賀蘭山低山區土壤種子庫空間分布特征
2022-04-06 06:53:04李國旗王雅芳謝博勛
西北植物學報 2022年2期
關鍵詞:物種
李國旗,王雅芳,劉 星,謝博勛,解 盛,石 云
(1 寧夏大學西北土地退化與生態恢復國家重點實驗室培育基地,銀川 750021;2 寧夏大學西北退化生態系統恢復與重建教育部重點實驗室,銀川 750021)
賀蘭山地處中國西北干旱區,具有重要的地理意義,其作為最后一道生態屏障,阻隔了毛烏素沙地、烏蘭布和沙漠、騰格里沙漠這三大沙漠進一步向銀川平原擴張,攔截了源自蒙古國、中國阿拉善地區等的沙塵暴;賀蘭山的生態安全不僅影響其周邊的黃河段、銀川平原,還可能進一步影響西北地區、華北地區甚至全國的生態安全[1-2]。賀蘭山地處華北平原、蒙古高原、青藏高原的交匯處,其地理位置決定了該地區生物類型復雜多樣,且具有交叉性和過渡性[3-4];賀蘭山位于溫帶草原與荒漠的交匯處,山體高聳,生物類型具有明顯的空間分異規律[5-6]。植物優勢種主要有短花針茅(Stipabreviflora)、蒙古扁桃(Amygdalusmongolica)、沙蒿(Artemisiadesertorum)、青海云杉(Piceacrassifolia)等[7-8]。
土壤種子庫是存在于土壤表面和土壤中有生命活力的種子的總量[9-10]。也可以定義為單位面積土壤中的全部有生命活力的種子[11-13]。土壤種子庫可作為植物的潛在種群,影響植被自然更新和恢復、生物多樣性保護、植被的演替進程和方向[14-16]。大量研究表明,環境空間異質性是制約或影響植物種群、群落乃至生態系統和物種多樣性等的重要因素,其中地形是主要來源[17-18]。地形通過其單一的生態作用或受一定的地形復合體影響地表的光、熱、水、土、肥等生態因子,從而改變能量的平衡,直接或間接地影響植被格局。而土壤種子庫中的種子主要來源于地上植被,地形可通過影響地上植被進一步影響土壤種子庫[19-20]。
對賀蘭山在不同海拔高度上開展土壤種子庫調查研究,主要目的是:① 探明土壤種子庫特征的空間差異性;② 比較研究各海拔土壤種子庫與相應地上植被的物種組成關系的差異性;③ 評估土壤種子庫對植被自然恢復與更新的貢獻潛力在海拔間的差異性。賀蘭山低山區風蝕嚴重、植被稀疏,植被自然恢復潛力尚未可知,對不同海拔高度種子庫開展研究,探討地上植被自然恢復潛力在海拔間的差異性,以期為賀蘭山低山區植被合理利用、恢復治理與生態重建提供理論依據。
1 研究區概況與研究方法
1.1 研究區概況
研究區位于寧夏回族自治區賀蘭山國家自然保護區,地理位置 105°49′~106°41′E,38°19′~39°22′N,東接銀川平原,西側和北側與阿拉善戈壁荒漠相接,位于溫帶草原與荒漠的交匯處[1-2]。賀蘭山地處中國西北中溫帶氣候區,為典型溫帶大陸性氣候并具有山地氣候特征。無霜期229 d,春季風大、沙多,氣候變化大,年均溫-0.8 ℃。年均降水量420 mm,年均蒸發量2 000 mm,降雨集中在7~9月份,占全年降水量的70%~80%[20]。低山區氣溫變化劇烈,干旱少雨,為顯著大陸性氣候。山地植被和土壤具有明顯的空間分異規律,海拔由低到高,植被可劃分為山前荒漠與荒漠草原帶(海拔1 600 m以下)、山麓與低山草原帶(海拔1 600~1 800 m)、中山和亞高山針葉林帶(海拔1 800~3 100 m)和高山亞高山灌叢、草甸帶(海拔3 100 m以上)4個垂直帶,土壤劃分為棕鈣土帶——灰褐土帶——高山亞高山灌叢、草甸土帶這3個帶[3-4]。
1.2 樣地設計與樣品采集
從海拔1 200到2 000 m,每隔200 m設置1個取樣點,選取群落生境相對均勻的地方,設置3個5 m×5 m的大樣方,進行灌木調查;在每個大樣方內沿對角線布置3個1 m×1 m的小樣方進行草本調查。調查記錄每個樣方內各植物物種名稱、蓋度、高度、多度等,其中蓋度采用方格網法直接計算;高度用測高尺測量,做3~5個重復;多度完全記錄小樣方內每種植物。在植被調查的同時在每個小樣方中采用五點取樣法采集土樣(取樣器采用20 cm×20 cm×10 cm規格)。
1.3 實驗方法
采用常規分析法測定土壤理化性質(表1)。土壤種子庫采用室內萌發鑒定的方法。將土樣均勻的平鋪在萌發盆里,厚度約為2 cm,盆底還鋪有5 cm的無籽毛細沙。將處理好的萌發盆置溫室中,采取少澆、勤澆的方式,保持土壤濕潤,等待種子萌發。幼苗種屬鑒定后去除,未鑒定出的繼續生長,直至可以鑒定出,連續4周沒有新幼苗萌發結束實驗。土壤種子庫中種子密度用單位面積(1 m2)土壤中有效種子數量來表示。

表1 土壤理化性質的測定方法
1.4 數據統計與分析
1.4.1 地上植被的物種重要值計算根據對5個海拔地上植被群落特征的調查研究,選取相對多度、相對蓋度、相對頻度及相對高度4個因素來計算植物各物種重要值,計算公式為:
重要值(IV)=(相對多度+相對蓋度+相對頻度+相對高度)/4
相對多度=(某一種多度/所有種多度之和)×100
相對蓋度=(某一種蓋度/所有種蓋度之和)×100
相對頻度=(某一種頻度/所有種頻度之和)×100
相對高度=(某一種的平均高度/所有種的平均高度之和)×100
1.4.2 物種多樣性指標計算Margalef 豐富度指數:
R=(S-1)/lnN;
Shannon-wiener 指數:
H′=∑PilnPi;
Simpson 優勢度指數:
Pielou 均勻度指數:
E=H′/lnS
式中:S為物種總數;Pi=Ni/N,N為樣方中總的個體數,Ni為樣方中第i物種的個體數。
1.4.3 相似性指數計算不同海拔高度土壤種子庫與地上植被的相似性、海拔間土壤種子庫的相似性以及地上植被的相似性,均采用Sorensen的相似性系數(similarity coefficient,SC)來表示,計算公式為:
式中SC為相似性系數;w代表土壤種子庫和地上植被共同擁有的物種數;a代表土壤種子庫中物種數;b代表地上植被物種數。
SC值在0~0.24為極不相似;0.25~0.49為中等不相似;0.50~0.74為中等相似;0.75~1.00為極相似。
1.5 數據統計與分析
野外調查及室內試驗所得實驗數據在Excel表格中進行初步處理,在SPSS 21.0軟件中進行多重比較和相關性分析,在Canoco4.5進行RDA分析。不同海拔間土壤各性質、種子庫特征及地上植被特征均采用單因素方差分析和LSD法進行多重比較,用Person相關系數法進行相關性分析,土壤種子庫物種多樣性與土壤因子間的關系進行RDA分析與制圖。
2 結果與分析
2.1 不同海拔高度土壤種子庫特征
2.1.1 土壤種子庫密度由表2可知,不同海拔高度土壤種子庫密度具有差異性。5個海拔高度土壤種子庫總密度分別為(825±16.07)、(390±10.41)、(685±5.00)、(30±5.00)和(860±18.93)seeds·m-2,表現為2 000 m>1 200 m>1 600 m>1 400 m>1 800 m,且差異顯著(P<0.05)。其中,一年生草本種子密度表現為1 200 m>1 600 m>1 400 m>2 000 m>1 800 m,多年生草本種子密度表現為1 800 m>1 400 m>2 000 m=1 200 m=1 600 m,灌木或小灌木種子庫密度表現為2 000 m>1 800 m=1 200 m,且海拔間均差異顯著(P<0.05)。

表2 不同海拔土壤種子庫組成及密度
海拔2 000和1 200 m處土壤種子庫總密度分別為(860±18.93)和(825±16.07)seed·m-2,顯著高于其他海拔,但兩者種子來源并不相同。海拔1 200 m處種子主要來源于一年生草本,為(810±15.28)seed·m-2,顯著高于其他海拔;海拔2 000 m土壤種子庫主要來源于灌木和小灌木,為(710±7.64)seed·m-2,顯著高于其他海拔。除此之外,海拔1 600 m土壤種子庫主要來源于多年生草本,為(120±7.64)seed·m-2,顯著高于其他海拔。
2.1.2 土壤種子庫多樣性和相似性特征如圖1所示,不同海拔高度的土壤種子庫多樣性具有差異性。物種多樣性常用Shannon-Wiener指數、Simpson優勢度指數、Pielou均勻度指數和Margalef豐富度指數來表示。5個海拔高度土壤種子庫中,全部植物種Shannon-Wiener指數為1 200 m>2 000 m>1 600 m>1 800 m>1 400 m,Simpson優勢度指數為1 600 m>1 200 m>1 800 m>2 000 m>1 400 m,Pielou均勻度指數為1 800 m>1 600 m>2 000 m>1 200 m>1 400 m,Margalef豐富度指數為1 200 m>2 000 m>1 600 m>1 800 m>1 400 m,且各指數在海拔間的變化差異均顯著(P<0.05)。
隨著海拔升高,一年生草本、灌木或半灌木的Shannon-Wiener指數和Simpson優勢度指數增大,而一年生草本的Pielou均勻度指數增大,灌木或半灌木的Margalef豐富度指數和Pielou均勻度指數均減小,且海拔間有顯著差異(P<0.05)。
如表3所示,海拔間植物群落土壤種子庫相似性不同。海拔1 200 m土壤種子庫與1 600 m土壤種子庫相似性最高,相似性系數為0.45,共同物種有5種;海拔2 000 m植物群落與1 200m、1 600 m和1 800 m海拔植物群落土壤種子庫相似性均較大,相似性系數分別為0.32、0.35、0.29,共同物種分別有4種、3種、2種;其余海拔間土壤種子庫相似性均較低。
由于明清兩朝時代接近,《四庫全書總目》(以下簡稱《四庫》)著錄明代別集的數量最多,“遠遠超過了之前各個朝代文獻數量的總和”[1],正印證了別集小序所說的“有明以后,篇章彌富”[2]12。雖然《四庫》是一部官家的目錄學代表作,卻也涉及到文學批評的范疇;不過鑒于其官學及政治背景,提要中反映出的文學評價仍有許多問題值得探討,而以晚明文學最為突出。

表3 不同海拔間土壤種子庫相似性
2.2 不同海拔土壤種子庫與相應地上植被的物種組成關系
2.2.1 土壤種子庫與地上植被物種組成由表2和表5可知,不同海拔高度土壤種子庫物種組成具有差異。在研究區土壤種子庫中共有植物22種,隸屬于7科,禾本科種類最多,為8種,藜科6種,菊科4種,蒺藜科、大戟科、豆科和玄參科各1種。其中海拔1 200 m處有植物12種,隸屬于7科,其中禾本科5種,藜科2種,蒺藜科、大戟科、豆科、玄參科和菊科各1種。
1 400 m處有植物2種,均隸屬于禾本科。1 600 m處有植物7種,隸屬于4科,其中禾本科4種,藜科、菊科和大戟科各1種。1 800 m處有植物4種,隸屬于2科,其中禾本科、菊科各2種。2 000 m處有植物10種,隸屬于3科,其中藜科5種,菊科3種,禾本科2種。隨著海拔升高,隸屬于禾本科的物種數呈下降趨勢,菊科和藜科呈增加趨勢,蒺藜科、大戟科、豆科和玄參科消失。
如表4和表5所示,樣地地上植被群落共出現33種植物,隸屬于13科,菊科種類最多,為8種,禾本科7種,藜科4種,豆科3種,唇形科和旋花科各2種,百合科、遠志科、茜草科、薔薇科、蒺藜科、鼠李科和罌粟科各1種。其中,一年生草本共有8種,隸屬于3科,藜科種類最多,為4種,菊科3種,禾本科1種;多年生草本共有17種,隸屬于8科,禾本科種類最多,為6種,菊科3種,唇形科和豆科各2種,百合科、遠志科和旋花科各1種;灌木或半灌木共有8種,隸屬于7科,菊科種類最多,為2種,旋花科、豆科、茜草科、薔薇科、蒺藜科和鼠李科各1種。

表4 不同海拔地上植被物種重要值

表5 不同海拔土壤種子庫與地上植被物種數及其科類分布
2.2.2 土壤種子庫與地上植被物種生活型構成由表6、表7可知,地上植被與土壤種子庫物種生活型構成在5個海拔高度存在差異性。其中,土壤種子庫物種生活型構成海拔間的變化差異顯著(P<0.05),而地上植被物種生活型構成在海拔間差異不顯著(P>0.05)。一年生草本在海拔1 200~1 800 m土壤種子庫占比較大,但在地上植被中占比明顯低于多年生草本和灌木或半灌木。在海拔1 200 m處土壤種子庫密度中一年生草本種子密度最大,貢獻了該海拔98.18%的種子,但地上植被占比最大的為多年生草本,這可能是由于一年生草本的繁殖策略所導致的。與其他海拔相比,海拔1 800 m土壤種子庫密度中多年生草本占比相對較高,相對應的地上植被占比最大的也為多年生草本。在海拔2 000 m處土壤種子庫密度中灌木或半灌木種子密度最大,貢獻了該海拔82.60%的種子,但地上植被占比最大的為多年生草本,這可能是由于灌木或半灌木的繁殖策略所導致的。
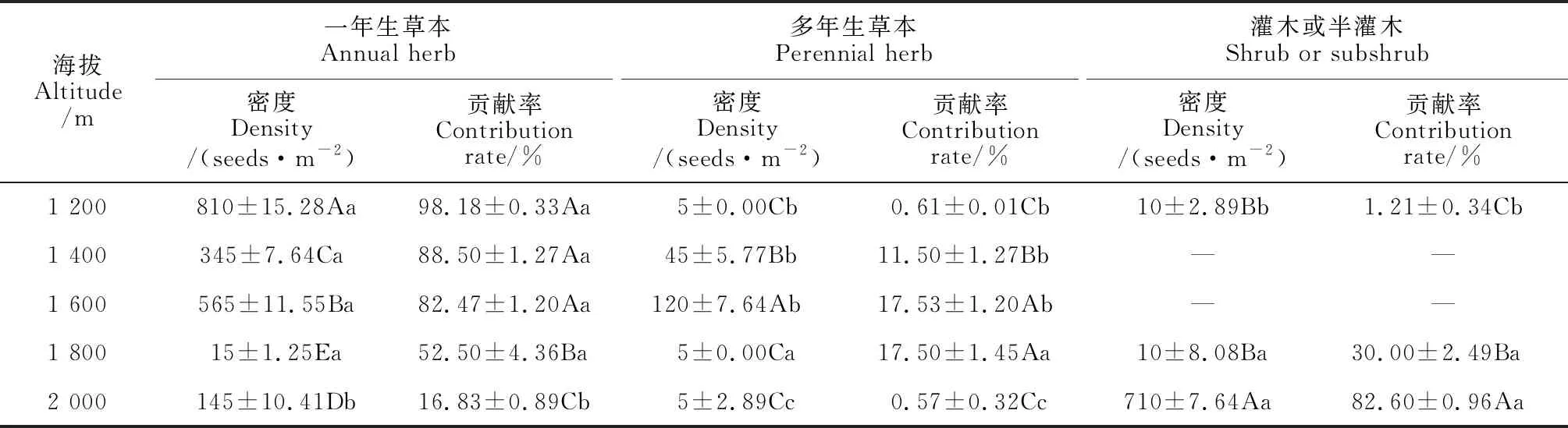
表6 不同海拔土壤種子庫物種生活型構成

表7 不同海拔地上植被物種生活型構成
2.2.3 土壤種子庫與地上植被耦合關系如表8所示,5個海拔高度地上植被與土壤種子庫的相似性均較低,且地上植被與土壤種子庫在物種組成上有顯著差異。5個海拔高度地上植被與土壤種子庫的相似性系數分別為0.12、0.33、0.11、0.12和0.12,共有種分別為3、2、1、1和1。海拔1 400 m相似性較高,為0.33;海拔1 600 m相似性較低,僅為0.11。說明該研究區不同海拔高度地上植被群落穩定性和發展趨勢具有差異性。

表8 不同海拔土壤種子庫與地上植被相似性
2.3 土壤種子庫物種多樣性與土壤理化因子的RDA分析
為了更好地揭示土壤種子庫與土壤環境因子之間的關系,采用冗余分析的方法進行排序,將不同海拔高度土壤種子庫物種多樣性指數(Shannon-Wiener指數、Simpson優勢度指數、Pielou均勻度指數和Margalef豐富度指數)作為響應變量,土壤各理化指標作為解釋變量,進行RDA分析,篩選能明顯影響土壤種子庫變化的土壤理化因子。由表9可知,RDA排序圖的前兩個排序軸特征值分別為0.708和0.248,相對應的物種數據變量積累百分比分別為70.8%和95.6%,故軸1能較好地反映土壤種子庫物種多樣性與土壤理化因子的變化特征。由圖2和表10可知,土壤種子庫物種多樣性指數與土壤容重、pH、有機質、全碳、全氮、堿解氮、速效磷呈正相關,與含水率、總孔隙度、電導率、碳氮比呈負相關,其中與pH呈顯著正相關,與電導率呈顯著負相關,即pH、電導率對土壤種子庫物種多樣性的影響較大。

表9 土壤種子庫物種多樣性與土壤理化因子的RDA排序分析
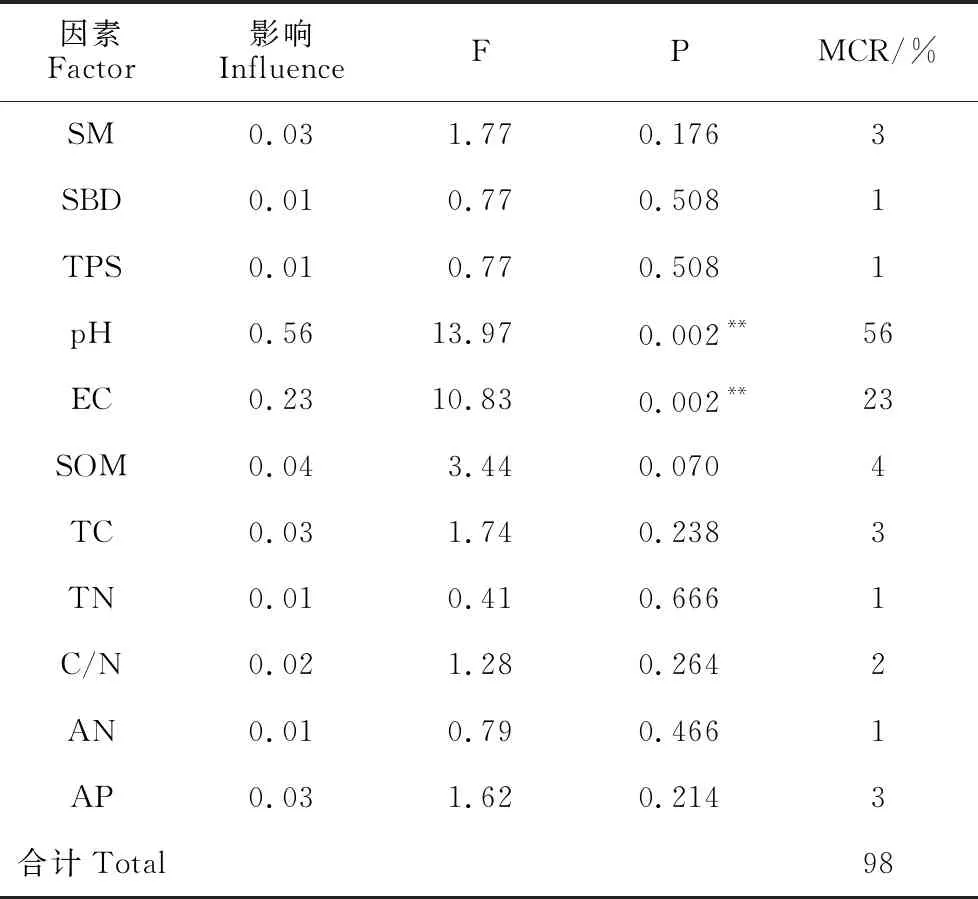
表10 土壤理化因子對土壤種子庫物種多樣性的RDA條件解釋變量
3 討論與結論
3.1 不同海拔土壤種子庫特征
土壤種子庫特征顯著影響該生態系統穩定性、演替趨勢、植被恢復潛力[21]。在本研究中,5個海拔高度的土壤種子庫種子密度、物種組成、生活型、多樣性均存在差異。海拔等地形因素顯著影響土壤種子庫種子密度[22]。海拔2 000 m和1 200 m處土壤種子庫密度顯著高于其他海拔,但其種子來源并不相同,海拔1 200 m處種子主要來源于一年生草本,海拔2 000 m土壤種子庫主要來源于灌木或半灌木,海拔1 600 m土壤種子庫主要來源于多年生草本。種子庫萌發物種以禾本科草本植物為主。隨著海拔升高,隸屬于禾本科的物種數呈下降趨勢,菊科和藜科物種呈增加趨勢,蒺藜科、大戟科、豆科和玄參科物種消失。
土壤種子庫間相似性可用來預測該植物群落發展、演替。在本研究中,5個海拔高度的植被群落土壤種子庫相似性具有差異性,海拔2 000 m植物群落與其他海拔植物群落土壤種子庫相似性均大于該海拔與其他海拔植物群落土壤種子庫相似性,可能是因為海拔2 000 m植物群落土壤種子庫中優勢種畫眉草、豬毛菜常見于其他海拔。
土壤種子庫多樣性指數常用來表征植物群落潛在種群穩定性,可用來預測群落演替方向等。物種多樣性常用Shannon-Wiener指數、Simpson優勢度指數、Pielou均勻度指數和Margalef豐富度指數來表示。在本研究中,隨著海拔升高,一年生草本、灌木或半灌木的Shannon-Wiener指數和Simpson優勢度指數顯著增加,一年生草本的Pielou均勻度顯著增加,灌木或半灌木的Margalef豐富度指數和Pielou均勻度指數顯著降低。海拔1 400 m處土壤種子庫的穩定性指數、生態優勢度指數、均勻度指數和物種豐富度指數均顯著低于其他4個海拔,即海拔1 400 m處土壤種子庫多樣性最低。海拔2 000 m處土壤種子庫表現為一年生草本和灌木或半灌木的物種多樣性有更高的穩定性、生態優勢度,一年生草本分布均勻,可能是因為海拔2 000 m處雖然物種數量多,但灌木或半灌木植物物種數量占優勢。賀蘭山低山區土壤種子庫多樣性受自身物種組成影響,還可能受地上植被制約,需進一步研究。
3.2 不同海拔土壤種子庫與相應地上植被的物種組成關系
地上植被與土壤種子庫物種組成顯著影響生態系統穩定性、演替趨勢、植被恢復潛力[21]。在本研究中,地上植被與土壤種子庫在密度、物種組成、物種多樣性以及相似性均具有較大差異,與Smith[23]、張起鵬等[24]研究結果一致,與Yuan[25]、Valk[26]不一致。本研究區內,賀蘭山5個海拔高度地上植被共計調查33種植物隸屬于13科,土壤種子庫共計萌發22種植物隸屬于7科,各海拔高度土壤種子庫物種數均顯著低于地上植被,表明賀蘭山不同海拔典型植物群落地上植被與土壤種子庫的物種組成差異較大。對5個海拔高度進行植被調查和土壤種子庫萌發實驗,發現禾本科、菊科、藜科植物在地上植被群落和土壤種子庫中出現比例均較高,且隨著海拔升高,禾本科和藜科植物占比減少,菊科植物占比增加。說明禾本科、菊科、藜科植物在賀蘭山適應度相對較高,且隨海拔變化而變化,繁殖策略偏向于種子繁殖。在物種生活型組成上,土壤種子庫中物種數占比最大為一年生草本,地上植被為多年生草本,差異較大,可能是因為植物的不同繁殖策略引起的,一年生草本生活史較短,種子小且量大,多年生草本、灌木或半灌木傾向于量小種子大的繁殖策略。
地上植被與土壤種子庫相似性可用來衡量該植物群落穩定性和預測該群落的發展趨勢[27]。在本研究中,有6種植物共同出現在土壤種子庫和地上植被中,16種植物僅出現在土壤種子庫中,27種植物僅出現于地上植被中,土壤種子庫和地上植被之間具有較低的相似性,除海拔1 400 m的相似性指數為0.33外,其余海拔高度相似性系數均為0.11和0.12。地上植被與土壤種子庫相似性較低,與齊丹卉[28]、張志明等[29]研究結果一致。可能是因為賀蘭山地處干旱半干旱區,氣候干旱少雨,種子在土壤種子庫中的持久性受到影響;也可能是由于地上植被的物種表現為以多年生草本占優勢,其繁殖策略形成的種子量少,對土壤種子庫形成的貢獻率較低。
植被恢復的潛力與土壤種子庫種子密度和物種多樣性密切相關[30]。土壤種子庫物種多樣性既受地上植被特征制約,同時也受土壤特性影響[31]。在本研究中,pH、電導率對土壤種子庫物種多樣性的影響較大,與王曉榮[32]、翟付群等[33]研究結果一致。可能是通過影響種子形成、儲存、萌發等過程直接或間接影響土壤種子庫,關于這方面的問題還需進一步研究。在本研究中,5個海拔高度土壤種子庫密度均較低,土壤種子庫種源尚不能滿足地上植被恢復所需種源量;5個海拔高度土壤種子庫物種多樣性均較低,無法滿足植被自然恢復需求,特別是海拔1 400 m處物種多樣性顯著低于其他海拔,恢復潛力最低。賀蘭山低山區風蝕嚴重、植被稀疏,土壤種子庫自身所具有的種子無法滿足植被自然恢復需求,可采取飛播等生態恢復措施來彌補表層土壤種子的不足,從而滿足地上植被恢復所需種源量。關于種子種類的選取和實際應用時的外界因素的干擾,還需進一步研究。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
