皮質前額葉和海馬PPAR-α通路參與N-棕櫚酰乙醇胺抗大鼠抑郁樣行為
2022-04-02 07:57:08李瑞瑞張露文張渺于海玲
天津醫藥 2022年3期
李瑞瑞,張露文,張渺,于海玲
抑郁癥是一種以顯著而持久的情緒低落為臨床特征的精神障礙,預計到2030年將成為全球第一大疾病負擔[1]。目前,以選擇性5-羥色胺再攝取抑制劑為代表的新型抗抑郁藥為臨床主要用藥,但存在起效慢、臨床治愈率低、不良反應較多等缺點[2]。內源性大麻素系統由內源性大麻素、內源性大麻素受體及與其合成相關的代謝酶類組成,參與調節情緒、記憶、認知、植物神經系統和自主活動等神經活動進程[3-4]。研究表明,內源性大麻素系統的功能障礙會促進焦慮和抑郁等精神疾病的發生,皮質前額葉(prefrontal cortex,PFC)和海馬分泌的內源性大麻素可防止應激誘導的行為改變,產生抗焦慮和抗抑郁作用[5-7]。N-棕櫚酰乙醇胺(N-palmitoylethanolamide,PEA)是內源性大麻素類似物,為過氧化物酶體增殖物 激 活α受 體(peroxisome proliferator-activated receptor α,PPARα)的內源性激動劑,具有抗炎、鎮痛、神經保護等活性,在抑郁動物模型中已表現出抗抑郁和抗焦慮樣作用[8-9]。有研究證明,PEA可改善小鼠的疼痛和認知障礙,而在PPARα基因敲除小鼠中上述作用大多消失[10]。本實驗室前期研究發現,PEA可改善海馬組織內部分氧化應激因子的異常表達,上調PFC 的突觸素表達,使海馬和PFC 內PPARα 表達恢復正常,從而減輕大鼠抑郁樣行為[9,11]。本研究旨在進一步探究PEA 通過PFC 和海馬的PPARα途徑發揮抗抑郁作用的可能機制,為抑郁癥的治療提供潛在靶點。
1 材料與方法
1.1 材料
1.1.1 實驗動物 SPF 級健康雄性SD 大鼠50 只,體質量180~200 g,購自延邊大學實驗動物中心,許可證號:SCXK(吉)2017-0003。大鼠自由攝食飲水,在自然光照、室溫(23±2)℃、相對濕度(55±5)%的清潔空調室飼養,嚴格遵循國家《實驗動物管理條例》。
1.1.2 實驗試劑 PEA(延邊大學藥學院提供,化學結構式見圖1);鹽酸氟西汀分散片(百憂解,20 mg∕片,禮來蘇州制藥);PPARα拮抗劑MK886(上海瀚香生物);鼠源多唾液酸-神經細胞黏附分子(polysialic acid neural cell adhesion molecule,PSA-NCAM)抗體、辣根過氧化物酶標記的山羊抗小鼠IgG二抗(SIGMA);DAB顯色試劑盒(北京中杉金橋);腦源性神經營養因子(brain-derived neurotrophic factor,BDNF)、膠質細胞源性神經營養因子(glial cell line-derived neurotrophic factor,GDNF)、腫瘤壞死因子-α(tumor necrosis factor-alpha,TNF-α)、白細胞介素-1β(interleukin-1β,IL-1β)和核轉錄因子(nuclear factor,NF)-κB酶聯免疫吸附試驗(ELISA)檢測試劑盒均購于南京森貝伽生物科技公司。
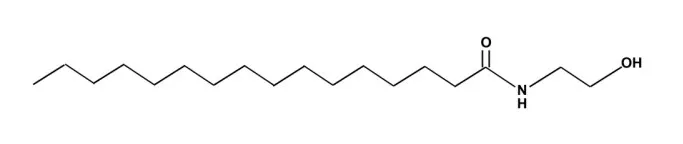
Fig.1 Structure of N-palmitoylethanolamide圖1 N-棕櫚酰乙醇胺結構式
1.1.3 實驗儀器 DW-40L262 醫用低溫保存箱(青島海爾特種電器有限公司),PRO200 勻漿器(美國PROScientific),TGL-16aR 高速冷凍離心機(上海安亭科學儀器廠),DW-2102A酶聯免疫檢測儀(北京六一生物科技有限公司),BX53光學倒置顯微鏡(日本奧林巴斯)。
1.2 動物分組、模型制備及實驗流程 大鼠適應性飼養1周后,按照隨機數字表法分為正常對照組、模型組、氟西汀組、PEA 組和PEA+MK886 組,每組10 只。除正常對照組外,其余4 組每天上午8:00 均給予慢性不可預見性溫和應激(CUMS)和孤養應激,連續4 周。CUMS 干預1 周后開始灌胃給藥,劑量均為10 mg∕kg,PEA+MK886 組在PEA 灌胃后腹腔注射MK886(3 mg∕kg)。
除正常對照組外,其余每組每日接受以下2 種不同的CUMS:電擊足底20 次、5 ℃水中游泳5 min、45 ℃水中游泳5 min、禁食禁水18 h、50 ℃熱刺激10 min、潮濕墊料17 h、合籠飼養17 h、空籠17 h、45°傾斜鼠籠24 h、夾尾1 min、懸尾1 min、異物放置24 h、燈光閃爍和晝夜循環顛倒24 h 等。動物每相鄰2 d 接受刺激不重復,確保大鼠不能預測特定刺激的出現。
1.3 腦切片的制備及免疫組化染色檢測PSA-NCAM 表達水平 實驗第36 天,每組按照隨機數字表法取3 只大鼠,將大鼠麻醉后處死,用預冷的生理鹽水從左心室灌注到右心房,待流出液變清透后,繼續灌注4%多聚甲醛(0.1 mol∕L 的PBS配制,pH=7.4)進行組織固定。低溫下迅速取出全腦,于4%多聚甲醛溶液中固定12 h后進行石蠟包埋與切片。
石蠟切片于烘箱60 ℃烤片,60 min 后進行脫蠟和水化,二甲苯溶液沖洗2 次,15 min∕次,無水乙醇2 次,10 min∕次,95%和85%乙醇各1 次,5 min∕次,結束后PBS 洗滌3 次,5 min∕次,抗原熱修復。冷卻至室溫后PBS洗滌3次,5 min∕次,滴加適量內源性過氧化物酶阻斷劑覆蓋于組織表面,室溫放置15 min,PBS 洗滌3 次,5 min∕次。切片滴加PSA-NCAM 鼠源抗體(1∶150),4 ℃過夜。次日室溫放置30 min后滴加適量的反應增強劑,PBS 洗滌3 次,5 min∕次。滴加適量辣根過氧化物酶標羊抗小鼠IgG 抗體,室溫30 min,PBS 洗滌3 次,10 min∕次。滴加DAB顯色液,適時流水沖洗,蘇木素復染,自來水沖洗返藍后進行脫水,最后用中性樹膠進行封片。光學倒置顯微鏡下觀察并拍照,Image Pro Plus 6.0 軟件分析平均光密度值。
1.4 腦組織取樣本采集和ELISA 檢測 剩余大鼠于樣本采集前12 h 禁食不禁水,麻醉大鼠后快速取腦,在低溫操作臺上分別分離PFC 和海馬組織,液氮速凍后置于-80 ℃冰箱保存待用。參照ELISA試劑盒說明書的步驟進行操作,根據標準品所得的標準曲線計算出大鼠PFC 中BDNF、GDNF 含量,及PFC 和海馬中TNF-α、IL-1β 和NF-κB 含量。BDNF、GDNF和IL-1β含量分別用ng∕g腦組織表示,NF-κB和TNFα含量用μg∕g腦組織表示。
1.5 統計學方法 采用SPSS 25.0 進行數據分析,采用GrapPad Prism 7.0 做圖。符合正態分布的計量數據采用均數±標準差(±s)表示,正常對照組、模型組、氟西汀組、PEA組間各指標比較采用單因素方差分析,組間多重比較采用LSD-t法;PEA+MK886組和PEA組間各指標比較采用獨立樣本t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 各組大鼠PFC 和海馬中PSA-NCAM 蛋白表達水平比較 正常對照組的PFC、海馬DG區以及CA3區的PSA-NCAM 陽性免疫反應產物呈棕黃色顆粒,數量較多,排列密集,染色深,神經元胞核被染成藍色。與正常對照組相比,模型組的PSA-NCAM 陽性免疫反應產物排列稀疏、密度明顯降低,染色變淺,平均光密度顯著降低(P<0.05)。與模型組相比,氟西汀組和PEA 組大鼠PFC 和海馬CA3 區PSANCAM免疫反應產物數量增多,顏色較深,平均光密度顯著增高(P<0.05)。與PEA 組相比,PEA+MK886 組的PSA-NCAM 陽性免疫反應產物染色變淺,平均光密度降低(P<0.05),見圖2~4。
2.2 各組大鼠PFC中BDNF 和GDNF 含量比較 與正常對照組相比,模型組大鼠PFC 中的BDNF、GDNF含量均顯著降低(P<0.05);與模型組相比,氟西汀組和PEA 組PFC 中的BDNF、GDNF 含量升高(P<0.05)。與PEA 組相比,PEA+MK886 組PFC 中BDNF、GDNF的含量均顯著降低(P<0.05),見表1。
2.3 各組大鼠PFC 和海馬中TNF-α、IL-1β 和NFκB含量比較 與正常對照組相比,模型組大鼠PFC和海馬中TNF-α、IL-1β 和NF-κB 含量均顯著增加(P<0.05);與模型組相比,氟西汀組和PEA 組大鼠PFC 中TNF-α、IL-1β 和NF-κB 含量均降低(P<0.05);與PEA 組相比,PEA+MK886組大鼠PFC 和海馬中TNF-α、IL-1β 和NF-κB 的含量均顯著增加(P<0.05),見表2。
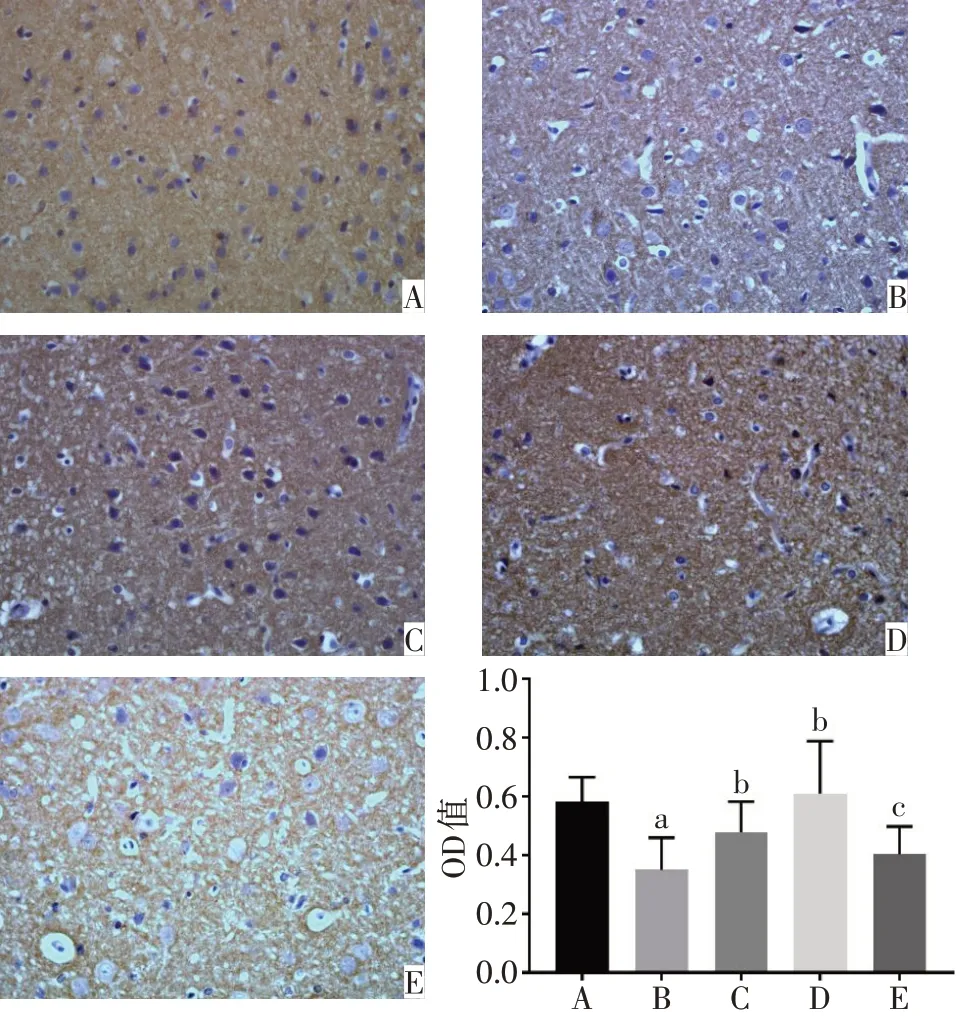
Fig.2 Immunohistochemical staining results of the expression of PSANCAM in prefrontal cortex of rats(×400)圖2 各組大鼠PFC中PSA-NCAM蛋白表達的免疫組化染色結果(×400)
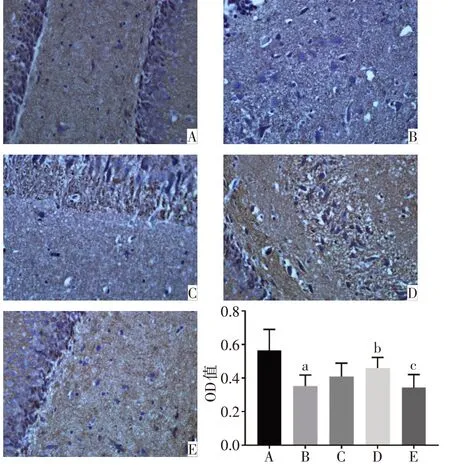
Fig.3 Immunohistochemical staining results of the expression of PSA-NCAM in DG of hippocampus of rats(×400)圖3 各組大鼠海馬DG區PSA-NCAM蛋白表達的免疫組化染色結果(×400)
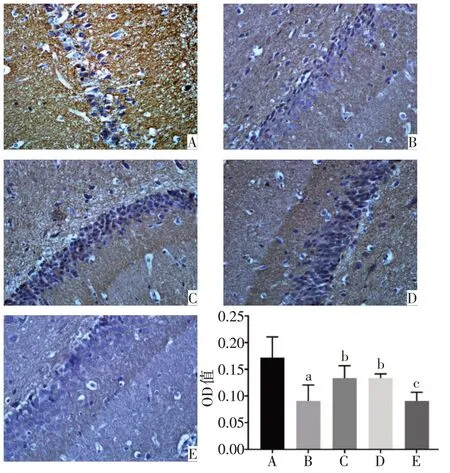
Fig.4 Immunohistochemical staining results of the expression of PSA-NCAM in CA3 of hippocampus of rats(×400)圖4 各組大鼠海馬CA3區PSA-NCAM蛋白表達的免疫組化結果(×400)
Tab.1 Comparison of the content of BDNF and GDNF in PFC tissue between the five groups表1 各組大鼠PFC中BDNF和GDNF含量的比較(n=7,ng∕g,±s)
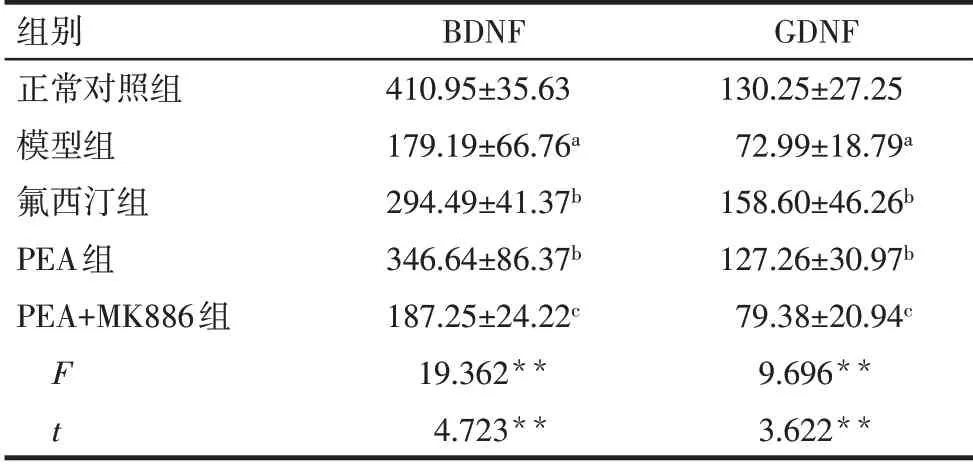
Tab.1 Comparison of the content of BDNF and GDNF in PFC tissue between the five groups表1 各組大鼠PFC中BDNF和GDNF含量的比較(n=7,ng∕g,±s)
**P<0.01;a與正常對照組比較,b與模型組比較,c與PEA組比較,P<0.05。
組別正常對照組模型組氟西汀組PEA組PEA+MK886組F t BDNF 410.95±35.63 179.19±66.76a 294.49±41.37b 346.64±86.37b 187.25±24.22c 19.362**4.723**GDNF 130.25±27.25 72.99±18.79a 158.60±46.26b 127.26±30.97b 79.38±20.94c 9.696**3.622**
3 討論
近年來,應激被認為是誘導抑郁癥等精神類疾病的主要因素之一。應激是指外界環境劇烈變化時,機體的生理和心理失衡而出現的精神、神經內分泌和免疫等方面的反應[12]。經CUMS刺激建立的抑郁動物模型,與人類抑郁癥的發病機制接近,已被廣泛應用于抑郁癥機制研究和藥物篩選,此模型主要以測量“快感缺失”來判斷模型是否成功[13]。本實驗室前期研究已證實,CUMS 誘導大鼠出現了行為異常和生理功能障礙,抑郁癥模型建立成功,經過28 d的PEA 治療后,大鼠的體質量增長,自發活動、蔗糖偏好率均明顯增加,表明大鼠的食欲和情緒有所改善、消化系統功能有所好轉,探索行為和自發活動性及好奇程度都有所改善,獎賞反應和快感提升[9,11]。本研究進一步探究PEA改善大鼠抑郁樣作用的機制。
PFC和海馬是與認知、情緒密切相關的腦區,兩者具有內在聯系,PFC 在聯想恐懼和獎勵學習的神經元回路中起著關鍵的調節作用,在慢性應激后PFC和海馬可能出現結構可塑性的改變,導致焦慮、抑郁情緒,認知改變等[14]。突觸是一種高特異性的細胞黏附結構,黏附分子和糖蛋白類在其形成、成熟和功能調節中起到重要作用。PSA-NCAM 是一種帶有唾液酸的特殊神經細胞黏附分子,在神經細胞黏附、軸突生長以及對神經元前體細胞的遷移等方面有重要作用。PSA-NCAM 與神經元的生長、發育和存活有密切聯系,同時PSA-NCAM 也參與了突觸的生長,突觸可塑性形成和維持過程[15]。本研究中,PEA 處理后PFC 以及海馬DG 和CA3 區的PSANCAM 蛋白表達較模型組均明顯增加。BDNF 和GDNF參與神經元的存活、分化和生長,在神經系統的生長過程中發揮營養和保護作用[16]。研究發現,抑郁癥患者BDNF 及GDNF 水平的下降可導致腦神經元損傷后修復、再生能力下降,進而影響5-羥色胺、多巴胺等神經遞質的合成及分泌,并影響抗抑郁藥的治療效果[17-18]。本課題組前期研究發現,PEA可以逆轉CUMS 誘導的海馬BDNF 和GDNF 水平的下調[11]。本次實驗中,PEA組PFC中BDNF和GDNF水平較模型組升高,提示PEA可促進神經元增殖、遷移、軸突生長和突觸形成,增加PFC和海馬的神經可塑性。
Tab.2 Comparison of TNF-α,IL-1β and NF-κB contents in PFC and hippocampus between the five groups表2 各組大鼠PFC和海馬中TNF-α、IL-1β和NF-κB含量比較 (n=7,±s)
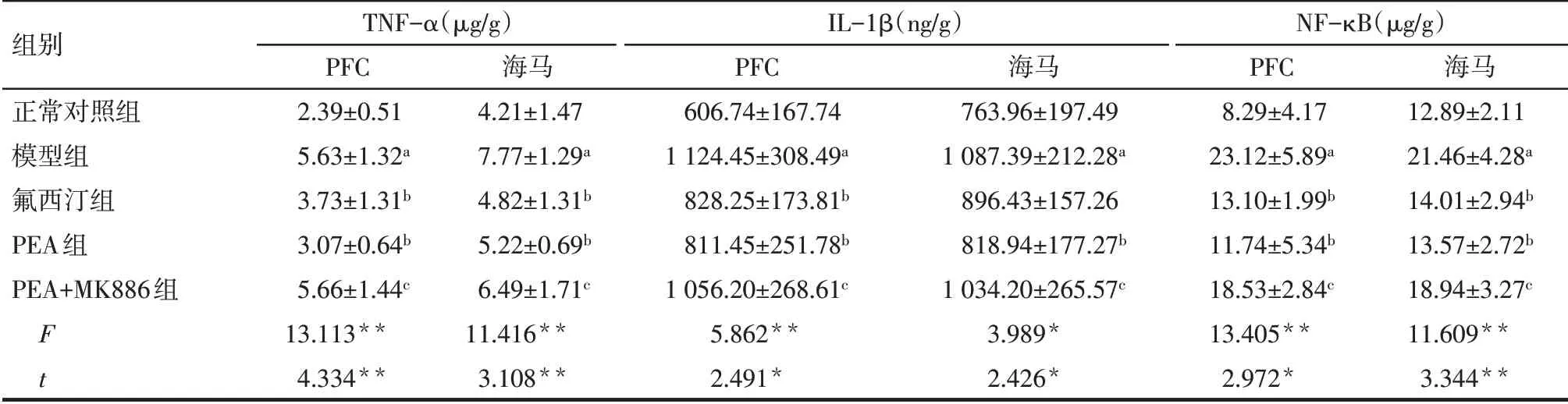
Tab.2 Comparison of TNF-α,IL-1β and NF-κB contents in PFC and hippocampus between the five groups表2 各組大鼠PFC和海馬中TNF-α、IL-1β和NF-κB含量比較 (n=7,±s)
*P<0.05,**P<0.01;a與正常對照組比較,b與模型組比較,c與PEA組比較,P<0.05。
組別正常對照組模型組氟西汀組PEA組PEA+MK886組F t TNF-α(μg∕g)PFC 2.39±0.51 5.63±1.32a 3.73±1.31b 3.07±0.64b 5.66±1.44c 13.113**4.334**海馬4.21±1.47 7.77±1.29a 4.82±1.31b 5.22±0.69b 6.49±1.71c 11.416**3.108**IL-1β(ng∕g)PFC 606.74±167.74 1 124.45±308.49a 828.25±173.81b 811.45±251.78b 1 056.20±268.61c 5.862**2.491*海馬763.96±197.49 1 087.39±212.28a 896.43±157.26 818.94±177.27b 1 034.20±265.57c 3.989*2.426*NF-κB(μg∕g)PFC 8.29±4.17 23.12±5.89a 13.10±1.99b 11.74±5.34b 18.53±2.84c 13.405**2.972*海馬12.89±2.11 21.46±4.28a 14.01±2.94b 13.57±2.72b 18.94±3.27c 11.609**3.344**
研究發現,精神疾病患者的腦組織中有多種炎癥標志物高表達,腦內炎性細胞因子水平的增加可引起神經細胞損傷,這為抑郁癥的細胞因子學說提供了有力支持[19]。TNF-α 通過促使炎癥介質的迅速生成導致細胞因子的級聯放大效應,從而導致“瀑布效應”,是判斷炎癥反應的可靠指標[20]。IL-1β是炎癥反應調控中關鍵的炎性細胞因子,也是體內誘導炎癥作用較強的炎癥介質之一。NF-κB 參與免疫、炎癥、應激反應以及細胞增殖和凋亡等過程,它作為炎癥信號通路中的關鍵調節因子,常被視為調控炎癥反應的重要靶點[21-22]。本研究發現,CUMS誘導后大鼠PFC 和海馬中的TNF-α、IL-1β 和NF-κB含量出現升高,經PEA 干預后明顯下降,提示PEA可能通過抑制NF-κB 信號通路從而減少中樞神經系統TNF-α、IL-1β等的釋放,阻斷炎癥的級聯放大效應,從而減輕過度炎癥引發的PFC 和海馬中神經元損傷,緩解慢性應激大鼠的焦慮抑郁樣行為表型。
在采用選擇性PPARα拮抗劑MK886預處理后,PEA對大鼠生理功能和行為學的改善作用不同程度被消除,本實驗室前期研究結果顯示PEA 可以上調CUMS大鼠PFC和海馬中PPARα的蛋白和mRNA的表達[9,11],進一步證實PEA改善CUMS大鼠的抑郁樣行為的機制可能與PPARα信號通路有關。
綜上所述,PEA 可通過調控PFC 和海馬的PPARα 通路,促進PFC 和海馬的神經元增殖、突觸形成,增加神經可塑性;同時可通過減少炎性細胞因子的分泌來減輕PFC 和海馬中神經元損傷,改善慢性應激大鼠的焦慮抑郁樣行為表型。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
