng)基+100 ng不同的ADAM33質粒(P1、P2、P3、P4、P5)或100 ng pGL3-Basic質粒+4 ng pRL-TK質粒(為海腎熒光素酶報告載體,含有SV 40增強子,該質粒起內(nèi)參作用)+0.2 μL轉染試劑;然后,將a液與b液混勻,室溫放置5 min;最后,將混合液滴至孔內(nèi)孵育24 h;棄培養(yǎng)基,PBS洗滌;每孔分別加入細胞裂解液,搖床振蕩20 min將細胞充分裂解。按雙熒光素酶活性檢測試劑盒說明書對每孔進行加樣并充分混勻,然后測定熒光素酶活性(A值)和海腎熒光素酶活性(B值),計算熒光活性比值(A/B)分析重組質粒啟動子活性。重復3次實驗。
1.5 雙熒光素酶報告基因法檢測不同轉染細胞中ADAM33啟動子活性
將HEK293T、BEAS-2B細胞分別分為pcDNA-GATA3組和pcDNA3.1對照組,分別轉染GATA3過表達質粒和空白對照質粒。當細胞融合達60%左右時,按每孔1.5×104個細胞密度接種于96孔板,每組3個復孔,用轉染試劑Lipofectamine 3000轉染HEK293T、BEAS-2B細胞,轉染方法及熒光素酶活性檢測同“1.4”,實驗重復3次。
1.6 實時熒光定量PCR檢測ADAM33 mRNA表達
取“1.5”分組細胞,當BEAS-2B細胞融合60%左右時,按每孔1.5×105個細胞密度接種于12孔板,用轉染試劑Lipofectamine 3000對BEAS-2B細胞進行轉染。轉染方法同“1.4”。
用Trizol試劑提取轉染后BEAS-2B細胞總RNA,并逆轉錄為cDNA。逆轉錄反應體系:5×PrimeScript RT Master 2 μL,RNA樣品和無RNA酶雙蒸水8 μL,總反應體系為10 μL。逆轉錄反應條件:37 ℃ 15 min,85 ℃ 5 s,4 ℃保存。實時熒光定量PCR反應體系:8.2 μL DEPC水、10 μL SYBR Premix ExTaqⅡ和上、下游引物各0.4 μL、1 μL cDNA;擴增條件:95 ℃預變性3 min,95 ℃變性5 s,56 ℃退火30 s,循環(huán)42次;熔解:95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。ADAM33mRNA引物序列:上游為5′-CTGGGTCCTGGATGGACAAC-3′,下游為5′-CAGCAGCCTGTGGTTCTTCT-3′。以GAPDH為內(nèi)參基因,對實驗結果數(shù)據(jù)進行分析:mRNA相對表達量=2-ΔΔCt;ΔCt=Ct目的基因-Ct內(nèi)參基因;ΔΔCt=ΔCt實驗組-ΔCt對照組。設對照組mRNA相對表達量為1,實驗重復3次。
1.7 統(tǒng)計學分析
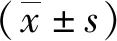
2 結果
2.1 ADAM33基因5′側翼區(qū)重組質粒的構建
以BEAS-2B細胞基因組的DNA為模板經(jīng)PCR擴增ADAM335′側翼區(qū)的基因序列,電泳結果顯示有特異性條帶出現(xiàn),長度約為1 953 bp,與預測片段長度相符(圖1A)。重組質粒經(jīng)雙酶切、電泳出現(xiàn)2條條帶(圖1B),其中一條約為4 818 bp,另一條約為1 953 bp,表明目的片段已插入pGL3-Basic載體。重組質粒經(jīng)測序后行Blast比對,結果顯示,其與GenBank中登記的ADAM33基因序列相同,表明含ADAM33基因啟動子的質粒構建成功。重組的5′側翼區(qū)不同長度的ADAM33截短質粒經(jīng)雙酶切,電泳出現(xiàn)2條條帶,一條約為4 818 bp,另一條分別約為1 953 bp、853 bp、363 bp、174 bp、97 bp(圖1C)。將選用的GenBank數(shù)據(jù)庫中ADAM33mRNA序列5′末端為轉錄起始位點定義為+1,構建的質粒分別命名為P1:pGL3-1880/+73;P2:pGL3-780/+73;P3:pGL3-290/+73;P4:pGL3-101/+73;P5:pGL3-24/+73。經(jīng)外送測序及Blast比對,證實重組的不同長度的啟動子質粒序列與GenBank中登記的ADAM33基因序列相同。
2.2 ADAM33基因啟動子的活性鑒定
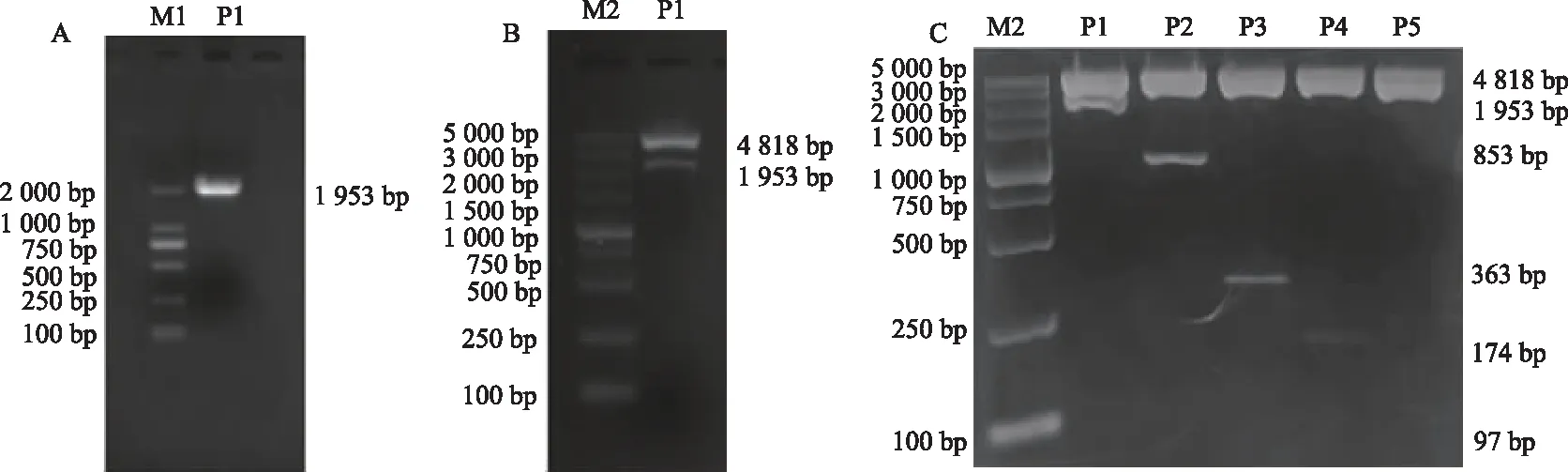
結果顯示,與pGL3-Basic組相比,HEK293T、BEAS-2B細胞中P1、P2、P3、P4、P5重組質粒熒光素酶活性明顯增高(P<0.05或P<0.01),其中,啟動子活性最高的均為P3質粒。當啟動子質粒進一步缺失至-24 bp時(P5),與pGL3-290/+73質粒(P3)相比,啟動子活性在HEK293T細胞和BEAS-2B 細胞中均降低,呈現(xiàn)大幅度下降(圖2),說明在-290 bp~-24 bp區(qū)域存在重要的正性調控元件。由此表明,ADAM33核心啟動子序列位于-290 bp~+73 bp。
M1: DNA2000標準參照物;M2: DNA5000標準參照物;P1: pGL-1880/+73;P2: pGL-780/+73;P3: pGL-290/+73;P4: pGL-101/+73;P5: pGL-24/+73;A: ADAM33基因5′側翼區(qū)序列PCR產(chǎn)物的瓊脂糖凝膠電泳結果;B: 含ADAM33基因啟動子的重組質粒的雙酶切結果;C:重組的截短質粒酶切鑒定結果
a: P<0.05,b: P<0.01,與pGL3-Basic組比較
2.3 GATA3上調ADAM33基因啟動子活性
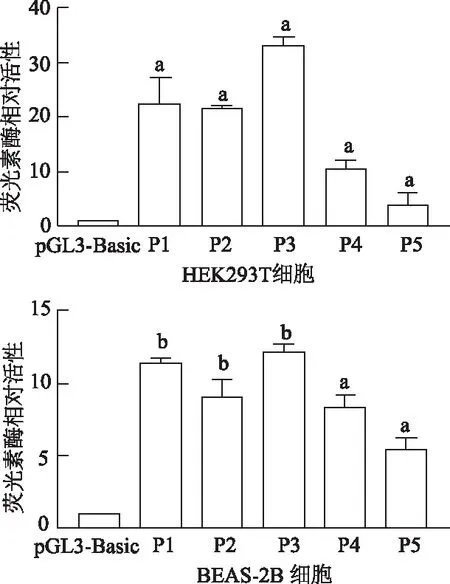
在HEK293T、BEAS-2B細胞中,與pcDNA3.1組相比,pcDNA-GATA3組ADAM33重組質粒pGL3-1880/+73的啟動子熒光素酶活性明顯增加(t=9.21,20.48,P<0.05或P<0.01),表明GATA3上調ADAM33基因啟動子活性。見圖3。
圖3 不同細胞中熒光素酶相對活性比較
2.4 GATA3上調ADAM33 mRNA表達
由圖4可見,與pcDNA3.1組相比,BEAS-2B細胞中pcDNA-GATA3組ADAM33mRNA相對表達量顯著增加(t=23.57,P<0.05),表明GATA3可上調ADAM33mRNA表達。
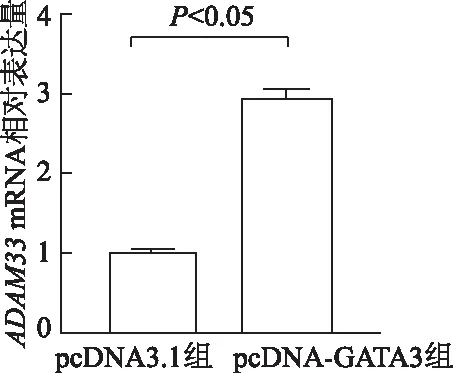
圖4 BEAS-2B細胞中ADAM33 mRNA相對表達水平比較
3 討論
本研究以GenBank數(shù)據(jù)庫中報道的人ADAM33基因(NM-153202)為模板,成功構建含人ADAM33啟動子的一系列截短重組質粒;并且發(fā)現(xiàn)在HEK293T、BEAS-2B細胞中,pGL3-290/+73質粒啟動子活性最高,ADAM33基因的核心啟動子序列位于該基因-290 bp~+73 bp。這與先前Yang等[8]研究所得到的結論相比,即ADAM33基因啟動子序列在-550 bp~+87 bp具有活性,進一步明確了ADAM33基因啟動子的核心序列所在位置。
本研究發(fā)現(xiàn)在HEK293T、BEAS-2B細胞中,過表達GATA3質粒可增強ADAM33啟動子活性。HEK293T細胞易被轉染,可作為功能細胞用于檢測ADAM33啟動子活性,但其為人體腎臟細胞系,不是肺組織相關細胞,利用其檢測過表達GATA3質粒對ADAM33mRNA的表達影響意義不大,故本研究僅在人正常肺上皮BEAS-2B細胞中檢測,結果發(fā)現(xiàn)過表達GATA3質粒可上調ADAM33mRNA表達。JASPAR網(wǎng)站(http://jaspar.genereg.net)預測表明,ADAM33基因的核心啟動子區(qū)域含有GATA3、EGR1、SOX18、KLF5和E2F1等轉錄因子結合位點。本研究結果提示轉錄因子GATA3通過與ADAM33啟動子結合增加ADAM33表達,從而可能誘導哮喘發(fā)生。這與既往研究[9-10]中GATA3能夠激活IL-4、IL-13基因的啟動子,上調二者表達相一致。此外,有研究發(fā)現(xiàn),通過T-bet/GATA-3途徑調節(jié)相關細胞因子(IL-4、IL-5、IL-9及IL-13)水平和Th1/Th2比值可減輕哮喘大鼠氣道炎癥及減輕哮喘癥狀[11]。由此,本實驗結果進一步表明GATA3與哮喘發(fā)生密切相關。
綜上所述,本研究表明,轉錄因子GATA3與ADAM33基因的核心啟動子區(qū)域結合,上調人哮喘易感基因ADAM33啟動子活性及mRNA表達,但具體與哪個區(qū)域結合,還需后續(xù)行染色質免疫沉淀和凝膠遷移等實驗進一步研究。
