紅景天苷抑制破骨細胞分化和極化的初步研究
2022-04-02 07:57:06易清清梁鵬晨孫苗苗楊榮梁冬雨沙爽常慶
天津醫藥 2022年3期
易清清,梁鵬晨,孫苗苗,楊榮,梁冬雨,沙爽,常慶△
骨是一種礦化的結締組織,破骨細胞和成骨細胞之間的信號傳導在骨重塑和維持骨微環境穩態中起著重要作用[1]。骨質疏松癥(osteoporosis,OP)是一種以骨量降低與骨微結構退化為特點的疾病,骨折是其最常見的并發癥[2]。破骨細胞來源于造血干細胞,是由單核細胞融合分化而成,是唯一具有骨吸收功能的細胞。破骨細胞通過分泌組織蛋白酶K(CK)和基質金屬蛋白酶-9(MMP-9)降解骨基質[3]。破骨細胞與骨組織接觸的細胞膜區域具有特殊的黏附結構,形成封閉區,其中的細胞膜具有微絨毛結構樣褶皺緣,兩者形成相對獨立的骨吸收間隙,從而完成細胞極化[4]。整合素(Integrin)能調節黏附、遷移及封閉區的形成,對破骨細胞分化和激活具有重要意義[5-6]。非受體酪氨酸激酶(c-Src)及其磷酸化產物p-Src 調節細胞極化、遷移及褶皺緣的形成,敲除c-Src的小鼠因體內破骨細胞無法形成正常偽足,導致產生骨硬化癥[7-8]。紅景天苷(Salidroside,SAL)為4-羥基-苯基-β-D-吡喃葡萄糖苷,是紅景天屬植物中廣泛存在的酚苷類化合物,可從植物根、莖提取[9]。SAL具有保護心腦血管、調節免疫、抗腫瘤、促成骨等藥理作用[10]。近年來研究表明,SAL 可促進骨折后的骨再生和血管生成[11]。有關SAL促成骨的研究多集中于促進成骨細胞和內皮細胞活性方面,關于SAL 和破骨細胞分化及極化關系的研究尚少見報道。本實驗旨在研究SAL對可溶性核因子κB受體活化因子配體(sRANKL)誘導的破骨細胞分化和極化的影響,以期為SAL的臨床應用提供理論依據。
1 材料與方法
1.1 材料 SAL購自成都瑞芬思生物科技公司,小鼠單核細胞RAW264.7 細胞株購自蘇州賽爾飛生物科技公司,高糖DMEM 培養液、胎牛血清(FBS)均購自美國Gibco 公司,重組小鼠sRANKL購自上海賽戈生物科技公司,抗酒石酸酸性磷酸酶(TRAP)染色試劑盒購自北京索萊寶公司,鬼筆環肽購自美國Sigma公司,DAPI和茜素紅染色液購自上海碧云天生物技術公司,兔源MMP-9、c-Src、CK、Integrin β3一抗及羊抗兔二抗購自美國Cell Signaling 公司,實時熒光定量逆轉錄聚合酶鏈反應(qPCR)引物購自上海捷瑞生物公司,RT-PCR試劑盒購自北京Takara公司。
1.2 方法
1.2.1 細胞培養 小鼠RAW264.7 細胞系用含10% FBS 的DMEM培養基于37 ℃、5%CO2條件下培養,隔天換液,細胞密度80%以上時于6 孔板中鋪板培養。隨后細胞用含10%FBS,100μg∕L sRANKL 的DMEM 培養基繼續培養5 d。對照組不含SAL 的培養液,實驗組分別加入含15、30 和60 mg∕L SAL 的培養液,于37 ℃、5%CO2條件下培養,隔日換液,連續培養5 d。本文中SAL 的質量濃度15、30、60 mg∕L 均為加到細胞培養基中的終濃度。
1.2.2 TRAP 染色 細胞培養5 d 后,棄培養基,PBS 洗2 次,每孔加入500 μL 4%多聚甲醛固定30 min,PBS 洗2 次,按TRAP 染色試劑盒說明書進行染色,顯微鏡下觀察,對TRAP染色陽性細胞進行計數。
1.2.3 纖維形肌動蛋白(F-Actin)環染色 將生長狀態良好的RAW264.7細胞系以1×105個∕孔的密度接種于6孔板中,細胞貼壁后分別更換含100μg∕L sRANKL 及0、15、30 和60 mg∕L SAL的DMEM培養基,于37 ℃、5%CO2環境下繼續培養。培養48 h,棄培養基,PBS洗2次,4%多聚甲醛固定30 min,PBS洗2次,加入5 mg∕L 鬼筆環肽染色液,37 ℃避光孵育30 min,棄染色液,PBS洗2次,加入DAPI染色液染色5 min,PBS洗2次。顯微鏡下觀察拍照,計數F-Actin環形成數。
1.2.4 茜素紅染色 細胞以1×105個∕孔接種于6 孔板中,貼壁后分別換含100μg∕L sRANKL、成骨誘導液及0、15、30 和60 mg∕L SAL 的DMEM 培養基,于37 ℃、5%CO2環境下繼續培養。培養14 d 后,吸除培養基,并用PBS 沖洗細胞,4%多聚甲醛中固定30 min,用茜素紅染色30 min,顯微鏡下觀察破骨細胞鈣化情況。顯微鏡拍攝后,用10%十六烷基氯化吡啶溶解,562 nm處測定光密度(OD)值。
1.2.5 qPCR 檢測MMP-9、c-Src、CK和Integrin β3的mRNA表達 0、15、30和60 mg∕L SAL干預細胞5 d,使用Trizol法冰上抽提總RNA,根據試劑說明書進行RNA的逆轉錄及cDNA的擴增。逆轉錄條件:42 ℃15 min,85 ℃5 s。擴增條件:95 ℃10 s;95 ℃3 s,60 ℃30 s,72 ℃34 s,循環40 次。以GAPDH 為內參,目的基因相對表達量采用2-ΔΔCt法計算。引物序列見表1。

Tab.1 Amplification of gene primers by qPCR表1 qPCR擴增基因引物序列
1.2.6 Western blot 檢測MMP-9、c-Src 蛋白表達 0、15、30和60 mg∕L SAL 干預細胞5 d,在6 孔板中加入高效組織裂解液RIPA,冰上反復吹打,12 000 r∕min 離心10 min 后取上清液。上清液中加入上樣緩沖液,95 ℃加熱10 min 使蛋白變性,10%SDS-PAGE 分離蛋白后轉至PVDF 膜上,5%脫脂奶粉封閉抗原1 h,按說明書稀釋一抗MMP-9(1∶1 000)、c-Src(1∶1 000),內參GAPDH(1∶1 000),14 ℃孵育過夜,TBST 洗膜3 次,稀釋二抗(1∶20 000),室溫孵育1 h,TBST 洗膜3 次,加入ECL 化學發光液顯影曝光。根據灰度值數據分析各組MMP-9、c-Src蛋白相對表達量。
1.3 統計學方法 采用GraphPad Prism 5.0軟件進行數據分析,符合正態分布的計量資料采用均數±標準差(±s)表示,多組間均數比較采用單因素方差分析,組間多重比較采用Dunnet-t檢驗,體外細胞實驗結果均重復3次。P<0.05為差異有統計學意義。
2 結果
2.1 SAL抑制RAW 264.7分化成TRAP 陽性破骨細胞 在sRANKL誘導下,各組RAW264.7細胞均分化成破骨細胞,TRAP染色陽性證實sRANKL誘導分化成功。與對照組相比,SAL 15、30 和60 mg∕L SAL 組TRAP染色陽性破骨細胞的數目逐漸減少,見圖1。

Fig.1 The effect of different concentrations of salidroside on the count of osteoclasts(TRAP staining,×100)圖1 不同質量濃度SAL對破骨細胞計數結果的影響(TRAP染色,×100)
2.2 SAL 抑制RAW 264.7 分化破骨細胞F-Actin 環形成 對照組破骨細胞F-Actin環完整,SAL處理后,破骨細胞的F-Actin變細,甚至消失。隨著SAL質量濃度增高,破骨細胞的F-Actin 環破壞更加嚴重,FActin環數量減少,破骨細胞的數目亦越少,見圖2。
2.3 SAL促進破骨細胞鈣化形成 隨著SAL質量濃度增高,破骨細胞鈣結節茜素紅染色加深,SAL具有促進破骨細胞鈣化的作用,見圖3。
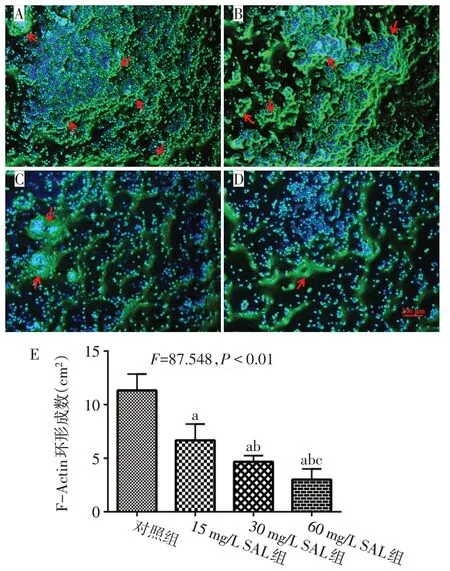
Fig.2 Effects of SAL on the formation of F-actin ring in osteoclasts圖2 不同質量濃度SAL對破骨細胞F-Actin環形成的影響
2.4 SAL 抑制MMP-9、c-Src、CK、Integrin β3mRNA的表達 與對照組相比較,30 和60 mg∕L SAL 組的MMP-9、CKmRNA,15、30 和60 mg∕L SAL 組的c-SrcmRNA,15、30 mg∕L SAL 組的Integrin β3mRNA 表達量降低(P<0.05),見表2。
Tab.2 Comparison of mRNA expression levels of MMP-9,c-Src,CK and Integrin β3 between the four groups of osteoclasts表2 各組破骨細胞MMP-9、c-Src、CK、Integrin β3 mRNA表達水平比較 (n=3,±s)
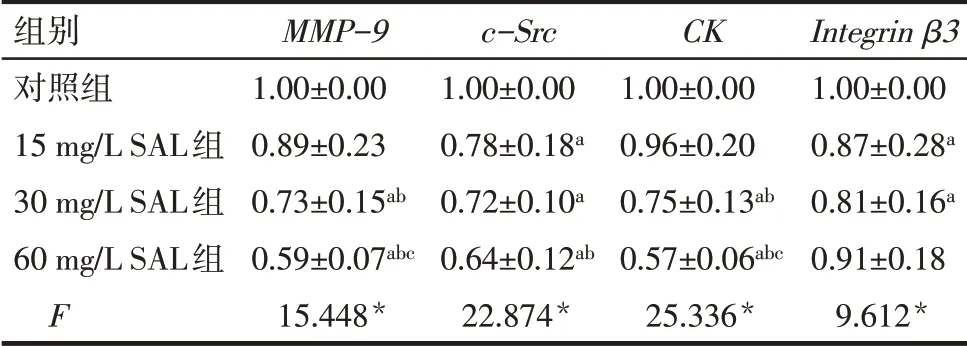
Tab.2 Comparison of mRNA expression levels of MMP-9,c-Src,CK and Integrin β3 between the four groups of osteoclasts表2 各組破骨細胞MMP-9、c-Src、CK、Integrin β3 mRNA表達水平比較 (n=3,±s)
*P<0.05;a 與對照組比較,b 與15 mg∕L SAL 組比較,c 與30 mg∕L SAL組比較,P<0.05。
組別對照組15 mg∕L SAL組30 mg∕L SAL組60 mg∕L SAL組F MMP-9 1.00±0.00 0.89±0.23 0.73±0.15ab 0.59±0.07abc 15.448*c-Src 1.00±0.00 0.78±0.18a 0.72±0.10a 0.64±0.12ab 22.874*CK 1.00±0.00 0.96±0.20 0.75±0.13ab 0.57±0.06abc 25.336*Integrin β3 1.00±0.00 0.87±0.28a 0.81±0.16a 0.91±0.18 9.612*
2.5 SAL 抑制破骨細胞MMP-9、c-Src 蛋白表達水平 與 對 照 組 比 較,15、30 和60 mg∕L SAL 組 的MMP-9、c-Src蛋白表達水平逐漸降低(P<0.05),見圖4、表3。
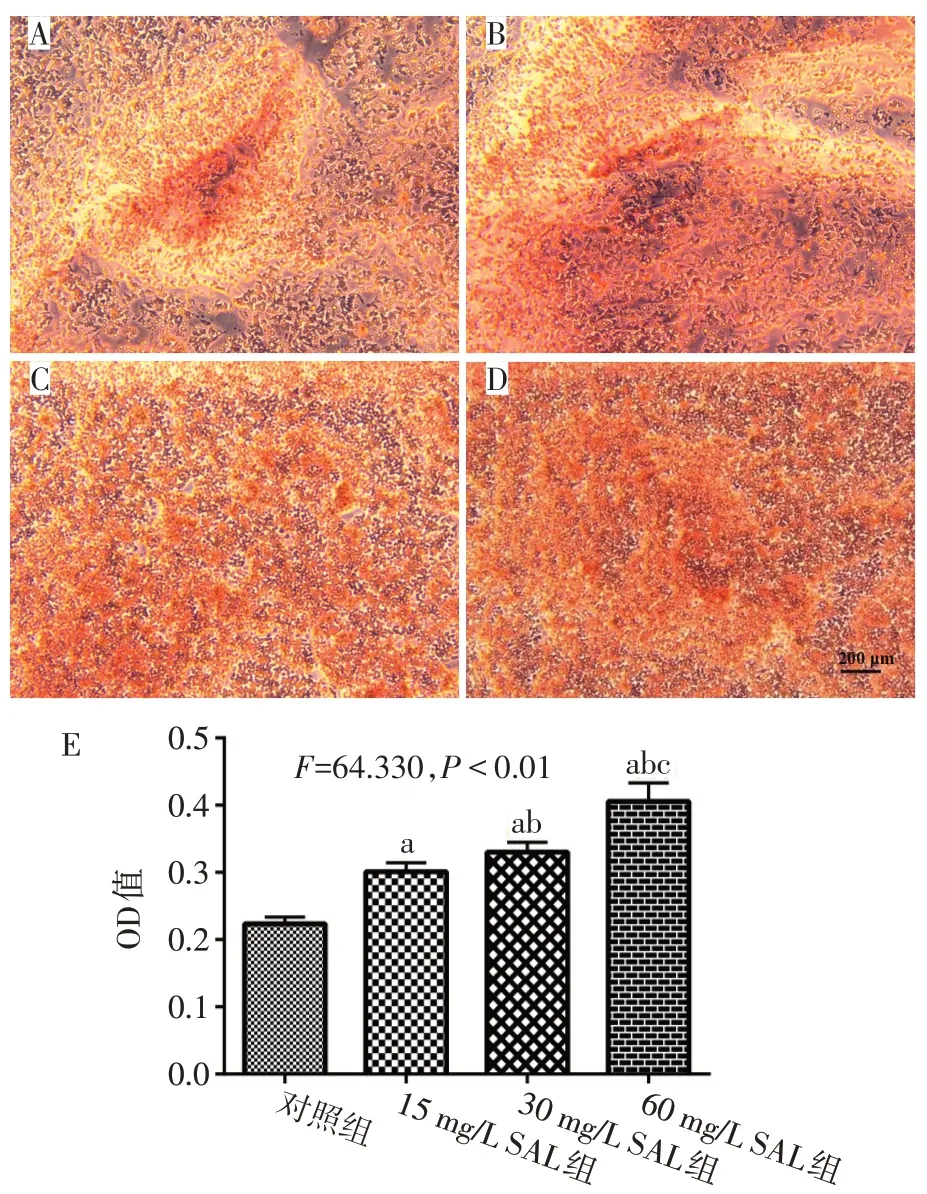
Fig.3 Effects of SAL on alizarin red staining of osteoclast calcium nodules圖3 SAL對破骨細胞鈣結節茜素紅染色的影響
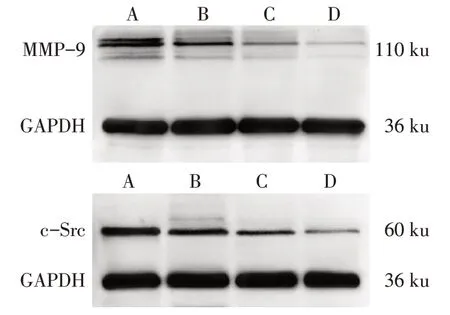
Fig.4 Effects of SAL on MMP-9 and c-Src protein expression圖4 SAL對破骨細胞MMP-9、c-Src蛋白表達的影響
3 討論
骨是一種具有礦化能力的結締組織,在骨重塑和骨微環境穩態中,破骨細胞和成骨細胞之間存在信號傳導現象[11]。破骨細胞可以通過空泡腺苷三磷酸(ATP)酶、補體C3a、微小RNA 等影響成骨細胞骨形成;同時,成骨細胞也可以通過骨保護素∕核因子受體活化因子∕核因子受體活化因子配體(OPG∕RANK∕RANKL)、死亡受體∕死亡配體(Fas∕FasL)等通路影響破骨細胞的分化與凋亡[12]。破骨細胞完成骨吸收主要是依靠其特有的超微結構,通過細胞骨架重組形成極化構象,將細胞表面膜分成封閉區、褶皺緣、基底外側區和功能分泌區[13]。極化后的破骨細胞通過足質體貼附于骨表面,同時F-Actin環圍繞褶皺緣形成獨立的微環境,CK 和MMP-9 等水解酶的釋放可以實現骨吸收[14]。
Tab.3 Comparison of relative expression levels of MMP-9 and c-Src protein between the four groups表3 各組MMP-9、c-Src蛋白相對表達量的比較(n=3,±s)
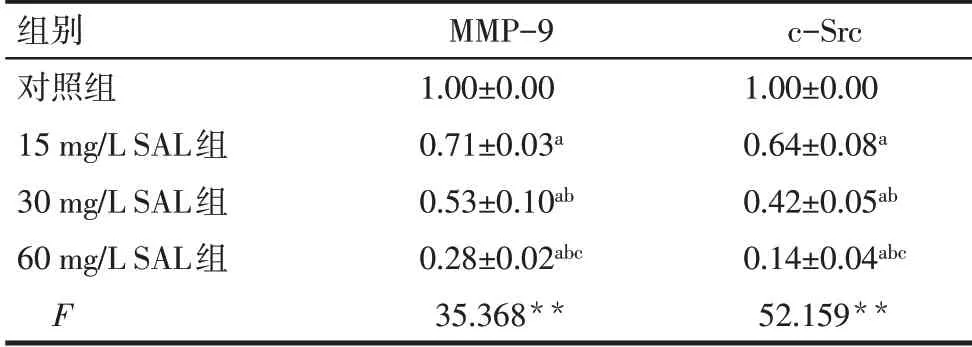
Tab.3 Comparison of relative expression levels of MMP-9 and c-Src protein between the four groups表3 各組MMP-9、c-Src蛋白相對表達量的比較(n=3,±s)
**P<0.01;a與對照組比較,b與15 mg∕L SAL 組比較,c與30 mg∕L SAL組比較,P<0.05。
組別對照組15 mg∕L SAL組30 mg∕L SAL組60 mg∕L SAL組F MMP-9 1.00±0.00 0.71±0.03a 0.53±0.10ab 0.28±0.02abc 35.368**c-Src 1.00±0.00 0.64±0.08a 0.42±0.05ab 0.14±0.04abc 52.159**
中藥紅景天具有補腎、扶正固本、理氣養血和滋補強身等功效。現代研究結果表明,紅景天含有40多種化合物,主要成分為SAL、酪醇等,具有抗缺氧、抗疲勞、抗腫瘤、抗病毒等作用[15-16]。研究還發現,其主要活性成分SAL具有上調成骨細胞骨形態發生蛋白-2的作用,能促進成骨性的骨生成[17]。本實驗發現,SAL 能夠抑制sRANKL 誘導的破骨細胞分化和極化、F-Actin環形成,促進鈣結節形成,抑制破骨細胞骨吸收關鍵蛋白酶MMP-9、c-Src 的活性,顯示出確切的抗骨質疏松作用。
基于SAL 確切的抑制骨吸收作用,本研究觀察了其對破骨細胞分化、極化和骨吸收的影響。破骨細胞由sRANKL誘導巨噬細胞分化而成。在破骨細胞分化和極化過程中,破骨細胞特異性表達骨吸收標志蛋白MMP-9、c-Src 等[18]。MMP-9 為破骨細胞分泌的蛋白水解酶,參與細胞外膠原基質的降解,維持破骨細胞的骨吸收功能。MMP-9 在哺乳動物胚胎期軟骨內成骨及骨重建時起到膠原酶的作用,是破骨細胞和血管內皮細胞侵入礦化組織所必須的生物因子,破骨細胞分化過程中MMP-9 表達上調,加速骨吸收[19]。因此,SAL可能通過抑制MMP-9的表達來增加鈣結節茜素紅染色程度。c-Src 及其磷酸化產物p-Src 參與調節破骨細胞極化及褶皺緣的形成。F-Actin 環是破骨細胞特有的進行骨吸收的細胞骨架蛋白,反映了破骨細胞的骨吸收功能[20]。本實驗結果表明,SAL 能顯著減少由sRANKL 誘導的破骨細胞數目,抑制破骨細胞的分化與極化,減少F-Actin 環形成,促進鈣結節增多,降低破骨細胞的MMP-9 及c-Src 的表達,表明SAL 能夠抑制破骨細胞骨吸收,減少骨丟失。
綜上,本研究發現,SAL能夠抑制破骨細胞的分化、極化和骨吸收作用,為SAL的開發和臨床應用提供了依據。本研究的不足之處是僅觀察了基因與蛋白水平的變化,尚未進行相關信號通路的深入研究。另外,本研究僅探究SAL 抑制體外破骨細胞的分化和極化,未通過體內實驗證實,有待于后續研究進一步探討。
